棉花類表皮特異性分泌糖蛋白基因GhA01EP1 的克隆和功能分析
2021-04-16 03:01:52李丹趙存鵬趙麗英劉旭劉素恩王凱輝王兆曉耿軍義郭寶生
棉花學報 2021年6期
李丹,趙存鵬,趙麗英,2,劉旭,劉素恩,王凱輝,王兆曉,耿軍義*,郭寶生*
(1.河北省農林科學院棉花研究所/ 農業部黃淮海半干旱區棉花生物學與遺傳育種重點實驗室/ 國家棉花改良中心河北分中心,石家 莊 050051;2.河北工程大學園林與生態工程學院,河北 邯鄲 056038)
在長期進化過程中,植物應對不斷變化的生物脅迫和非生物脅迫,發展出獨特的、高度復雜的生物調控機制來精確地控制其生長發育,研究人員對此機制已進行了許多探索。 但是,與細胞內信號傳導通路的組成和效應相比,植物分泌蛋白在植物脅迫反應中的作用還未引起足夠重視[1]。分泌蛋白作為植物與外界信息、物質、能量交流和交換的重要載體,對植物生理活動的調控起著非常重要的作用,因此,對分泌蛋白的研究有助于完善對植物抵抗環境脅迫調節機制的認識。
分泌蛋白調控作用涉及一個關鍵的生物學系統——細胞分泌系統。 細胞分泌系統就是將細胞內產物釋放進入細胞外空間。 細胞外空間通常指質膜以外的所有區域,其中的液體稱為細胞外液[2],細胞外液由分泌蛋白、代謝產物、離子和許多其他物質組成。 分泌蛋白在細胞壁結構形成、細胞間相互作用、細胞外/細胞內信號傳遞、細胞對環境刺激的響應和病原體防御等方面具有重要的生物學功能,目前研究較多的是分泌蛋白在植物逆境適應中的作用[3-8]。
許多研究已證實,環境脅迫能夠明顯地影響分泌蛋白的質量和數量[9],鹽[10]、低溫[11]、水楊酸[12]、金屬毒害[13]和病原菌入侵[14]等都對植物分泌蛋白質組具有較大影響。Song 等[15]比較了水稻鹽處理和對照的分泌蛋白質組, 鑒定出37 種差異表達分泌蛋白, 它們主要參與碳水化合物代謝、氧化還原、蛋白質的加工和降解等。 對沙棘的分泌蛋白質組分析鑒定出61 種響應低溫的蛋白, 其中34 個上調表達的蛋白質主要包括信號傳導、氧化還原調節和防御相關蛋白[16]。李元寶[17]對棉花響應大麗輪枝菌侵染的質外體蛋白質組學進行了分析,共分離到49 個差異蛋白,這些差異蛋白參與活性氧代謝、防御反應、細胞壁修復、信號傳導等多個生物學過程。 在質外體蛋白質組學研究不斷深入的同時,抗逆相關質外體蛋白的功能研究也取得了一些進展。張磊[18]研究表明,質外體蛋白OsAPRLK1 在水稻抗鹽反應中起著調控作用。 Wang 等[19]發現大豆細胞質外體定位BURP結構域蛋白GmRD22 的表達增強了植株對非生物脅迫的耐受性。 李元寶發現質外體蛋白GbNRX1 和GbCRR1 能夠提高植物對病原菌的抗性[17]。小麥中定位于細胞質、細胞膜和質外體的TaXTH-7A 蛋白可以提高轉基因擬南芥(Arabidopsis thaliana)的抗旱性[20]。 孫金月等[21]和簡令成等[22]研究發現,在環境脅迫下,抗逆性強的品種細胞表面分泌的糖蛋白增多增厚,而敏感型品種細胞表面糖蛋白則會變少甚至脫落。 在鹽脅迫下,逆境誘導轉錄因子DREB轉基因小麥葉片的細胞表面糖蛋白層更致密,野生型植株葉片的細胞表面糖蛋白層出現細胞壁外緣脫落現象[23]。 這些研究表明:在逆境條件下,細胞分泌蛋白的增加對提高植株抗逆性具有重要作用。
本研究室前期測定了耐旱棉花品種冀2658根系干旱脅迫應答早期(脅迫后1 h)的蛋白質組(數據未發表),發現一個推測為類表皮特異性分泌糖蛋白EP1 的蛋白質(Access:1050567811)差異表達倍數較高。 通過參考基因組定位和序列比對,我們將編碼此蛋白的基因命名為GhA01EP1,用實時定量聚合酶鏈式反應(Quantitative realtime PCR,qRT-PCR) 證實GhA01EP1在干旱脅迫后12 h 差異表達倍數最大。 為了研究GhA01EP1在干旱脅迫中的作用, 我們對該基因的結構、表達模式、功能等進行了初步分析,探究GhA01EP1的特性以及在植物抗旱中的作用,為進一步解析GhA01EP1的抗旱分子機制提供基礎。
1 材料與方法
1.1 試驗材料
試驗材料為河北省農林科學院棉花研究所培育的品種冀2658,經中國農業科學院棉花研究所鑒定為耐旱品種。
1.2 GhA01EP1 基因的克隆
利用Seqhunter[24]軟件將蛋白質對應的基因序列在陸地棉TM-1 基因組[25]中進行位置預測,然后提取該基因所在位置的核苷酸序列,用軟件Premier 5 在開放閱讀框 (Open reading frame,ORF)前后設計引物(正向引物:GGAATCCAATGTGCCTTTATTG;反向引物:CAAATACTTAATAGGGATTCCCG), 以冀2658 根部組織的cDNA 為模板克隆基因。
1.3 GhA01EP1 基因表達分析
將種子種植在滅菌的蛭石中,待子葉完全平展,將幼苗轉移至水培箱,生長到二葉一心時,取植株的根、莖和葉片分析GhA01EP1組織特異性表達,每個組織取3 個生物學重復。
將長至二葉一心的植株用13%(質量分數)PEG6000 溶液處理(模擬干旱脅迫),取脅迫后0 h、0.25 h、0.50 h、1 h、3 h、6 h、12 h、24 h 的幼苗根部組織提取RNA 進行反轉錄, 利用qRT-PCR檢測GhA01EP1在干旱脅迫前后表達量的變化,每個時間點取3 個生物學重復。
1.4 RNA 提取和qRT-PCR 檢測
用BioFlux Plant Total RNA Extraction Kit(DNA-free)試劑盒(購自BioFlux)按照說明書制備總RNA。 第一鏈cDNA 合成和qRT-PCR 分別采用HiScript IIQ RT SuperMix for qPCR(+gDNA wiper)試劑盒(購自諾唯贊)和SuperReal PreMix Plus(SYBR-Green)(購自天根)進行。 在Biorad CFX96 實時熒光定量PCR 儀中進行qRT-PCR,以1 μL 合成的cDNA 為模板, 以棉花Actin1作為參考基因。 基因的相對表達量采用2-ΔΔCt法分析,設3 次技術重復。 利用Premier 5 軟件設計基因特異性引物,GhA01EP1的正向引物ATTTACGGGTATTACAGCAGTCCA, 反向引物為TGAACATTTCCAATCAACTCCG; 內參基因Actin1正向引物為ATCCTCCGTCTTGACCTTG,反向引物為TGTCCGTCAGGCAACTCAT。
1.5 生物信息學分析
通過KEGG(http://www.genome.jp/kegg/)數據庫分析GhA01EP1 可能參與的主要代謝途徑。利用在線軟件ExPASy-ProtParam(https://web.ex pasy.org/protparam/)對GhA01EP1 蛋白的理化性質進行分析;運用SignalP 5.0 Server(http://www.cbs.dtu.dk/services/SignalP)和TMHMM Server 2.0(http://www.cbs.dtu.dk/services/TMHMM)分別進行信號肽及跨膜結構預測; 利用PSI-predictor(http://bis.zju.edu.cn/psi/) 進行亞細胞定位預測;運用SPOMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=np%20sa_sopma.html)和Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index) 分別進行GhA01EP1 蛋白二級結構和三維模型的預測;在NCBI(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM=blastp&PAGE_TYPE=BlastSearch&LINK_LOC=blasthome)分析GhA01EP1 蛋白結構,并利用Blastn(https://blast.ncbi.nlm.nih.gov/Blast.cgi?PROGRAM = blastn&PAGE _ TYPE = BlastSearch&LINK_LOC=blasthome 對GhA01EP1基因的同源序列進行聚類分析。
1.6 亞細胞定位
本試驗所用載體為pCAMBIA2300-GFP,經SacI 和XbaI 雙酶切后,回收大片段,與目的基因連接轉化。 采用農桿菌轉化法將構建好的過表達載體CaMV 35S∷GhA01EP1-GFP 轉化農桿菌菌株GV3101。按照煙草瞬時轉化方法,將合適濃度的農桿菌菌液注射進完整且厚的煙草葉片中,然后將煙草放在黑暗處過夜。一般注射后2~3 d 表達量最高,切取侵染區域,撕下表皮制片,用顯微鏡觀察。
1.7 轉基因擬南芥的獲得和檢測
利用農桿菌蘸花法進行擬南芥野生型(Col-0)轉化,將滅菌的T0代轉基因擬南芥種子置于含50 mg·L-1卡那霉素的MS 培養基平板上篩選,選取長勢較好的植株轉移至基質中,待長出7~8 片真葉時,以野生型擬南芥為陰性對照,取最底端葉片用于PCR 檢測, 正向引物CCTTCGCAAGACCCTTCCTC,反向引物CCACCCCGGTGAACAGC。 然后用同樣的方法篩選獲得T2代和T3代陽性轉基因擬南芥種子。
1.8 干旱脅迫下轉基因擬南芥表型及根長測定
將Col-0 種子和T3代轉基因純系(共5 個株系:D1、D2、D4、D7、D8)種子(種子存放在4 ℃,不再春化處理)滅菌后種在MS 培養基上(22 ℃,16 h 光照/8 h 黑暗), 待子葉完全展開后將長勢一致的幼苗分別轉移至含有0 mmol·L-1、250 mmol·L-1、300 mmol·L-1D-甘露醇的MS培養基, 垂直放置于人工培養箱中培養,8 d 后觀察并測定野生型和轉基因擬南芥株系的根長。
1.9 干旱復水法鑒定轉基因擬南芥的抗旱性
Col-0 和轉基因擬南芥種子消毒后在MS 培養基上培養1 周,然后轉移至土壤中,每個品系種24 棵植株,設3 個重復。 在人工培養箱中(條件同1.8)繼續培養,待幼苗生長較健壯時,停止澆水,隨時觀察擬南芥生長狀態;等幼苗出現嚴重干枯、萎蔫時,對植株進行復水并觀察擬南芥生長狀態。
2 結果與分析
2.1 GhA01EP1 的克隆與序列分析
利用同源克隆法從冀2658 中克隆得到GhA01EP1的ORF 序列, 序列全長1 371 bp,編碼456 個氨基酸。GhA01EP1位于A01 號染色體上。 在NCBI 上對氨基酸序列進行結構分析,發現序列包含B-lectin、Plant PAN/APPLE-like 2 個結構域。
2.2 GhA01EP1 的表達分析
取植株的根、莖和葉片對GhA01EP1進行組織特異性表達分析,結果顯示(圖1),GhA01EP1在根、莖和葉片中均表達,且在根中表達量最高。為驗證GhA01EP1與干旱脅迫的相關性,對干旱脅迫前后GhA01EP1在根部表達量的變化進行了分析,GhA01EP1的表達量在脅迫初期(脅迫后0.25 h 和0.50 h)呈下降趨勢,但1 h 后明顯上升,12 h 后達到峰值(圖2),說明GhA01EP1響應干旱脅迫。
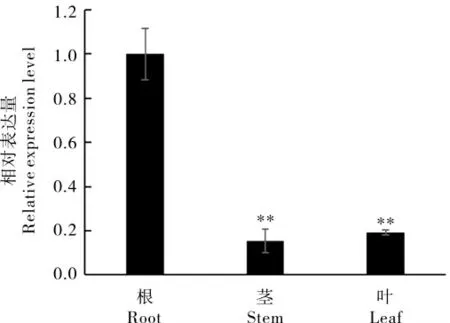
圖1 GhA01EP1 的組織表達分析Fig. 1 The expression levels of GhA01EP1 in roots,stems, and leaves
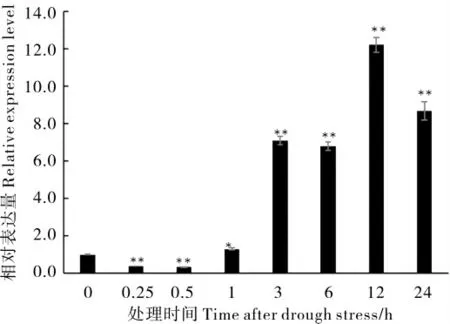
圖2 GhA01EP1 在干旱脅迫下根部的表達變化Fig. 2 Expression pattern of GhA01EP1 under drought stress in root
2.3 GhA01EP1 的代謝通路分析
根據蛋白質組KEGG 通路分析發現,GhA01EP1在7 種代謝途徑中上調表達, 包括甘氨酸、絲氨酸和蘇氨酸代謝、苯丙氨酸代謝、異喹啉類生物堿的生物合成、β- 丙氨酸代謝、 莨菪烷類、哌啶、吡啶生物堿生物合成、酪氨酸代謝和mRNA 監視通路等,其中前兩種代謝途徑可信度較高。 在甘氨酸、 絲氨酸和蘇氨酸代謝途徑中,GhA01EP1參與合成甲基乙二醛(Methylglyoxal,MG)(附圖1)。近幾年的研究表明,高濃度的MG是一種細胞毒素,而低濃度的MG 作為一種信號分子,在植物遭受脅迫時含量迅速上升,可調節多種生理過程和抗逆性[26-29]。 苯丙氨酸代謝途徑是植物生成木質素和芪類化合物等次級代謝產物的重要途徑[30],木質素是細胞壁的重要組成部分,木質素填充于纖維素構架中增強植物的機械強度,利于輸導組織的水分運輸、抵抗不良外界環境的侵襲[31]。這表明,GhA01EP1可能通過以下2 種途徑響應干旱脅迫: 一是通過合成積累MG響應脅迫信號,二是通過細胞壁增厚協助輸導組織水分運輸來抵抗干旱脅迫。
2.4 GhA01EP1 的生物信息學分析
利用ExPASy-ProtParam 分析得到GhA01EP1蛋白的分子式為C2257H3464N602O684S21; 分子質量為50 649.21 D;理論等電點為6.61;不穩定指數為37.20,為穩定性蛋白;總親水性平均數為-0.235。GhA01EP1 蛋白包含456 個氨基酸, 其中亮氨酸占10.1%,絲氨酸占9.9%,蘇氨酸占7.2%,甘氨酸占7.5%。
生物信息學分析顯示,GhA01EP1 蛋白N 端存在信號肽的概率為68.7%, 信號肽剪切位點位于第42 個與第43 個氨基酸之間(附圖2);預測不存在跨膜區域; 亞細胞定位預測結果為GhA01EP1 定位在細胞外的概率為75%(附圖3)。
預測顯示,GhA01EP1 蛋白質二級結構中α-螺旋占8.55%,延伸主鏈占32.02%,無規則卷曲占53.29%,β-折疊占6.14%。 三維模型預測結果見圖3,模型的置信度達到100%,氨基酸序列的覆蓋度達到83%,因此,可以認為該蛋白結構的預測是可靠的。
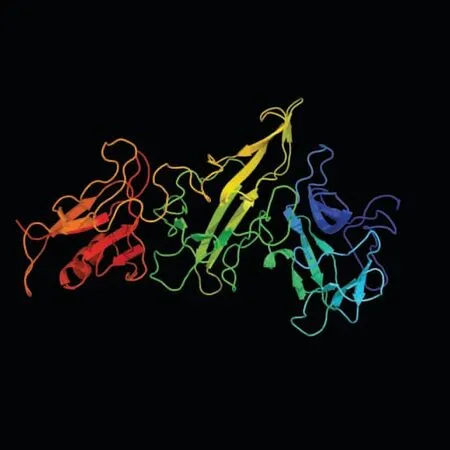
圖3 GhA01EP1 蛋白三維模型Fig. 3 The 3D model of GhA01EP1 protein
2.5 GhA01EP1 同源蛋白比較和同源基因聚類分析
將GhA01EP1 蛋白序列在UniProt(https://www.uniprot.org/)網站進行檢索比對,發現編號為A0A1U8KDH7 的序列和GhA01EP1 同源性最高, 只有1 個氨基酸差異 (圖4)。 經分析,A0A1U8KDH7 為來自于墨西哥棉和陸地棉的Epidermis-specific secreted glycoprotein EP1-like,但A0A1U8KDH7 基因功能尚未被注釋。此外,我們在NCBI 上進行Blastn 比對,選取相似度較高且來源于不同物種的序列進行了聚類分析 (圖5),這些序列明顯被分成了3 大類,來源于棉屬的序列被分成同一類,木槿(Hibiscus syriacus)被單獨分為一類, 榴蓮 (Durio zibethinus)、 可可(Theobroma cacao) 和哥倫比亞錦葵(Herrania umbratica)被分為一類,這些物種都屬于錦葵目。

圖4 GhA01EP1 和A0A1U8KDH7 蛋白序列比對Fig. 4 Sequence alignment of GhA01EP1 and A0A1U8KDH7 protein
2.6 GhA01EP1 的亞細胞定位
對GhA01EP1 進行亞細胞定位分析, 對照35S∷GFP 的綠色熒光分布在整個細胞中; 在正常狀態下(質壁分離前),GhA01EP1 與GFP 融合蛋白的熒光定位于細胞膜和/ 或細胞壁上,經質壁分離發現,GhA01EP1 與GFP 融合蛋白被分泌到胞外,定位于細胞壁和細胞膜之間(圖6),這與在線預測結果一致。
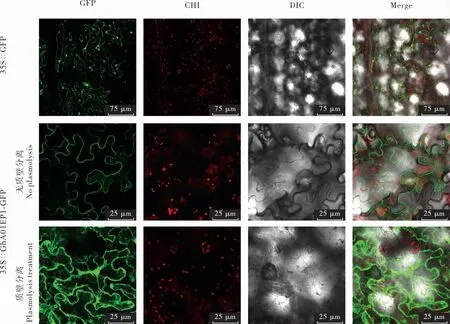
圖6 GhA01EP1 的亞細胞定位Fig. 6 Subcellular localization of GhA01EP1 protein
2.8 干旱脅迫下轉基因擬南芥抗旱性分析
用農桿菌蘸花法轉化擬南芥, 經篩選獲得5個T3代純系(圖7)。 對Col-0 和轉基因擬南芥進行干旱處理,比較其抗旱性。 從圖8 可以看出,在0 mmol·L-1D- 甘露醇處理下,Col-0 和轉基因株系的根長沒有明顯差異; 在250 mmol·L-1和300 mmol·L-1D- 甘露醇MS 培養基中,Col-0 和轉基因株系生長受到抑制,根系變短、葉片變小,而且隨著濃度的增加抑制程度更強,但轉基因株系比Col-0 植株根系長。 在250 mmol·L-1D- 甘露醇脅迫下,D4 和D7 株系與Col-0 根長呈極顯著差異(圖8E)。 在300 mmol·L-1D-甘露醇脅迫下,D1、D2 和D8 與Col-0 根長呈顯著差異,D4和D7 株系與Col-0 根長呈極顯著差異(圖8F)。同時, 我們觀察到300 mmol·L-1D- 甘露醇脅迫下Col-0 和轉基因株系之間植株大小存在差異,于是我們將擬南芥根部切除,根部以上部分固定在干凈的MS 培養基上(圖8G),從圖中可以看出,株系D1、D2 和D8 的植株明顯比野生型擬南芥偏大,但D4 和D7 株系與野生型差異不明顯,可能D4 和D7 株系中更多營養向根系供給。
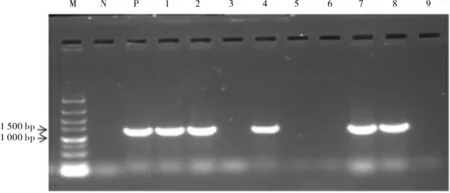
圖7 擬南芥株系的PCR 檢測結果Fig. 7 Identification of GhA01EP1 by PCR in transgenic A. thaliana
為了對轉基因擬南芥株系進行抗旱鑒定,對正常生長條件下生長3 周左右的Col-0 和轉基因擬南芥植株進行干旱處理, 停止澆水后30 d,幾乎所有植株都受到脅迫影響,葉片發黃、干枯、萎蔫,但是轉基因植株受影響程度較低(圖8H)。 復水后5 d, 部分轉基因植株可以恢復正常生長狀態,存活率為33%~50%;而Col-0 植株的存活率只有16.7%。以上結果表明,GhA01EP1可以提高干旱脅迫下轉基因擬南芥的抗旱性。
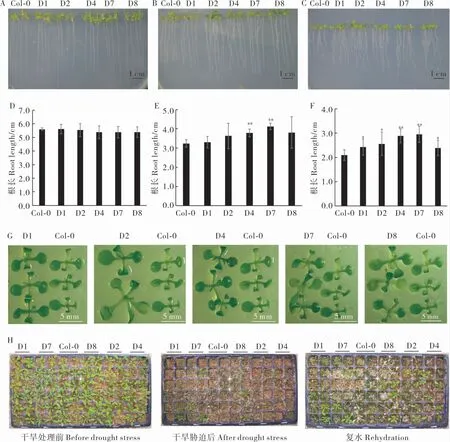
圖8 轉基因擬南芥抗旱性分析Fig. 8 Drought stress tolerance of transgenic Arabidopsis
3 討論
植物質外體作為連接環境和植物原生質體的紐帶, 在植物代謝和信號傳遞中具有多重功能[32]。 作為質外體的重要組成部分,分泌蛋白調控許多生理和生物學過程,如信號傳導、細胞擴張、應激反應和細胞間相互作用等。 本研究發現GhA01EP1 蛋白被分泌到細胞膜和細胞壁之間,為分泌蛋白。 利用轉GhA01EP1的擬南芥證實,GhA01EP1能夠提高轉基因植株的抗旱能力,與抗旱調控相關。
將GhA01EP1 蛋白在UniProt 進行blast 比對,共比對到250 條序列,只有2 條經過驗證,分別是來自擬南芥(Arabidopsis thaliana)的類EP1糖蛋白3 (Entry:Q9ZVA4) 和類EP1 糖蛋白4(Entry:Q9ZVA5), 它們和GhA01EP1 的一致性分別是50.9%和51.3%。 類EP1 糖蛋白3 定位在細胞壁[33-34],可能參與細胞間程序性細胞死亡信號機制[35]。 類EP1 糖蛋白4 定位在細胞壁[34]和細胞外區域[36],GO(Gene Ontology)功能注釋顯示,此蛋白可以和糖結合。類EP1 糖蛋白3 和4 雖已證實定位在細胞壁上, 但其具體功能仍然未知。本研究中,KEGG 通路分析GhA01EP1 蛋白參與苯丙氨酸代謝,與木質素的合成有關,而木質素是細胞壁的重要組成部分,所以GhA01EP1 蛋白可能參與細胞壁合成,調節細胞壁代謝以適應干旱脅迫環境。
另外,GhA01EP1基因還調節甘氨酸、絲氨酸和蘇氨酸的代謝,參與合成MG。 MG 被認為是一種細胞毒害劑[26,37-38],而近幾年的研究顯示,低濃度的MG 是植物響應環境脅迫的信號。乙二醛酶系統包括乙二醛酶Ⅰ(glyoxalaseⅠ,GlyⅠ)、乙二醛酶Ⅱ(glyoxalaseⅡ,GlyⅡ)及乙二醛酶Ⅲ(glyoxalase Ⅲ,GlyⅢ)[26-27],是植物保護機制之一。 乙二醛酶系統參與MG 的主要清除途徑,不僅可直接清除過量的MG,還能通過增強抗氧化酶、抗氧化劑等抵抗MG 脅迫和由MG 引發的氧化脅迫[27,39-40]。 研究發現,外源MG 能提高NaCl 脅迫下小麥幼苗體內GlyⅠ和GlyⅡ的活性,減輕NaCl 對小麥的抑制作用。 同時,MG 可以提高小麥細胞中超氧化物歧化酶、過氧化氫酶、抗壞血酸過氧化物酶和谷胱甘肽還原酶的活性,并提高谷胱甘肽和抗壞血酸水平, 降低NaCl 脅迫下超氧化物自由基、過氧化氫和丙二醛的含量[41]。 在玉米中,MG 可誘導玉米幼苗的耐熱性, 抗氧化系統(抗氧化酶和非酶抗氧化劑)和乙二醛酶系統(GlyⅠ和GlyⅡ)在其中發揮重要作用[42]。
綜上所述, 在棉花抵御干旱脅迫的過程中,GhA01EP1上調表達可能導致MG 合成增多,激活乙二醛酶系統和過氧化酶系統提高植株的抗旱性。棉屬及其近緣種屬中有關GhA01EP1功能的研究尚未見報道, 雖然本研究證實了GhA01EP1可以提高轉基因擬南芥的抗旱能力,但其作用機理尚不明晰。 需要進一步驗證內源MG 作為信號分子是通過激活乙醛酶系統和抗氧化系統調控植株適應性,還是通過苯丙氨酸代謝促進木質素合成、提高細胞壁厚度控制水分流動提高抗旱性。
4 結論
本研究通過陸地棉根部蛋白質組測序發現,一個被預測為類表皮特異性分泌糖蛋白EP1 的蛋白質在干旱脅迫下上調表達, 我們將控制其合成的基因命名為GhA01EP1。對過表達GhA01EP1的轉基因擬南芥進行抗旱鑒定發現,轉基因擬南芥相比Col-0 抗旱性增強, 脅迫復水后更容易恢復到正常生長狀態,說明GhA01EP1對提高植物抗旱性起積極作用。
附圖:
詳見本刊網站(http://journal.cricaas.com.cn)本文網頁版。
附圖1 GhA01EP1 參與甘氨酸、絲氨酸和蘇氨酸代謝
Fig. S1 GhA01EP1 is involved in glycine, serine and threonine metabolism
附圖2 GhA01EP1 信號肽預測
Fig.S2 The prediction of signal peptide of GhA01EP1
附圖3 GhA01EP1 亞細胞定位預測結果
Fig.S3 The prediction result of subcellular localization of GhA01EP1
