基于高通量測序的淮山根際土壤真菌多樣性分析
2021-05-25 08:27:32勞承英申章佑周靈芝韋本輝李艷英黃渝嵐
西南農業學報 2021年1期
勞承英,申章佑,周靈芝,韋本輝,李艷英,胡 泊,黃渝嵐,周 佳
(廣西農業科學院經濟作物研究所,廣西 南寧 530007)
【研究意義】淮山(DioscoreaoppositaThunb.)為藥食同源的薯蕷科薯蕷屬一年生或多年生單子葉纏繞性藤本植物[1],其塊莖肥厚、美味、滋補,深受廣大消費者喜愛,極具經濟價值和市場前景[2]。但淮山塊莖垂直向下生長的習性使其采收難度增加,采收成本很高[3]。為降低種植和采收難度,改變深挖溝的種植和收獲方式,韋本輝等[4]發明了淮山定向結薯栽培方法(專利號:ZL200510018294.1),使淮山塊莖沿著定向膜傾斜生長以方便采收,不僅節省大量人工和成本,且產出的淮山塊莖粗大、筆直,產量和品質均有所提高。淮山袋料種植方法(專利申請號:CN201610241942.8)的原理與定向結薯栽培方法基本相同,是按照淮山品種特性和環境要求制作可滿足其生長要求的袋料(細碎松土與肥料的混合物),最后在袋料上種植淮山和收獲薯塊。不管是定向結薯栽培的土壤還是袋料種植的土壤,其理化性質及微生物種類和數量均與淮山的生長和產量密切相關。已有研究表明,高通量測序(High-throughput sequencing)技術可全面、準確地獲得微生物的群落結構信息,已在水稻、玉米、枸杞和油蒿等作物根際土壤的微生物研究中廣泛應用[5-9],且明確了這些作物根際微生物的群落組成和多樣性,但在淮山根際土壤中的應用效果尚不清楚。因此,研究淮山定向結薯栽培和袋料種植方式下淮山根際土壤微生物的種群結構,對創造有益微生物生存環境及提高袋料種植淮山的產量和質量具有重要意義。【前人研究進展】土壤微生物種群結構和多樣性受土壤類型、植被類型和空氣溫濕度等多種自然因素及耕作、施肥和除草等人為因素的影響[10]。在不同的農業生態系統中,施用化肥土壤的微生物群落多樣性與不施用化肥土壤的微生物群落存在顯著差異[11]。馬瑩[12]研究發現,以稀釋平板法測定的河南省溫縣懷山藥土壤微生物量以細菌占多數,占總量的84.28 %~93.35 %,其次是放線菌,占6.54 %~15.64 %,而真菌僅占0.07 %~0.11 %。孫凱寧等[13]研究顯示,山藥連作和輪作的土壤微生物占比排序為細菌>放線菌>真菌。雷鋒杰等[14]采用第二代高通量測序技術分析野生撫育模式、農田栽參模式和伐林栽參模式下人參根部的內生真菌多樣性,結果發現野生撫育模式下人參根部內生真菌的Chao1指數、ACE指數和Shannon指數均高于伐林栽參模式,農田栽參模式下這3種指數均最低,表明栽培模式會顯著影響人參根部內生真菌的形成及其多樣性。張紅霞等[15]進行連作山藥根際土壤和根莖高通量測序,結果顯示山藥根際土壤的細菌優勢群落有變形菌、綠彎菌、酸桿菌和放線菌,且土壤pH對細菌群落結構及分布具有重要影響。康捷等[16]對麻山藥不同時期根際土壤進行高通量測序也發現,麻山藥的根際土壤細菌多樣性和豐富度在苗期、花期和收獲期間無顯著差異,細菌的最優菌群是變形菌門和酸桿菌門,真菌的最優菌群是放射毛霉屬和被孢霉屬。劉元等[17]應用GSS高通量測序技術測定懷山藥根和莖的內生真菌,結果表明其根的內生真菌多樣性略低于莖,葡萄孢盤菌屬和鏈格孢屬分別為根和莖的優勢菌群。【本研究切入點】相比傳統的微生物平板培養法和生物標記法等土壤微生物研究方法,高通量測序技術具有覆蓋度更高的基因數據優勢,能更真實地反映土壤微生物的多樣性和豐富度,但目前以高通量測序技術對不同種植方式淮山根際土壤真菌多樣性進行分析的研究鮮見報道。【擬解決的關鍵問題】采用高通量測序技術,通過Illumina HiSeq測序平臺分析定向結薯栽培和袋料種植兩種方式下淮山根際土壤真菌群落的多樣性和分布規律,探明淮山袋料種植方式的土壤生態環境,為利用微生物多樣性指導淮山科學種植提供參考依據。
1 材料與方法
1.1 試驗材料
試驗地位于廣西南寧市隆安縣那桐鎮大滕村(東經107°52′22″,北緯22°58′11″),供試淮山品種為桂淮2號。試驗地前茬作物為香蕉,種植前測定土壤速效氮含量為131.2 mg/kg,速效磷含量為34.7 mg/kg,速效鉀含量為593.5 mg/kg,有機質含量為30.0 g/kg,pH 5.85。
1.2 試驗方法
1.2.1 試驗設計 兩種種植方式的淮山均于2017年4月21日種植。設定向結薯栽培和袋料種植2個處理,3次重復,共6小區。定向結薯栽培是在起壟的壟面上斜挖一條深約15.0 cm的種植溝,斜度與水平面約成25°角,在溝上鋪設淮山定向栽培專用模具(一種淮山種植的U型塑料薄膜槽,規格為長120.0 cm、寬7.5 cm、深3.5 cm),在鋪設的模具上覆土約3.0 cm后播種淮山種薯,使淮山塊莖膨大期的塊莖順著鋪設的模具定向生長(圖1);袋料種植是先在種植地上開一條深約25.0 cm、長1.2 m,斜度與水平面約成15°角的種植槽,在槽內鋪設經裁剪好的塑料薄膜(普通加厚塑料膜,長120.0 cm、寬50.0 cm),并用圓形鐵管或木條壓緊,抽出鐵管或木條后回填細碎松土,最后將塑料膜交疊成袋狀,并將肥料施在頭部喇叭口位置,即成袋料。種植時將種薯種在料袋頭部喇叭口處,并在種薯上覆土約8.0 cm,使淮山塊莖膨大期的塊莖沿著塑料膜斜向下生長(圖2)。

圖1 淮山定向結薯栽培法Fig.1 Schematic of conventional orientation planting method of yam
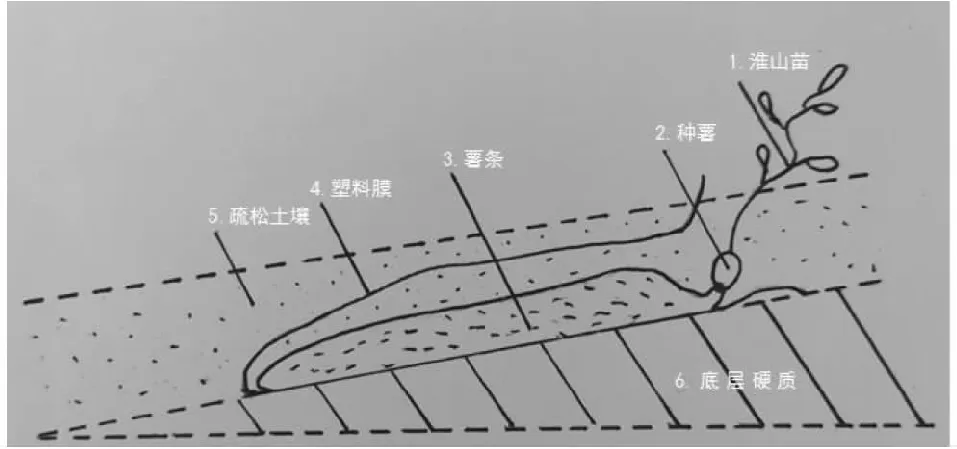
圖2 淮山袋料種植法Fig.2 Schematic of bag material planting method of yam
1.2.2 樣品采集 于收獲期(2018年4月15日)采集淮山根際20.0 cm深的土壤樣品,每小區按照S形選取6個點的土壤混勻合成1個土樣,淮山定向結薯栽培的土壤樣品標記為DX,袋料種植的土壤樣品標記為DL。采集的樣品置于50 mL無菌管,迅速放在液氮中帶回實驗室-80 ℃凍存,用于土壤微生物DNA提取和后續高通量測序。
1.2.3 土壤DNA提取及HiSeq測序 參照Soil DNA Kit 試劑盒說明提取土壤總DNA,根據所擴增的ITS1區域特點,基于Illumina HiSeq測序平臺,利用雙末端測序(Paired-end)方法,構建小片段文庫進行雙末端測序。通過對序列(Reads)拼接過濾和操作分類單元(OTU)聚類,進行物種注釋和豐富度分析。根據物種注釋結果,選取每個土壤樣品在不同分類水平上豐富度排名靠前的物種及其相對豐富度制圖;再對OTU進行Alpha多樣性指數等的統計和分析。樣品測序與分析委托北京諾禾致源科技股份有限公司完成。
1.3 統計分析
利用Uparse軟件[18]對樣品的全部Effective Tags進行聚類,以97 %的一致性(Identity)將序列聚類成為OTUs。使用Qiime Version 1.9.1[19]與Unit數據庫[20]進行物種注釋分析,并分別在門(phylum)、綱(class)、目(order)、科(family)、屬(genus)和種(species)分類水平統計各樣本的群落組成。使用Qiime Version 1.9.1計算Chao1、Shannon、Simpson、ACE指數和覆蓋度(coverage),以SAS 9.0進行方差分析。
2 結果與分析
2.1 土壤真菌的測序結果和多樣性指數
利用Illumina HiSeq測序平臺對ITS1區進行測序,從淮山定向結薯栽培和袋料種植方式的土壤樣品中得到用于構建操作OTU且獲得注釋信息的序列分別為83 457和78 991條,分別包含725和668個OUTs;兩種種植方式土壤樣品的測序覆蓋度均為99.90 %,表明測序深度包含了樣品中的絕大多數真菌,測序數據量合理。由表1可知,淮山定向結薯栽培和袋料種植的ACE指數為730.72和683.41、Chao1指數為720.87和679.48,Shannon為5.15和4.49,Simpson指數為0.91和0.88,表明袋料種植淮山土壤的真菌多樣性和豐富度均低于定向結薯栽培,但方差結果顯示,二者各指數間的差異不顯著(P>0.05,下同),即二者的真菌多樣性和豐富度相近。
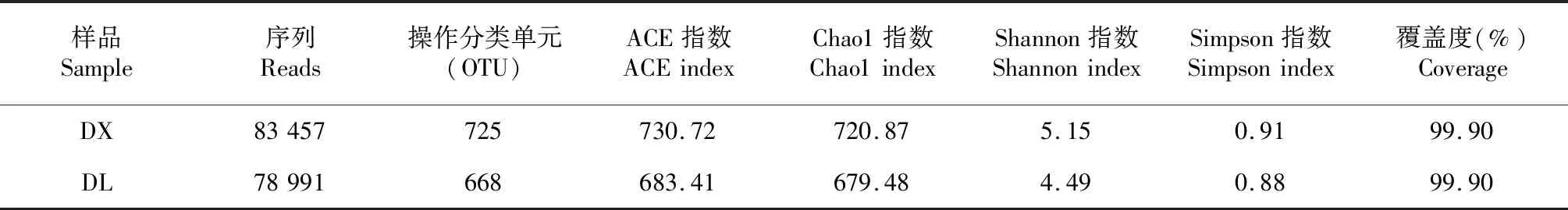
表1 淮山不同種植方式的土壤真菌高通量測序結果Table 1 Illumina HiSeq sequencing results of the fungal community in different planting methods of yam
2.2 在門水平上的土壤真菌群落豐富度分析
從圖3可看出,在兩種種植方式下,淮山根際土壤的真菌群落在門水平上主要有子囊菌門(Ascomycota)、擔子菌門(Basidiomycota)、接合菌門(Zygomycota)、壺菌門(Chytridiomycota)、球囊菌門(Glomeromycota)、新麗鞭毛菌門(Neocallimastigomycota)和其他門,其中,第一優勢門均為子囊菌門,其次均為擔子菌門;子囊菌門和擔子菌門在不同種植方式中的相對豐度總和均超過90.00 %(子囊菌門和擔子菌門在定向結薯栽培淮山土壤中的相對豐度分別為80.29 %和11.09 %,在袋料種植淮山土壤中的相對豐度分別為63.95 %和27.47 %)。方差分析結果顯示,在門水平上兩種種植方式土壤不同菌門的相對豐度均無顯著差異,說明不同淮山種植方式在門水平上未改變土壤真菌群落的相對豐度。
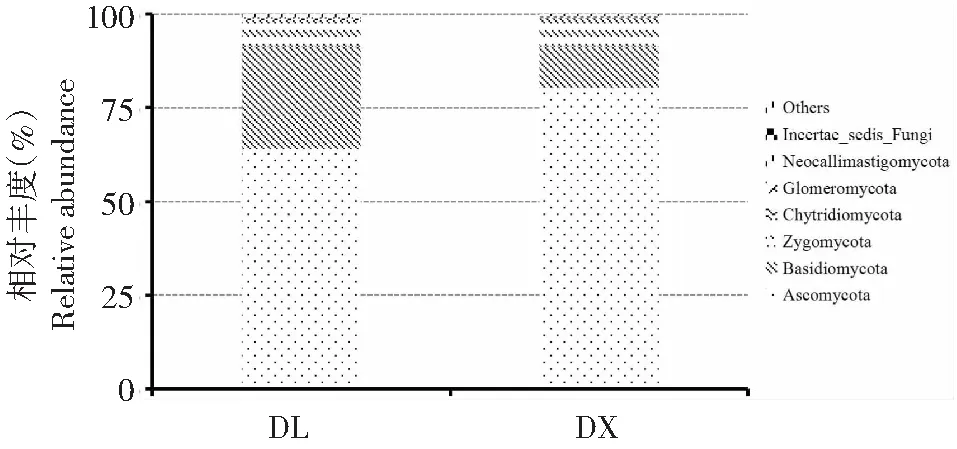
圖3 兩種種植方式淮山根際土壤真菌群落在門水平上的相對豐度Fig.3 Rhizosphere soil fungal relative abundance of yam on the phylum level of two planting methods
2.3 在綱水平上的土壤真菌群落豐富度分析
兩種種植方式淮山根際土壤真菌群落在綱水平上分為23個類群,排名前10位的有錘舌菌綱(Leotiomycetes)、糞殼菌綱(Sordariomycetes)、傘菌綱(Agaricomycetes)、Incertae-sedis-Zygomycota、散囊菌綱(Eurotiomycetes)、座囊菌綱(Dothideomycetes)、Incertae_sedis_Ascomycota、盤菌綱(Pezizomycetes)、球囊菌綱(Glomeromycetes)和節擔菌綱(Wallemiomycetes)(圖4)。其中,糞殼菌綱為定向結薯栽培淮山根際土壤真菌群落的第一優勢菌綱,相對豐度達36.37 %,其次為錘舌菌綱,相對豐度為31.14 %;而錘舌菌綱為袋料種植淮山根際土壤真菌群落的第一優勢菌綱,相對豐度為29.45 %,其次為糞殼菌綱,相對豐度為28.26 %。此外,袋料種植淮山根際土壤真菌群落的傘菌綱相對豐度也較高,為26.68 %。方差分析結果顯示,在綱水平上,兩種種植方式下的土壤真菌群落只有傘菌綱的相對豐度間均存在顯著差異(P<0.05),其他類群菌綱的相對豐度間差異不顯著;淮山袋料種植與淮山定向栽培土壤的真菌群落結構總體上相似,相對豐度只存在個別差異。
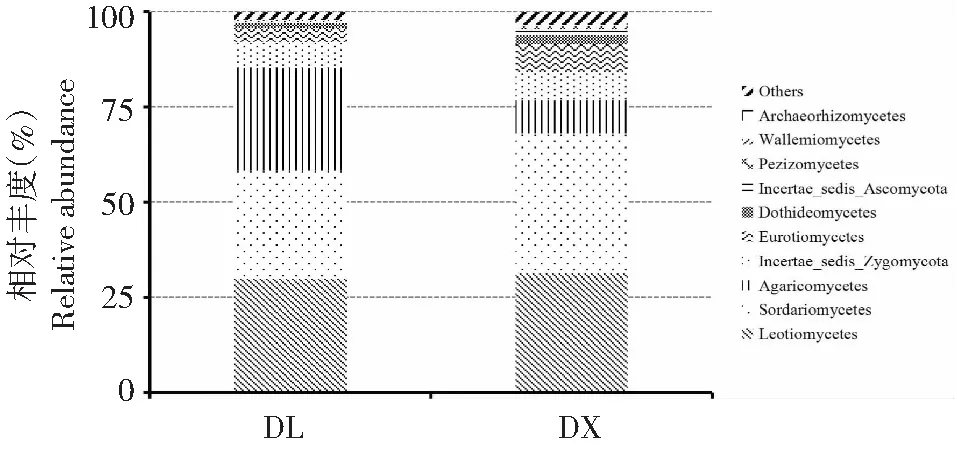
圖4 數量排名前10位的兩種種植方式淮山根際土壤真菌群落在綱水平上的相對豐度Fig.4 Top 10 of rhizosphere soil fungal relative abundance of yam on the class level of two planting methods
2.4 土壤真菌群落在目水平上的豐富度分析
兩種種植方式淮山根際土壤的真菌群落在目水平上均分為78個類群,排名前10位的有白粉菌目(Erysiphales)、肉座菌目(Hypocreales)、Geastrales、散囊菌目(Eurotiales)、糞殼菌目(Sordariales)、微囊菌目(Microascales)、被孢霉目(Mortierellales)、糙孢孔菌目(Trechisporales)、炭角菌目(Xylariales)和格孢腔菌目(Pleosporales)(圖5)。其中,第一優勢菌目均為白粉菌目,其次為肉座菌目;在定向結薯栽培方式中,白粉菌目和肉座菌目的相對豐度分別達30.44 %和23.14 %,在袋料種植方式中分別為28.78 %和21.02 %。方差分析結果顯示,在目水平上,兩種種植方式下的土壤真菌相對豐度間均無顯著差異,說明兩種淮山種植方法在目水平上未改變土壤真菌群落的相對豐度。
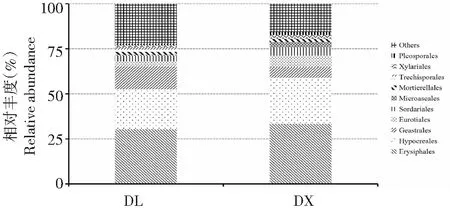
圖5 數量排名前10位的兩種種植方式淮山根際土壤真菌群落在目水平上的相對豐度Fig.5 Top 10 of rhizosphere soil fungal relative abundance of yam on the order level of two planting methods
2.5 在屬水平上的土壤真菌群落豐富度分析
在屬水平上,定向結薯栽培淮山根際土壤的真菌群落分為216個類群,袋料種植淮山的根際土壤真菌群落分為202個類群,排名前10位的有Microidium、鐮刀菌屬(Fusarium)、Nidulariopsis、帚枝霉屬(Sarocladium)、青霉菌屬(Penicillium)、頭束霉屬(Cephalotrichum)、Purpureocillium、粗糙孔菌屬(Trechispora)、Gibellulopsis和Cosmospora(圖6)。其中,兩種種植方式淮山根際土壤的第一優勢菌屬均為Microidium,第二優勢菌屬均為鐮刀菌屬,第三優勢菌屬均為Nidulariopsis;在定向結薯栽培方式的根際土壤真菌中,Microidium、鐮刀菌屬和Nidulariopsis的相對豐度分別為30.44 %、10.90 %和5.44 %,在袋料種植方式的根際土壤真菌中,Microidium、鐮刀菌屬和Nidulariopsis的相對豐度分別為28.78 %、14.62 %和11.89 %。方差分析結果顯示,在屬水平上,兩種種植方式下的土壤真菌相對豐度均無顯著差異,說明兩種淮山種植方法在屬水平上也未改變土壤真菌群落的相對豐度。
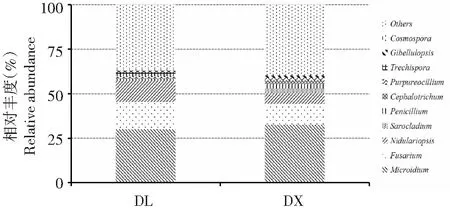
圖6 數量排名前10位的兩種種植方法淮山根際土壤真菌群落在屬水平上的相對豐度Fig.6 Top 10 of rhizosphere soil fungal relative abundance of yam on the genus level of two planting methods
3 討 論
子囊菌門真菌多數為腐生真菌,主要降解土壤中木質素和角質素等難降解的有機質[21],是土壤中真菌的優勢菌門[22],在各種土壤生態系統中進化速率較快[23];擔子菌門真菌多數為腐生或寄生真菌,也是土壤中的重要分解者,在土壤養分循環中發揮重要作用[24-26]。本研究中,淮山定向結薯栽培和袋料種植兩種種植方法土壤的子囊菌門和擔子菌門相對豐度總和均超90.00 %,說明兩種種植方式土壤中所含的優勢真菌類群能保障該種植方式下土壤的養分循環,淮山袋料種植方式具有可行性,可在旱坡地和石山地等非耕地種植;在屬水平上,淮山定向結薯栽培和袋料種植兩種種植方法土壤的第一優勢真菌屬為Microidium,是在感染白粉病的西印度醋栗根莖葉上被發現和命名并分類為白粉菌科的一個新的亞屬[27],但目前暫未查閱到有關Microidium特性的報道;第二優勢真菌屬為鐮刀菌屬,是一類全球性分布的真菌,可在土壤中越冬越夏,并侵染多種植物,使植物發生較難防治的枯萎病[28-29]和根腐病[30-32]等多種病害[33],主要侵染寄主植物維管束系統,破壞輸導組織維管束,并在代謝過程中產生毒素危害寄主作物,導致作物萎蔫死亡,影響作物品質和產量。已有研究表明,鐮刀菌是引起香蕉枯萎病[29]、山藥根腐病[30]、大蒜根腐病[31]和三七根腐病[32]等的主要菌類。譚雪蓮等[34]、蔡柏巖等[35]研究顯示,鐮刀霉菌等土傳病原真菌繁殖增多會抑制土壤細菌活性,降低各種土壤酶活性,最終導致土壤環境惡化,表明病原菌屬的微小變化可能會破壞真菌群落原有結構的平衡,從而導致淮山真菌病害發生。本研究結果表明,淮山兩種種植方式下土壤真菌的第二優勢真菌屬均為鐮刀菌屬,說明兩種種植方式種植的淮山均有可能發生根腐病,而袋料種植方式土壤鐮刀菌屬的相對豐度高于定向結薯栽培土壤,表明袋料種植的淮山感染根腐病風險高于定向結薯栽培。在實際生產中,可利用微生物菌劑改善土壤微生態環境,做好根腐病的預防工作,以保證袋料種植淮山的產量和品質。
本研究采用高通量測序技術揭示兩種種植方式下淮山根際土壤真菌群落的多樣性,發現不同淮山種植方式在門、綱、目和屬水平上均未改變其根際土壤真菌群落的相對豐度(除傘菌綱外),表明兩種淮山種植方式下土壤真菌的群落組成相似,淮山袋料種植方式可行,推廣應用前景可期。
4 結 論
在不同分類水平上,淮山袋料種植方式中土壤根際真菌的優勢類群與定向結薯栽培方式基本相同,表明淮山袋料種植方式具有可行性;袋料種植方式土壤中的鐮刀菌屬相對豐度高于定向結薯栽培方式土壤,可作為改善淮山袋料種植土壤生態環境的參考依據。
