棉花HDAC 基因家族鑒定及其在黃萎病菌侵染下的表達分析
2021-04-16 02:59:22王艷情鄭杰許艷超蔡小彥周忠麗侯宇清王坤波王玉紅陳浩東劉方李志坤
棉花學報 2021年6期
關鍵詞:分析
王艷情,鄭杰,許艷超,蔡小彥,周忠麗,侯宇清,王坤波,王玉紅,陳浩東,劉方*,李志坤
(1.河北農業大學/ 華北作物改良與調控國家重點實驗室,河北 保定 071001;2.中國農業科學院棉花研究所/ 棉花生物學國家重點實驗室,河南 安陽 455000)
棉花是重要的經濟作物,其產量和品質經常受到黃萎病的影響[1]。 我國棉田中的黃萎病主要由大麗輪枝菌(Verticillium dahliae)引起[2]。 黃萎病也被稱為棉花的“癌癥”[3],棉株感染大麗輪枝菌后,葉片嚴重失綠,形成黃色病斑,造成減產甚至絕收[4]。棉屬(Gossypium)有4 個栽培棉種和49個野生棉種[5-6]。 栽培棉種對病蟲害及非生物脅迫較為敏感,而野生棉種長期生長在惡劣的自然環境中, 具有豐富的遺傳多樣性與抗病蟲害基因源[7],是棉花遺傳改良的優異資源。Dong 等[8]比較分析了瑟伯氏棉(Gossypium thurberi)、雷蒙德氏棉(G.raimondii)和三裂棉(G.trilobum)在大麗輪枝菌脅迫下的表型,發現瑟伯氏棉具有很強的黃萎病抗性。 因此,進一步挖掘野生棉,特別是瑟伯氏棉中的抗病基因對棉花抗黃萎病遺傳改良具有重要意義。
組蛋白去乙酰化酶 (Histone deacetylase,HDAC)通過降低染色體的乙酰化水平從而抑制轉錄[9]。 植物中HDAC 家族分為3 個亞家族,分別是RPD3/HDA1(Reduced potassium dependency 3/Histone deacetylase 1)、SIR2 (Silent information regulator 2)和HD2(Histone deacetylase 2)。 RPD3/HDA1 作為該家族最大的亞家族,成員具有典型的HDAC 結構域[10]。 SIR2 又被稱為Sirtuins,該亞族成員與其他HDAC 沒有結構相似性,并且需要煙酰胺腺嘌呤二核苷酸(NAD)作為輔助因子[11]。 HD2 亞家族成員主要有3 個結構域:具有保守五肽序列的氮末端結構域、高電荷的中央酸性結構域和可變的碳末端結構域[12];HD2 首先在玉米中被鑒定,僅存在于植物中[13]。
近年來,在擬南芥[14]、水稻[15]和大豆[16]基因組中分別鑒定出18、17 和28 個HDAC家族基因,并發現其在植株生長發育與響應逆境脅迫中具有重要功能。 擬南芥中AtHDA6和AtHDA19均表現為促進開花[17],而AtHDA9抑制開花[18]。擬南芥AtHDA9過表達植株對鹽脅迫更加敏感;相反,hda9突變體植株對鹽脅迫的抗性增強[19]。 此外,HDAC 家族成員在病原菌防御中也起著關鍵作用,過表達AtHDAC19基因的擬南芥通過激活ERF1而增加病程相關蛋白(Pathogenesis-related proteins,PRs)的表達增強植株的抗性,沉默該基因則導致植株對病原菌更加敏感[20]。
棉花GhHDT4D 通過調節GhWRKY33啟動子區H3K9 乙酰化水平, 從而抑制GhWRKY33的表達,在應對干旱脅迫響應中發揮作用[21]。 然而,HDAC 在棉花抗黃萎病方面的作用還未見報道。 本研究以棉花基因組數據為基礎,鑒定并比較分析了陸地棉、亞洲棉和瑟伯氏棉中HDAC基因家族成員基因結構、保守結構域、進化關系及順式作用元件的差異,并結合瑟伯氏棉轉錄組數據[8]和實時熒光定量聚合酶鏈反應(Quantitative real-time polymerase chain reaction,qRT-PCR)明確了在黃萎病菌侵染情況下GthHDAC家族基因的表達模式,研究結果為進一步挖掘野生棉中優異抗病基因提供了基礎,并為棉花抗病育種提供了新的基因資源。
1 材料與方法
1.1 試驗材料
所用的二倍體瑟伯氏棉(D1)由中國農業科學院棉花研究所野生棉課題組/ 國家種質三亞野生棉圃提供。
1.2 試驗方法
1.2.1棉花HDAC 基因家族成員的鑒定。二倍體亞洲棉(G.arboreum,A2)和瑟伯氏棉、四倍體陸地棉(G. hirsutum,AD1)基因組數據來自中國農業科學院棉花研究所(http://cgp.genomics.org.cn/page/species/index.jsp)。 按照李曉斐等[22]的方法從Pfam[23](http://pfam.xfam.org/)數據庫中下載HDAC結構域(PF00850)、SRT 結構域(PF02146),從TAIR(http://www.arabidopsis.org)網站下載擬南芥HD2 的氨基酸序列后,以PF00850、PF02146以及HD2 的序列為種子序列,利用本地Blast 搜索3個棉種的蛋白質數據庫,得到候選HDAC 家族蛋白序列。 通過在線網站Pfam、NCBI CDD 數據庫(http://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.sg)網站驗證候選HDAC 蛋白中的保守結構域。
1.2.2基因結構、保守基序和順式作用元件分析。HDAC家族基因的基因結構信息,包括開放閱讀框(Open reading frame,ORF)長度、蛋白質長度、外顯子數均從棉花功能基因數據庫(https://cottonfgd.org/)獲得。 使用MEME(http://memesuite.org/)在線網站分析家族成員的保守基序,并使用TBtools[24]軟件對其進行可視化處理。 使用ExPaSyServer(http://web.expasy.org/computer_pi) 預測蛋白質的相對分子質量和等電點。 使用TBtools 軟件獲得HDAC基因起始密碼子上游的2 000 bp 序列, 然后將獲得的序列提交數據庫PlantCARE(http://www.dna.affrc.go.jp/PLACE/signalscan.html),預測其中的順式作用元件。
1.2.3染色體分布和亞細胞定位預測。 根據HDAC家族成員的基因ID,使用TBtools 軟件在3 個棉種的基因組注釋文件中進行搜索, 以確定HDAC基因的染色體位置信息并對其進行可視化處理。 使用在線工具TargetP 1.1(http://www.cds.dtu.dk/services/TargetP)對HDAC基因編碼的蛋白進行亞細胞定位預測,并通過WoLFPSORT(https://wolfpsort.hgc.jp/) 網站中植物模式進行驗證。
1.2.4系統發育分析。為了更好地了解HDAC基因家族成員的進化關系, 從TAIR 網站下載擬南芥HDAC基因家族的蛋白質序列,與鑒定出的陸地棉、亞洲棉和瑟伯氏棉中HDAC基因編碼的氨基酸序列進行多序列比對。 利用MEGA X[25]軟件,采用最大似然法(Maximum likelihood method)構建系統發育樹,Bootstrap 值設為1 000,其余參數均為默認值。
1.2.5脅迫處理。 瑟伯氏棉幼苗長到兩葉一心時,采用傷根法[25]接種大麗輪枝菌,取接種后0 h、24 h、48 h 的根、莖、葉于液氮速凍,放入超低溫冰箱(-80 ℃)保存,用于后續試驗。 每個處理設3次生物學重復。
在棉花兩葉一心期分別使用5 mg·L-1乙烯(Ethylene, ET)、10 μmol·L-1茉莉酸(Jasmonic acid,JA) 和10 μmol·L-1水楊酸 (Salicylic acid,SA)均勻地噴施在葉片上,在處理后0 h、6 h、12 h和48 h 收集根、莖、葉。所有樣品在液氮速凍后放入超低溫冰箱(-80 ℃)保存,用于后續基因表達分析。 每個處理設3 次生物學重復。
1.2.6瑟伯氏棉HDAC 基因家族抗病表達模式分析。 基于大麗輪枝菌脅迫下瑟伯氏棉根、莖、葉的轉錄組數據, 將HDAC家族基因的表達量用FPKM(Fragments per kilobase of exon per million fragments mapped)值表示,經log2(1+FPKM)標準化處理, 采用TBtools 軟件繪制熱圖并進行歸一化處理,最終得到HDAC家族成員的熱圖。
1.2.7RNA 提取和qRT-PCR。 采用RNAprep Pure Plant Plus Kit 提取試劑盒提取樣品RNA,通過瓊脂糖凝膠電泳以及分光光度計檢測樣品質量。使用TranScript-All-in-One First-Strand cDNA Synthesis SuperMix 反轉錄試劑盒獲得每個樣品的cDNA。 使用Primer Premier 5 設計GthHDAC家族基因的特異性引物(表1),以棉花GthUBQ7(Genebank:No. AY189972) 作為內參基因進行qRT-PCR 分析,按照2-ΔΔCt法計算基因的相對表達量,具體方法見參考文獻[26],每組數據來自3個生物學重復以及3 次技術重復。
2 結果與分析
2.1 棉花HDAC 家族基因的鑒定
在陸地棉、亞洲棉以及瑟伯氏棉中分別鑒定出30、15 和13 個HDAC基因, 按照其在染色體上的位置分別命名為GhHDAC1~GhHDAC30、GaHDAC1~GaHDAC15、GthHDAC1~GthHDAC13(附表1~3)。
理化性質分析顯示,3 個棉種HDAC家族成員的ORF 序列差異較大,長度從480 到2 100 bp,大多數為1 000~2 000 bp。陸地棉HDAC基因的平均外顯子長度 (212.62 bp) 明顯大于亞洲棉(145.88 bp)和瑟伯氏棉(138.21 bp)HDAC基因(圖1A); 亞洲棉HDAC成員的平均內含子長度大于陸地棉和瑟伯氏棉HDAC家族基因 (圖1B);瑟伯氏棉HDAC家族成員的外顯子數量與亞洲棉和陸地棉HDAC 家族成員的外顯子數量差異較大,而亞洲棉和陸地棉間無明顯差異(圖1C);3 個棉種的HDAC基因ORF 的GC 含量差異較小,在40%~48%(圖1D)。 亞細胞定位預測結果顯示,3 個棉種大多數HDAC 蛋白定位在細胞核中,部分定位在細胞質,個別定位于線粒體和葉綠體(附表1~3)。
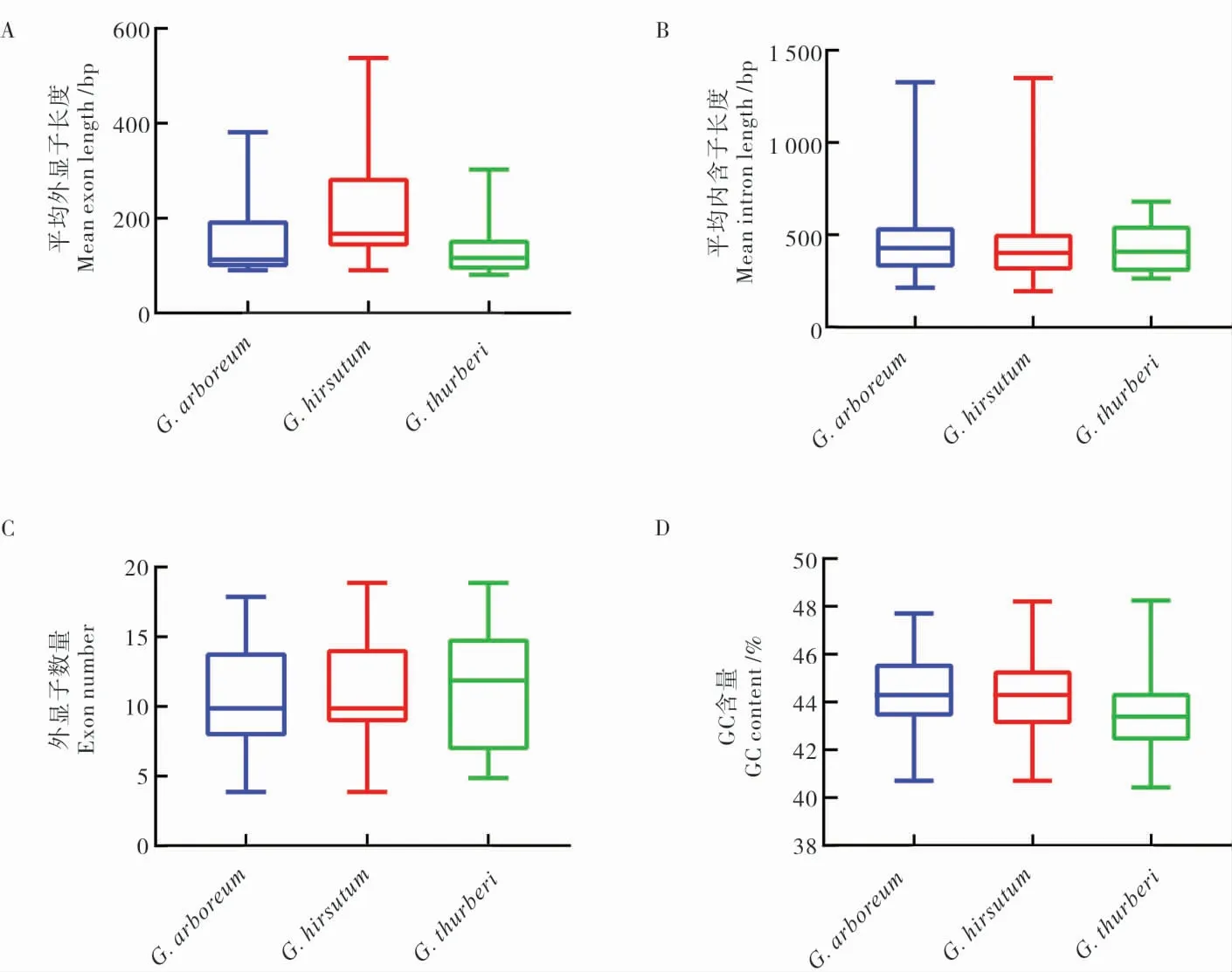
圖1 3 個棉種中HDAC 家族基因結構分析Fig. 1 Structural analysis of HDAC genes in three Gossypium species

表1 qRT-PCR 所用引物序列Table 1 Primer sequences used in qRT-PCR
2.2 棉花HDAC 家族基因的染色體分布
根據亞洲棉、瑟伯氏棉以及陸地棉的基因組注釋信息, 對58 個HDAC基因進行了染色體分布分析。由結果(圖2)發現,亞洲棉HDAC家族的15 個成員分布在9 條染色體上,其中在Chr13 染色體上有較多的分布,分別是GaHDAC13、GaHDAC14和GaHDAC15。 陸地棉30 個GhHDAC基因分布在17 條染色體上,每條染色體上有1~3 個GhHDAC基因;瑟伯氏棉的13 個GthHDAC基因分布在8 條染色體上,每條染色體上有1~3 個基因, 其中13 號染色體上最多, 分別是GthHDAC11、GthHDAC12和GthHDAC13。
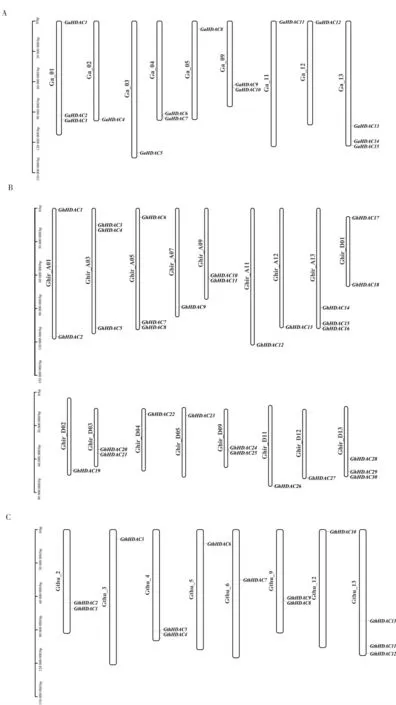
圖2 3 個棉種HDAC 基因的染色體分布Fig. 2 Chromosome distribution of HDAC genes in three Gosspium species
2.3 HDAC 基因的系統發育分析
為了探究HDAC基因的進化規律,將3 個棉種中的HDAC 蛋白與擬南芥HDAC 蛋白共同構建系統發育樹(圖3),結果顯示,3 個棉種和擬南芥中的HDAC 蛋白在RPD3/HDA1、HD2 和SIR2 亞族均有分布, 表明HDAC 蛋白在棉花和擬南芥不同物種間具有保守性。
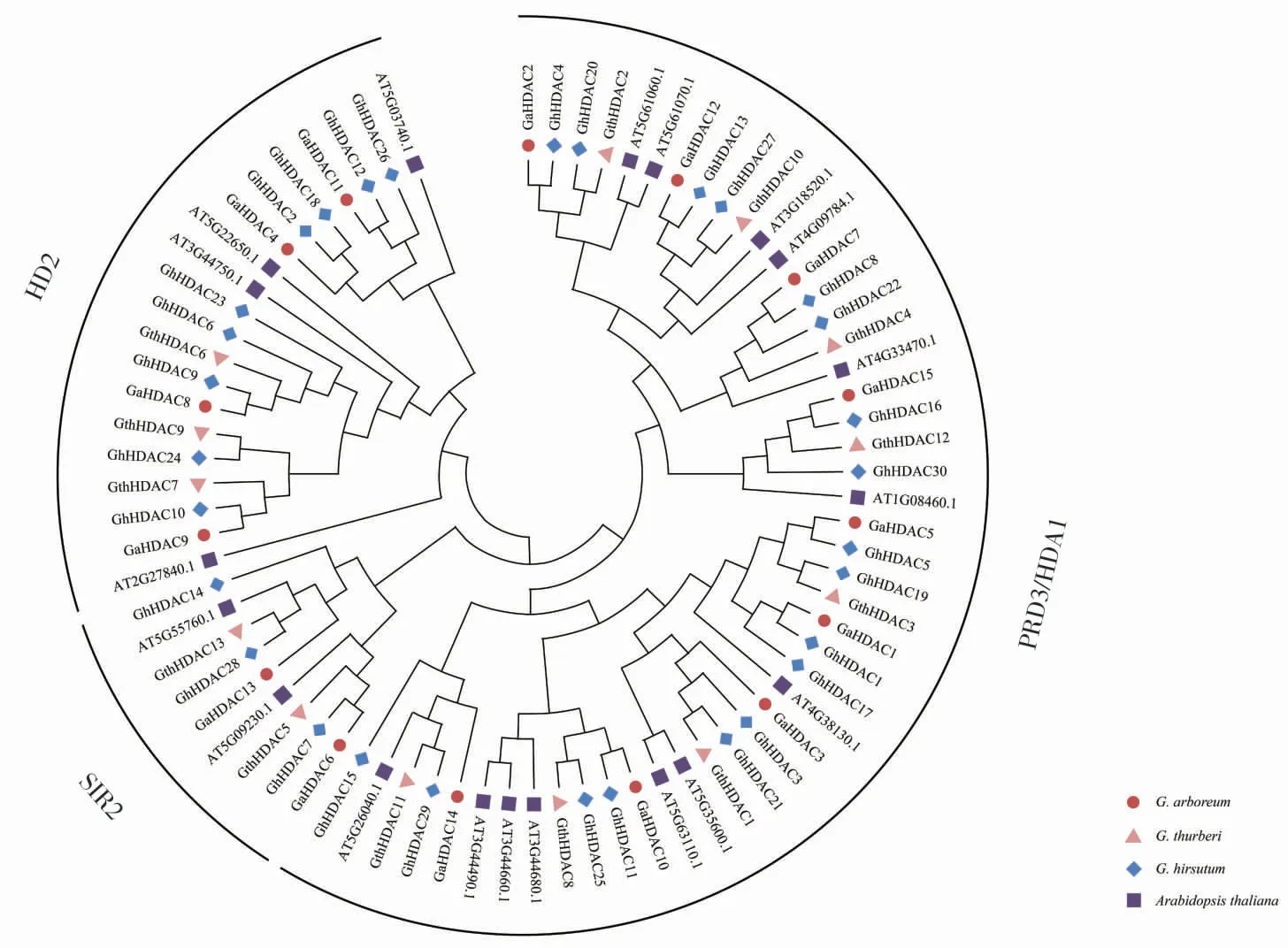
圖3 陸地棉、亞洲棉、瑟伯氏棉和擬南芥中HDAC 蛋白的系統發育分析Fig. 3 Phylogenetic analysis of HDAC proteins in three Gosspium species and Arabidopsis thaliana
2.4 棉花HDAC 蛋白質結構域及其保守基序分析
保守基序及結構域分析(圖4)顯示,SIR2 中的保守基序為motif3、motif9;RPD3/HDA1 的保守基序最多, 分別是motif1、motif2、motif4、motif5、motif6、motif7、motif8 和motif10;HD2 中的成員僅含有motif10。 由此可見,同一亞族成員含有相同的保守基序, 推測它們可能具有相似功能,不同亞組間的保守基序差異較大,推測在不同亞族的HDAC 蛋白可能存在功能差異。
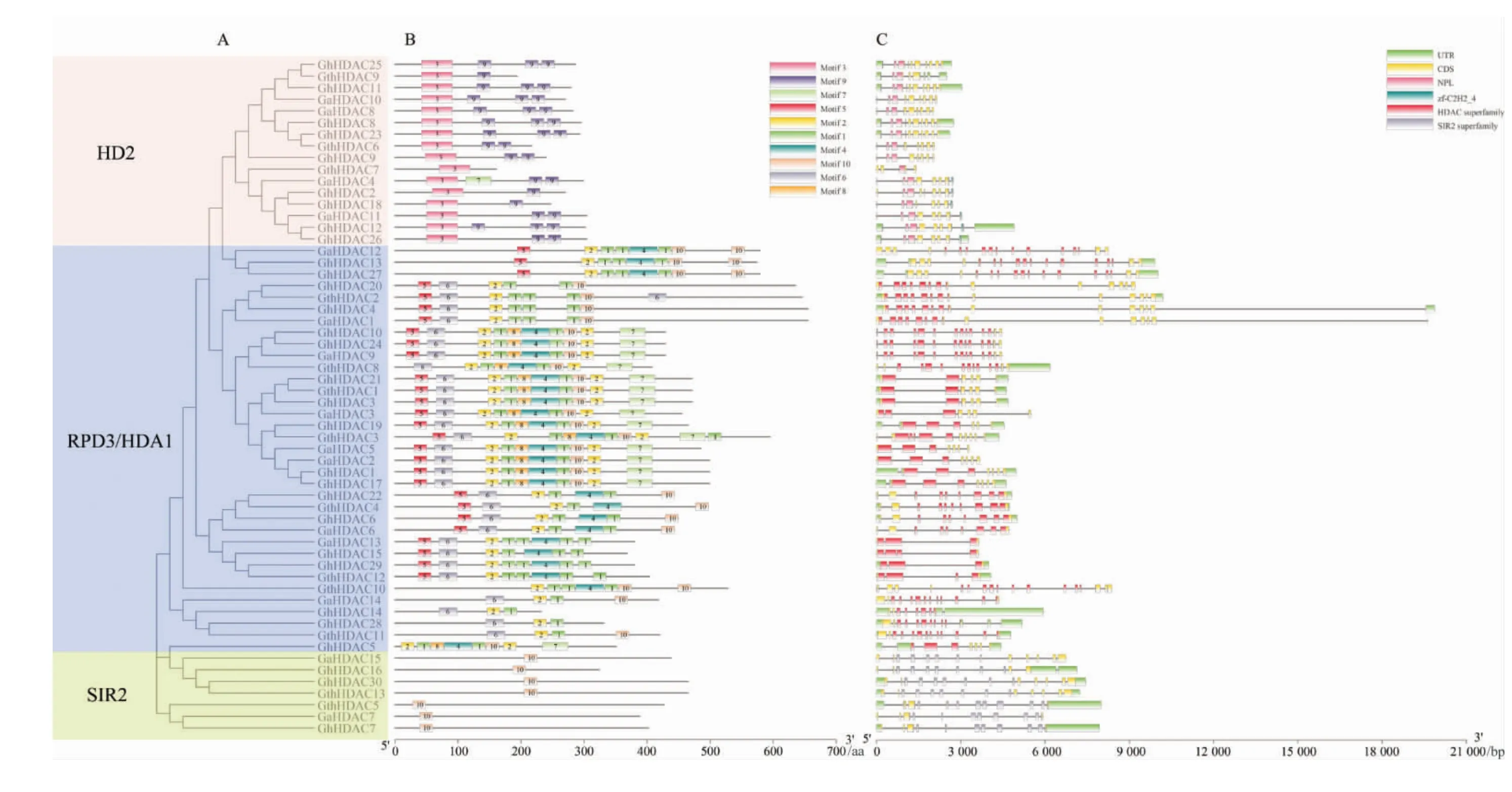
圖4 3 個棉種HDAC 的結構分析Fig. 4 Structural analysis of HDAC proteins in three Gossipium species
2.5 順式作用元件分析
順式作用元件預測發現,HDAC基因的啟動子區域有大量與植物生長發育相關的順式作用元件 (圖5), 如生長素反應元件(AuxRR-core,TGA-element)、分生組織表達相關的元件、胚乳表達相關的元件、MYB 反應元件。 其中,MYB 反應元件含量最多,GaHDAC基因中有120 個,GhHDAC基因中有217 個,GthHDAC基因中有125 個。 此外,在棉花HDAC基因的啟動子中還發現了一些與激素反應相關的元件,例如脫落酸反應元件(ABRE)、茉莉酸甲酯反應元件(CGTCA-motif 和TGACG-motif)、SA 反應元件(TCA-element)。還發現了一些與脅迫相關的順式作用元件,分別是干旱響應元件(MBS)、低溫響應元件(LTR)、應急應答元件(TC-rich repeats)以及一些響應病原體的順式作用元件(W-box,G-box,as-1 element 和P-box)等。
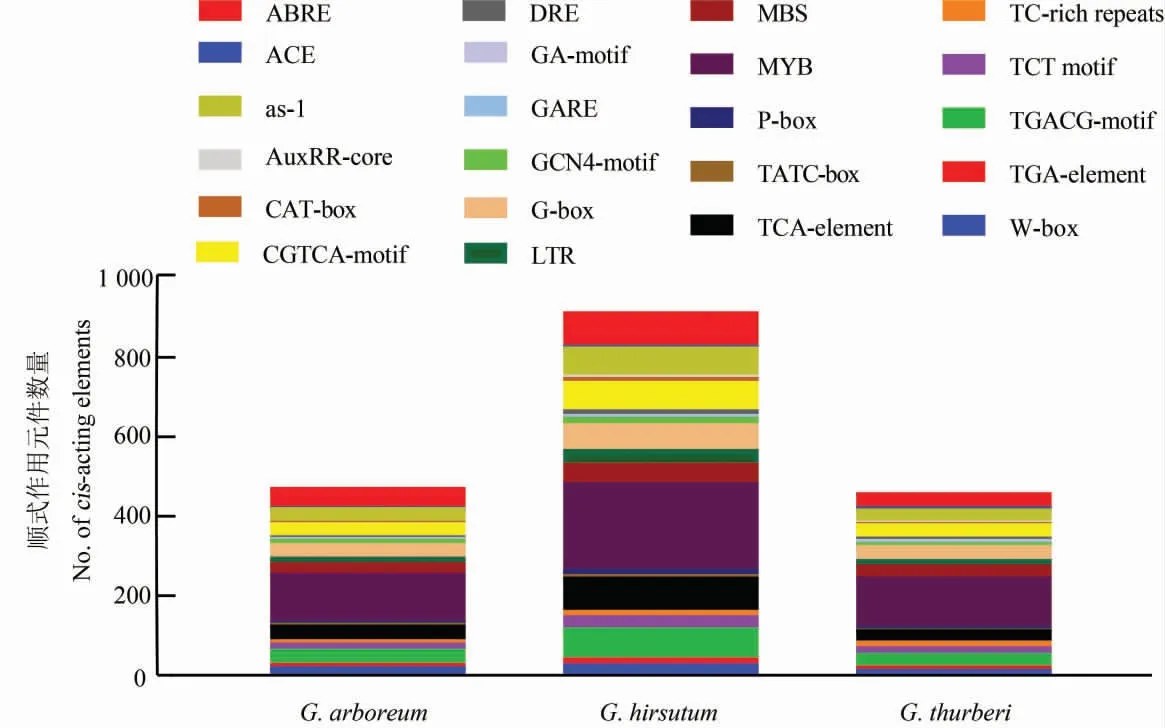
圖5 棉花HDAC 基因的順式作用元件分析Fig. 5 Cis-acting elements analysis of HDAC genes in cotton
2.6 瑟伯氏棉HDAC 基因家族在大麗輪枝菌脅迫下的表達模式分析
轉錄組數據分析表明, 瑟伯氏棉中13 個GthHDAC基因在黃萎病菌脅迫下具有不同的時空表達模式(圖6)。黃萎病菌脅迫后12 h,大部分GthHDAC基因在葉片中差異表達,其中GthHDAC1、GthHDAC2、GthHDAC6、GthHDAC8、GthHDAC9、GthHDAC10、GthHDAC12和GthHDAC13上調表達,GthHDAC3、GthHDAC4、GthHDAC5和GthHDAC7下調表達, 說明GthHDAC基因可能在棉花抵御黃萎病菌中發揮作用。 有趣的是, 在黃萎病脅迫下,GthHDAC11在根和莖中高水平表達;而在黃萎病菌脅迫后48 h,在葉片中出現一定程度的上調表達,主要在莖和葉中表達。 在黃萎病菌脅迫后12 h,GthHDAC7在莖和葉中下調表達;48 h 時其在莖中表達量提升,但在葉片中依然下調表達。
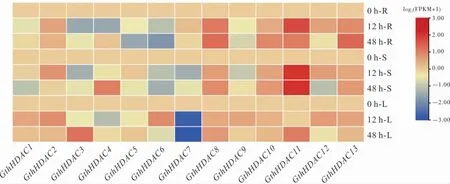
圖6 黃萎病菌脅迫下GthHDAC 基因轉錄組分析Fig. 6 Transcriptome analysis of GthHDAC genes under V. dahliae stress
利用qRT-PCR 技術進一步分析GthHDAC基因在黃萎病菌侵染后0 h、24 h 和48 h 的表達模式(圖7)。 黃萎病菌侵染后,幾乎所有GthHDAC基因在葉片中上調表達,GthHDAC4、GthHDAC7、GthHDAC11和GthHDAC12在侵染后24 h 和48 h 上調表達倍數均較高。 黃萎病菌侵染后24 h,GthHDAC7和GthHDAC13在根中顯著下調表達;GthHDAC4、GthHDAC7和GthHDAC10在莖中上調表達,GthHDAC1、GthHDAC2、Gth-HDAC3、GthHDAC5、GthHDAC8、GthHDAC11、GthHDAC12和GthHDAC13在莖中下調表達。其中:GthHDAC4在黃萎病菌侵染后24 h 在根中下調表達,48 h 時上調表達; 黃萎病菌侵染后GthHDAC7在根中下調表達,在葉片上調表達。
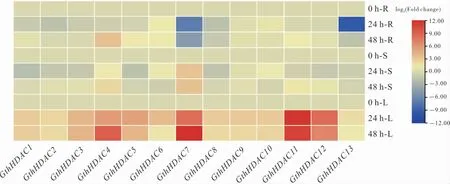
圖7 黃萎病菌脅迫下GthHDAC 基因在棉花不同組織的表達分析Fig. 7 Expression analysis of GthHDAC genes in different cotton tissues under V. dahliae stress
從響應大麗輪枝菌侵染的瑟伯氏棉HDAC基因家族成員中隨機挑選GthHDAC4、GthHDAC5、GthHDAC11和GthHDAC13四個基因,分析其在不同激素處理下的表達模式 (圖8)。GthHDAC4基因在ET 處理下表達水平逐漸上升, 在處理后48 h 達到峰值;SA 處理后48 h 表達量最高;JA 處理后12 h 達到峰值, 超過對照20 多倍。GthHDAC5基因的表達量在ET 處理后6 h 達到峰值;SA 處理后48 h,GthHDAC5表達量逐漸達到最高;JA 處理后12 h,GthHDAC5的表達量達到峰值。GthHDAC11和GthHDAC13基因的表達量在ET、SA 和JA 處理后6 h 達到峰值, 隨后降低。 上述結果表明, 瑟伯氏棉中GthHDAC基因對黃萎病菌侵染產生應答, 并且其行使功能可能與ET、SA 和JA 等信號通路有關。
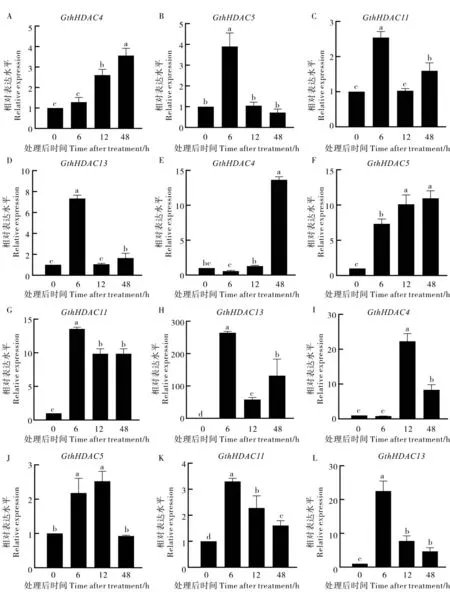
圖8 不同激素處理下GthHDAC 基因的表達Fig. 8 Expression of GthHDAC genes under different hormone treatments
3 討論
組蛋白乙酰化修飾通過調節相關基因的轉錄在植物發育和抵御病原菌侵染中發揮重要作用,而HDAC 介導的去乙酰化是組蛋白乙酰化修飾中重要的一環[27]。 四倍體陸地棉(AADD)是由A 基因組供體棉種與D 基因組供體棉種通過遠緣雜交形成的,亞洲棉屬于A 基因組棉種,具有較強的黃萎病抗性[28-32]。 瑟伯氏棉是D 基因組棉種中的抗黃萎病的棉種[26]。 因此,研究亞洲棉、瑟伯氏棉和陸地棉HDAC基因家族的系統發育關系,有利于加深對HDAC基因家族的進化及其在黃萎病抗性中的作用的認識。
本研究在陸地棉、亞洲棉以及瑟伯氏棉中分別鑒定出30、15 和13 個HDAC基因。GthHDAC基因成員數量與GaHDAC基因的成員數量之和近似于GhHDAC家族基因成員數量, 這一結果進一步印證了Beasley[33]關于四倍體種結合了A染色體組和D 染色體組的遺傳物質這一觀點。系統發育分析顯示,3 個棉種和擬南芥中的HDAC基因親緣關系較近,表明HDAC基因家族在進化上高度保守。
HDAC 通過影響植物激素合成、信號轉導等過程中相關基因的組蛋白乙酰化水平,調控相關基因的表達量,從而調節植物的生長發育、逆境響應等過程[22]。本研究發現在HDAC基因的啟動子區域富含多種激素反應元件, 如SA 脅迫響應元件、JA 脅迫響應元件、 脫落酸脅迫響應元件等。 擬南芥AtHDA6 通過與F-box 元件相互作用進而參與到JA 信號通路, 在植株抵抗病原菌過程中發揮作用[17];AtHDA19基因可以被ET 和JA 誘導, 過表達該基因可以誘導ERF1以及JA調控的病程相關蛋白基因的表達,進而增強擬南芥對蕓薹生鏈格孢菌(Alternaria brassicicola)的抗性[20]。 本研究中,在ET、SA 和JA 的處理下,GthHADC4、GthHADC5、GthHADC11和GthHADC13這4 個基因均不同程度上調表達,這些結果表明GthHDAC基因可能參與了ET/JA和SA 信號通路,進而參與到棉花黃萎病抗性反應。
Brosch 等[34]發現玉米與圓斑病菌之間的相互作用可以抑制寄主HDAC 的活性,即HC 毒素通過抑制HDAC的表達充當誘導子,激活防御反應。 本研究利用轉錄組數據分析和qRT-PCR 分析驗證了瑟伯氏棉HDAC基因家族成員在黃萎病菌脅迫下的表達譜。 但在2 種分析方式下HDAC基因的表達量存在一定差異,可能是因為兩者表達量的計算方式不同。 其中GthHDA1、GthHDAC3、GthHDAC11和GthHDAC12基因顯著上調表達,這些基因大多屬于RPD3/HDA1 亞家族。 在擬南芥中,RPD3/HDA1 型HDAC基因(HDA19和HDA6)可能在ET 和JA 信號轉導和病原體反應中發揮重要作用[17]。此外,鄧琴霖等[35]發現RPD3/HDA1 家族成員可調控植物逆境應答, 推測GthHDAC家族基因中RPD3/HDA1 亞組成員可能參與植物對黃萎病的防御。
4 結論
通過全基因組分析,在陸地棉、亞洲棉和瑟伯氏棉中分別鑒定出30、15 和13 個HDAC家族基因。 通過進化關系分析,將其分為3 個亞家族。亞細胞定位預測發現,大部分HDAC 定位于細胞核。 表達模式分析發現,在黃萎病菌脅迫下,大多數瑟伯氏棉GthHDAC基因在葉片中上調表達,并且受ET、SA 和JA 的誘導。 本研究通過解析陸地棉、亞洲棉和瑟伯氏棉中HDAC基因的進化規律,明確了不同激素處理下HDAC基因在瑟伯氏棉中的表達模式,為研究HDAC 在抵御棉花黃萎病中的作用機理奠定了基礎。
附表:
詳見本刊網站(http://journal.cricaas.com.cn)本文網頁版。
附表1 亞洲棉HDAC家族的理化性質
Table S1 Physicochemical properties ofHDACgenes inG.arboreum
附表2 陸地棉HDAC家族的理化性質
Table S2 Physicochemical properties ofHDACgenes inG.hirsutum
附表3 瑟伯氏棉HDAC的理化性質
Table S3 Physicochemical properties ofHDACgenes inG.thurberi
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
