4種臨床抗結核藥物對A549細胞TLRs信號通路和炎癥因子表達的影響①
2021-05-26 06:13:50李文靜胡育龍高蔚豐
中國免疫學雜志 2021年6期
關鍵詞:信號
王 娟 程 龍 李文靜 胡育龍 曾 瑾 李 敏 高蔚豐 李 勇
(寧夏大學西部生物資源保護與利用教育部重點實驗室,銀川750021)
結核病(tuberculosis,TB)是一種慢性、消耗性人獸共患傳染病,由結核分枝桿菌(mycobacterium tuberculosis,MTB)單一傳染性病原體感染引起[1]。目前TB已成為全球十大死因之一,死亡率高于包括HIV在內的單一傳染病[2]。我國每年TB新發病例約為90萬,是全球第三大TB高負擔國家[3]。根據疾病預防控制局發布數據顯示,2018年我國現有肺結核患者82.33萬例,死亡病例3 149例,報告發病率和報告死亡率均位居乙類傳染病第二。對于細菌感染性TB臨床常采用抗生素治療,但常伴有MTB耐藥問題[4]。目前臨床治療肺結核主要采用抗生素組合療法,但多種抗生素長期使用導致機體免疫相關細胞相互作用,進而影響機體抗結核免疫反應[5]。
MTB是典型的胞內寄生菌,肺泡巨噬細胞作為免疫調節細胞和效應細胞,在感染過程中通過吞噬、抗原提呈和分泌多種細胞因子等功能調控機體免疫應答和炎癥反應[6]。由于感染巨噬細胞的能力對細菌在宿主體內的傳播和擴散至關重要,既往抗結核免疫機理研究中,結核菌與肺泡巨噬細胞的相互作用一直是研究重點和熱點[7]。但近年研究發現,肺泡Ⅱ型上皮細胞(alveolar epithelial typeⅡcells,AECⅡ)與巨噬細胞類似,也是MTB感染的主要靶細胞,是由呼吸道吸入TB病原菌最先感染的細胞[8]。AECⅡ細胞不僅可通過Toll樣受體家族(TLRs)、表面活性蛋白(SPs)和Dectin-1等調控宿主先天免疫反應,抵御外源病原菌,并可產生炎癥因子、趨化因子和細胞因子等激活獲得性免疫反應,募集和活化吞噬細胞至感染部位清除病原菌,是阻止結核菌入侵宿主的第一道細胞防線,在抗MTB感染免疫調節中的作用備受關注[9-10]。
目前臨床常用抗結核藥物主要有一線抗結核藥物、二線抗結核藥物、新型抗結核藥物及固定劑量復方制劑[11]。一線抗結核藥物主要包括異煙肼(isoniazid,INH)、利福平(rifampin,RFP)等,抗菌活性強、副作用少[12];二線類藥物包括喹諾酮類、大環內酯類、氨基糖苷類、對氨基水楊酸(para-aminosalicylic acid,PAS)類等,用于治療一線類藥物不耐受及耐藥性TB患者[11]。近年體內和體外試驗證實,部分抗結核藥物不僅具有殺菌作用,還具有免疫調節作用,如大環內酯類藥物通過促進中性粒細胞活化和提高免疫細胞功能增強機體防御能力[13];INH可增強小鼠腹腔巨噬細胞酸性磷酸酶活性及吞噬能力[14];RFP在體外可強烈抑制由有絲分裂原和特異性抗原誘導的致敏和非致敏小鼠脾淋巴細胞轉化,且與濃度及時間呈正相關[15]。但抗結核藥物應用過程中會出現過敏反應、血液系統毒性等不良反應,是否會對機體免疫相關細胞的免疫功能,尤其是抗結核先天免疫調控功能產生影響有待進一步研究[16]。因此,本研究采用不同濃度的一線抗結核藥RFP、INH和二線抗結核藥硫酸阿米卡星(amikacin sulfate,AMK)、PAS處理AECⅡA549細胞,分析不同抗結核藥物對A549細胞中TLRs信號通路信號分子和炎癥因子表達的影響,探討抗結核藥物對AECⅡ抗結核先天免疫功能的影響,為臨床TB治療提供理論參考。
1 材料與方法
1.1 材料 人肺腺癌細胞株A549由寧夏大學西部特色生物資源保護與利用重點實驗室保存提供;RFP注射液購自沈陽雙鼎制藥有限公司;INH注射液購自天津金耀藥業有限公司;AMK注射液購自蘇州第壹制藥有限公司;PAS粉針劑購自貴州光正制藥有限責任公司;TRIzol試劑購自美國Invitrogen公司;DMEM高糖培養基、FBS購自美國Gibco公司;反轉錄和qRT-PCR試劑盒均購自北京全式金生物技術有限公司;全蛋白提取試劑盒、BCA蛋白定量試劑盒和SDS-PAGE凝膠配制試劑盒購自南京凱基生物;一抗TLR2(CST#12276)、NF-κB p50(CST#135 86)和MyD88(CST#4283)購自Cell signal technology公司;一抗β-actin(TA-09)和二抗羊抗兔IgG-HRP(ZB-2301)購自北京中杉金橋;Plus-ECL化學發光試劑盒購自美國PerkinElmer;人IL-8和IL-12 p70購自美國R&D公司。
1.2 方法
1.2.1 A549細胞培養 A549細胞接種于含10%FBS和青鏈霉素雙抗的DMEM高糖培養液中,37℃、5%CO2恒溫培養,細胞密度達到70%~80%時,0.25%胰蛋白酶-EDTA消化,收集培養細胞。
1.2.2 給藥方式及分組 A549細胞以4×105個/孔接種于6孔板,培養箱中培養過夜至細胞密度達到60%時,更換新鮮培養液繼續培養24 h。參考各抗生素說明中的血藥代謝濃度分組:Control組(只加等量溶劑)、不同濃度RFP組(1.00、5.00、10.54μg/ml),不同濃度INH組(1.00、5.00、10.00μg/ml),不同濃度AMK組(2.00、16.00、33.38μg/ml),不同濃度PAS組(10.00、100.00、213.00μg/ml)。
1.2.3 細胞總RNA提取和cDNA合成 胰蛋白酶消化處理24 h的各組細胞,PBS洗滌3次,加入TRIzol,按照TRIzol試劑說明書提取細胞總RNA,超微量分光光度計測定RNA濃度及純度,瓊脂糖凝膠電泳檢測RNA完整性。將提取的總RNA反轉錄為cDNA,反應體系20μl:總RNA 5μg,Oligo(dT)181.0μl,2×TSReaction Mix 10μl,TransScript RT/RI Enzyme Mix 1.0μl,gDNA Remover 1.0μl,添 加RNase-free Water至20μl,42℃15min,85℃5s終止反應。
1.2.4 qRT-PCR 檢測根據人TLR2、TLR4、NFκB、MyD88、IL-6、IL-8、IL-12A基因序列,通過Primer Premier 5.0軟件設計引物,以β-actin為內參并設計引物,引物由上海基康生物技術有限公司合成,序列見表1。qRT-PCR反應體系20μl:cDNA 1.0μl,上下游引物(10μmol/L)1.0μl,2×Tip Green qPCRSuperMix 10μl,H2O 7μl。Quant Studio 5熒光定量PCR儀上進行qRT-PCR反應,反應條件:94℃預變性30 s,94℃變性5 s,60.8℃退火15 s,72℃延伸10 s,40個循環,2-ΔΔCt法分析數據。
1.2.5 Western blot 按照全蛋白提取試劑盒說明書提取總蛋白,BCA蛋白定量試劑盒測定蛋白濃度,根據不同蛋白相對分子質量大小配制不同濃度SDS-PAGE凝膠,取相同量蛋白樣品進行電泳(恒壓80 V),轉膜(濕轉,恒流300 mA);5%脫脂奶粉室溫封閉1~2 h,加入一抗4℃孵育過夜,次日洗膜,加入二抗孵育,洗膜,化學發光試劑發光并采用化學發光檢測儀成像。
1.2.6 IL-8及IL-12 ELISA檢測 收集各組培養細胞上清,按照ELISA試劑盒說明書進行操作。
1.3 統計學處理 采用SPSS19.0軟件進行統計學分析,所有實驗重復3次,數據以±s表示,數據比較采用t檢驗,Graph Pad Prism7.0軟件繪圖。
2 結果
2.1 A549細胞培養與總RNA提取 A549細胞為貼壁生長的上皮細胞,為多角形,形態隨密度變化略有差異。本實驗選取對數生長期生長狀態良好的細胞進行后續實驗(圖1)。提取各實驗組處理24 h后的A549細胞總RNA,1%瓊脂糖凝膠電泳檢測結果見圖2,各組28 S rRNA、18 S rRNA和5 S rRNA條帶完整,且RNA純度(A260/A280)為1.9~2.1,符合后續實驗要求。

表1 qRT-PCR引物序列Tab.1 Primer sequences for qRT-PCR
2.2 抗結核藥物對A549細胞TLRs信號通路相關分子mRNA表達的影響 RFP各濃度組TLR2和TLR4受體mRNA表達顯著高于對照組(P<0.05或P<0.01),且均隨濃度增加而升高。各組下游接頭分子MyD88表達顯著降低,轉錄因子NF-κBmRNA表達僅在RFP濃度為10.54μg/ml時顯著降低(P<0.05),其余濃度RFP對NF-κB表達影響無統計學意義(圖3A),提示RFP雖誘導較高水平的TLR2、TLR4表達,但MyD88、NF-κB表達并未被激活,反而受到抑制,總體表現為對TLRs信號通路的抑制作用。INH各劑量組TLR2、TLR4表達與對照組差異顯著(P<0.01),TLR2表達與濃度呈負相關;MyD88、NF-κB表達變化與TLR2類似,與濃度呈負相關,其中INH濃度為10.00μg/ml時,MyD88表達明顯降低,NF-κB表達在INH濃度為1.00μg/ml時顯著升高(P<0.05,圖3B),提示INH可抑制A549細胞TLR2信號通路。AMK各組A549細胞TLR2、TLR4表達顯著升高(P<0.05),2.00μg/ml升高最為顯著,且NF-κBmRNA表達僅在AMK濃度為2.00μg/ml時顯著提高(P<0.05),但AMK處理對MyD88表達無顯著影響(圖3C),表明隨濃度增加,AMK對TLR2、TLR4信號通路的活化作用逐漸減弱。PAS可顯著提高TLR2、TLR4表達,且呈一定的濃度依賴性;MyD88和NF-κB表達僅在PAS濃度為100.00μg/ml時顯著降低(P<0.05,圖3D)。
圖1 A549細胞生長狀態(×100)Fig.1 Growth status of A549 cells(×100)
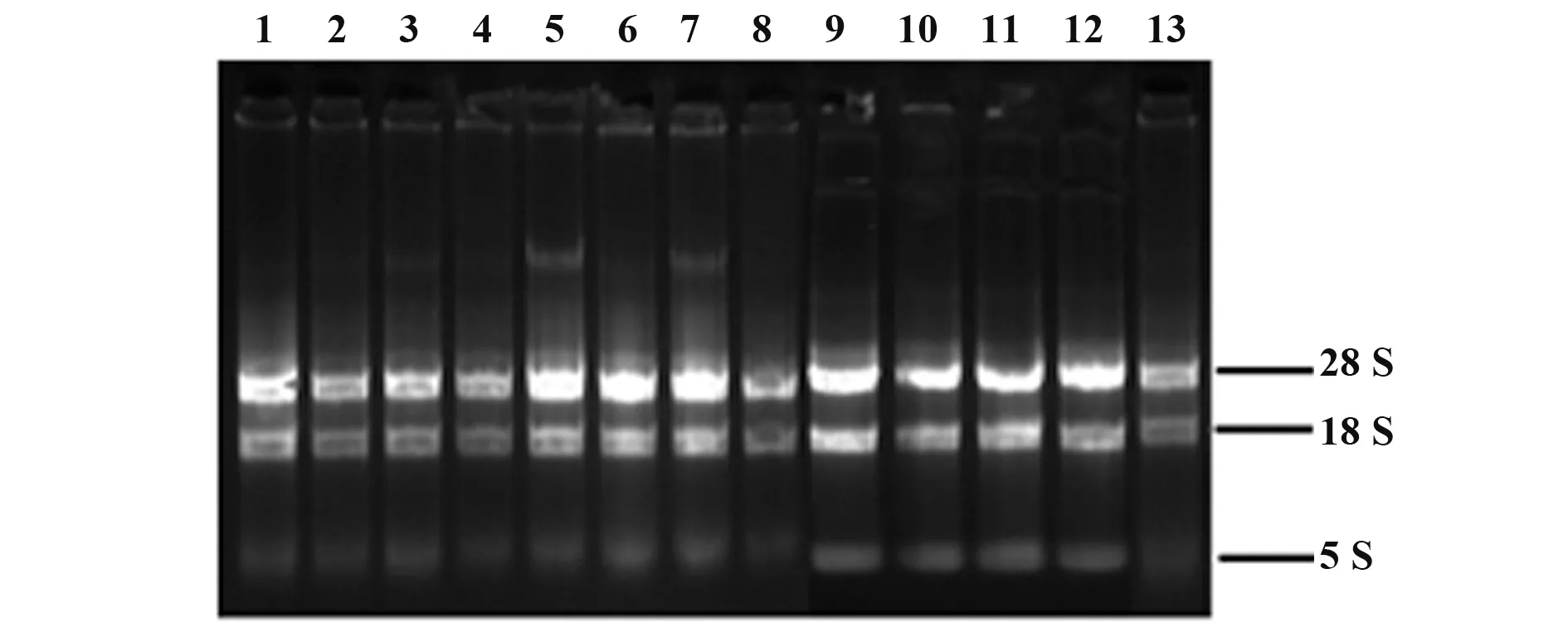
圖2 提取的A549細胞總RNA瓊脂糖凝膠電泳分析Fig.2 Gel electrophoresis of total RNA extracted from A549 cell
2.3 抗結核藥物對A549細胞TLRs信號通路相關分子蛋白表達的影響 TLR2、MyD88和NF-κB p50蛋白表達在RFP濃度為1.00μg/ml、5.00μg/ml時均有一定增加,其中5.00μg/ml RFP組TLR2表達顯著升高(P<0.05);當RFP濃度達到10.54μg/ml時,3種蛋白表達水平基本與對照組相同,NF-κBp50表達有所下調,說明高濃度RFP處理對A549細胞中TLR2信號通路有抑制作用(圖4)。INH對A549細胞中TLR2和MyD88表達的影響具有一致性,但對NF-κB p50表達的影響卻與前兩者正好相反(圖4B),表明INH可能通過其他途徑實現對NF-κBp50的活化。2.00μg/ml AMK處理顯著降低TLR2、MyD88表達(P<0.05),但隨著AMK濃度增加,兩種蛋白表達均有所提高;各濃度組NF-κB p50表達與對照組相比都有一定上調(圖4C),提示AMK可激活A549細胞中TLR2信號通路。TLR2、MyD88表達在PAS各濃度組均高于對照組,NF-κB p50表達隨PAS濃度增加而升高,說明PAS可活化A549細胞中TLR2信號通路(圖4D)。
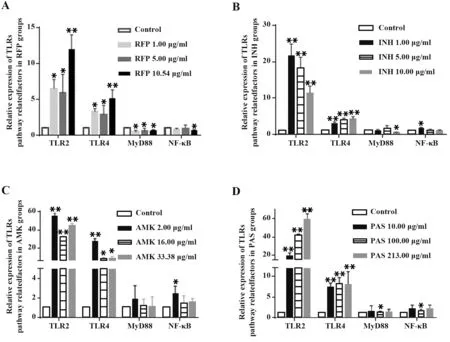
圖3 抗結核藥物對A549細胞TLRs信號通路相關分子mRNA表達的影響Fig.3 Effect of anti-tuberculosis drugs on mRNA expression of signal molecules in TLRs signaling pathway in A549 cells
2.4 抗結核藥物對A549細胞IL-8、IL-12amRNA表達的影響 隨RFP處理濃度增加,IL-8表達變化無統計學意義,IL-12amRNA表達顯著降低(P<0.05),3個濃度組差異無統計學意義(圖5A)。INH各濃度組IL-8、IL-12a表達顯著高于對照組(P<0.05或P<0.01),5.00μg/ml時變化最為顯著(圖5B)。AMK處理對IL-8表達無顯著影響,但各實驗組IL-8表達與濃度呈負相關,僅2.00μg/ml AMK處理對IL-12amRNA表達有顯著上調作用(P<0.05),其余2組IL-12a表達變化無統計學意義。AMK各濃度組IL-12a表達同樣與濃度呈負相關(圖5C),僅100.00μg/ml PAS組A549細胞IL-8、IL-12amRNA表達顯著上調(P<0.05,圖5D),表明4種抗結核藥物作用于A549細胞后,僅RFP對炎癥因子IL-12a表達有抑制作用,其余各抗結核藥物對炎癥因子IL-8和IL-12amRNA表達影響無統計學意義。
2.5 ELISA法測定IL-8、IL-12含量 隨RFP濃度增加,IL-8含量下降;與對照組相比,1.00μg/ml RFP組IL-12含量顯著升高(P<0.05),而5.00μg/ml RFP組IL-12含量卻顯著降低(P<0.05),10.54μg/ml RFP組IL-12含量變化無統計學意義(圖6A),表明RFP在一定程度上抑制A549細胞中IL-8、IL-12產生。INH可不同程度提高IL-8及IL-12含量,5.00μg/ml時作用最強(P<0.05,圖6B)。AMK處理A549細胞,IL-8含量在2.00μg/ml、16.00μg/ml時顯著增加(P<0.01),33.38μg/ml AMK組IL-8含量與對照組差異無統計學意義;當AMK濃度為2.00μg/ml時,IL-12含量變化無統計學意義,AMK濃度為16.00μg/ml、33.38μg/ml時,IL-12含量顯著降低(P<0.01,圖6C),提示AMK對A549細胞炎癥因子分泌的抑制作用具有劑量依賴性。與對照組相比,PAS各濃度組A549細胞IL-8、IL-12含量明顯上升(P<0.01),IL-12含量與劑量呈負相關(圖6D)。
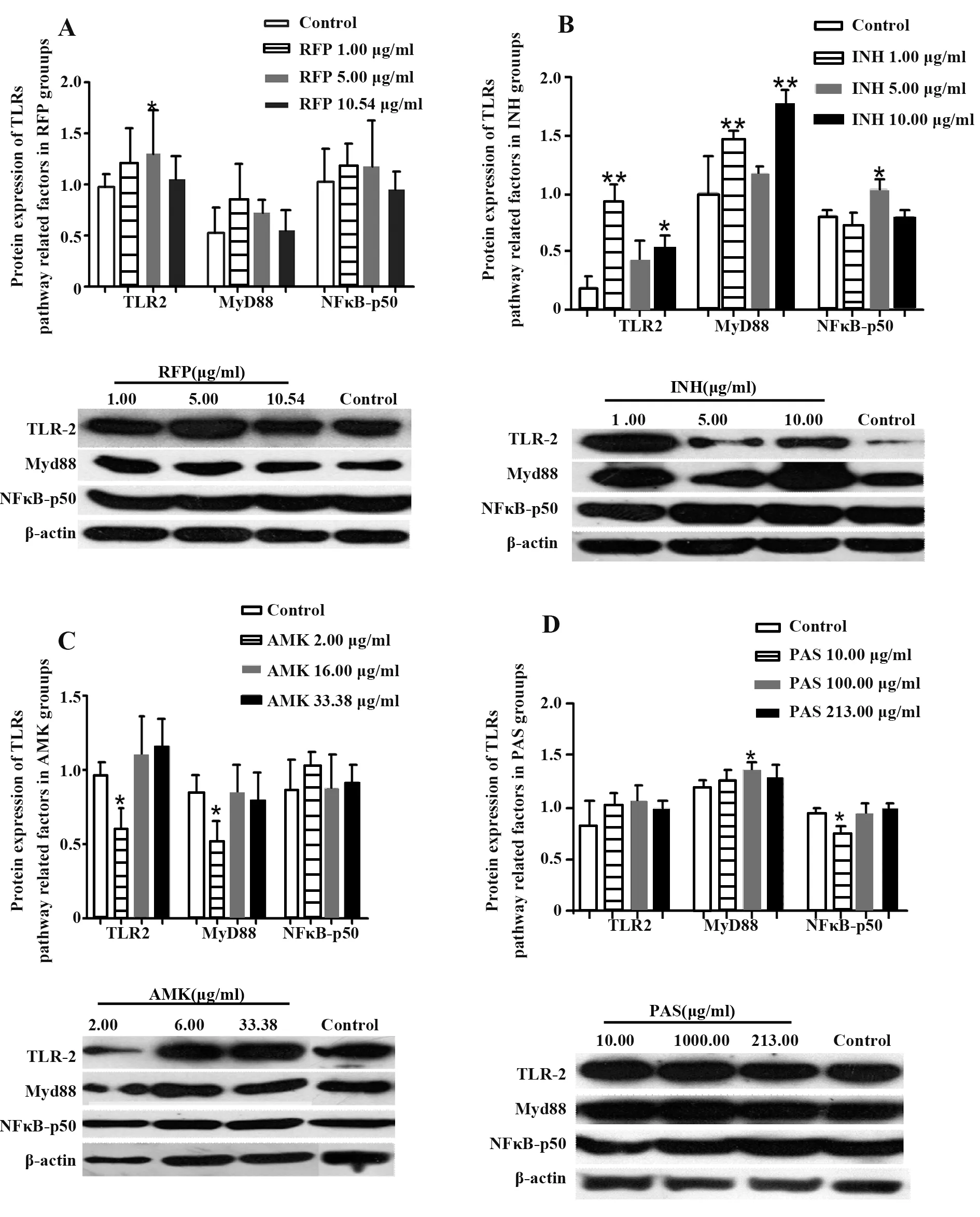
圖4 抗結核藥物對A549細胞中TLRs信號通路相關分子蛋白表達的影響Fig.4 Effect of anti-tuberculosis drugs on protein expressions of signal molecules in TLRs signaling pathway in A549 cells
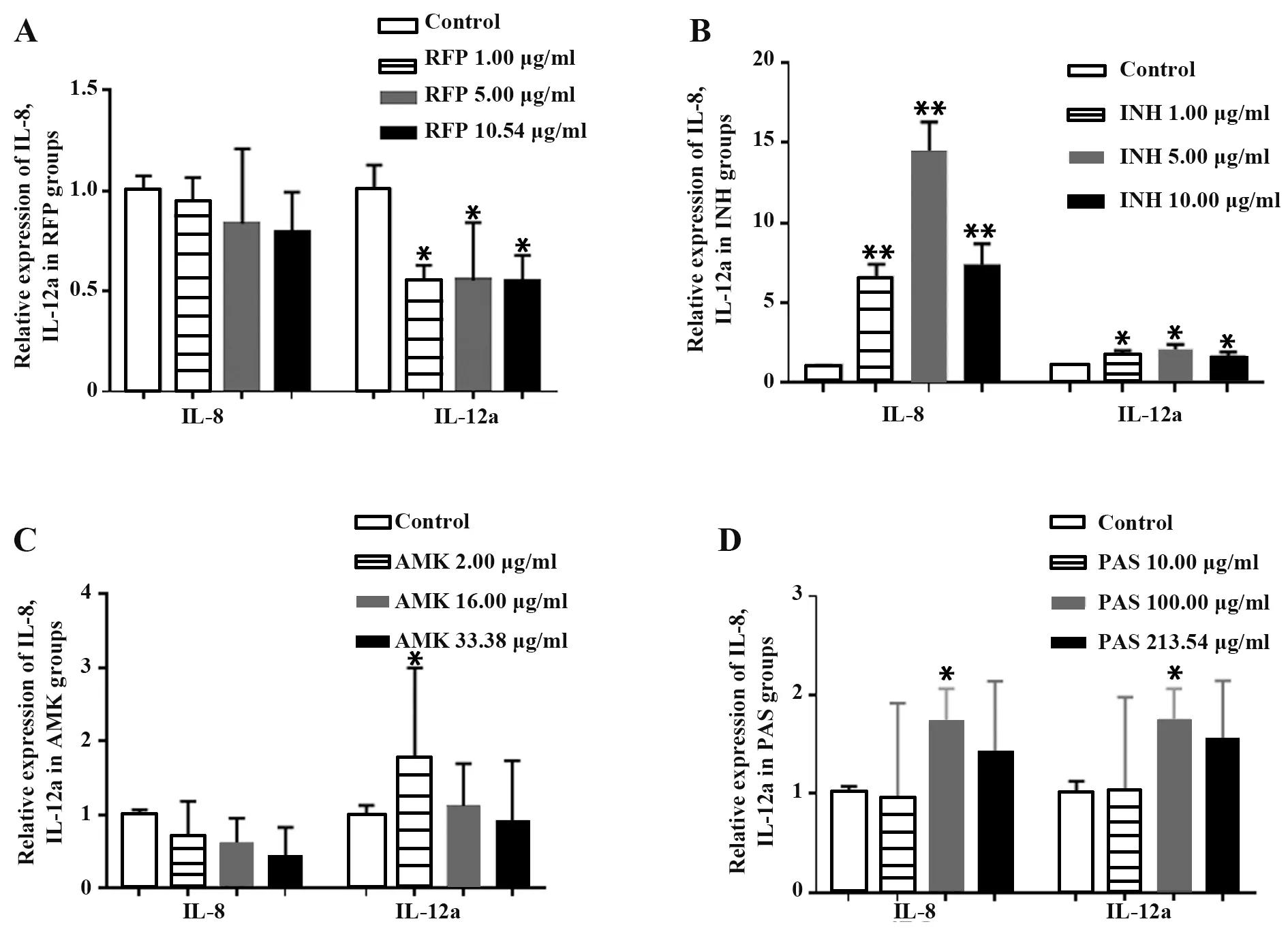
圖5 抗結核藥物對A549細胞IL-8、IL-12a mRNA表達的影響Fig.5 Effect of anti-tuberculosis drugs on expressions of IL-8 and IL-12a mRNA of A549 cells
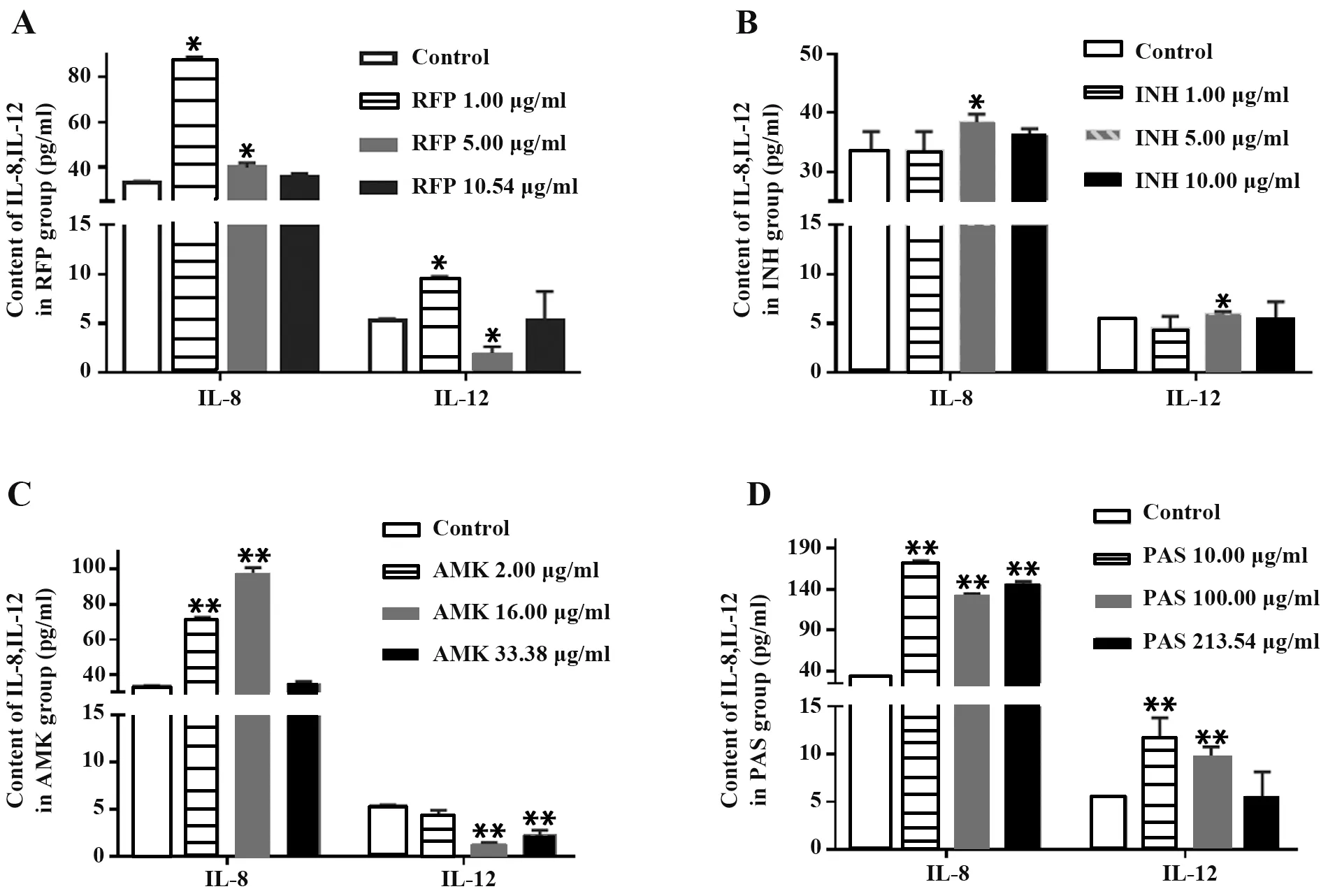
圖6 抗結核藥物對A549細胞IL-8、IL-12分泌的影響Fig.6 Effect of anti-tuberculosis drugs on IL-8 and IL-12 production in A549 cells
3 討論
近年耐藥尤其是耐多藥TB流行呈逐年上升趨勢,而結核藥物研發相對較慢,導致TB治療形勢日趨嚴峻。隨著研究深入,除對現有抗結核藥物的抗菌機制有了更全面透徹地認識外,還發現部分抗結核藥物具有免疫調節作用。抗結核藥物的免疫調節作用呈雙向性,部分藥物可改善宿主免疫功能,協同抗菌作用,但部分藥物卻表現出免疫抑制效應[17]。因此,深入了解抗結核藥物的免疫調節作用,與抗菌活性相結合,將為TB臨床治療提供理論參考。課題組研究了具有代表性的一線抗結核藥物RFP、INH及二線抗結核藥物AMK、PAS對A549細胞TLRs通路相關信號分子和炎癥細胞因子表達的影響,其中,RFP是利福霉素衍生物,是最有效的抗結核抗生素之一[15];INH抗菌活性較強,可有效殺滅細胞內外MTB,是目前TB治療的首選藥物[12];AMK常與INH、RFP等聯用,是肺結核治療的二線注射首選藥物[18];PAS是繼鏈霉素之后發現的第二種TB治療藥物。
TLRs是由先天免疫系統細胞表達的一種跨膜蛋白,與機體免疫和炎癥反應密切相關,是連接天然免疫和獲得性免疫的橋梁,在機體抵抗病原微生物免疫反應中扮演重要角色[19-21]。機體感染MTB后會激活TLRs信號通路,TLR2、TLR4和TLR9是機體抗MTB的主要炎癥受體[21]。活化的TLRs可促進炎癥因子、趨化因子、黏附分子及其受體等多種靶基因轉錄與表達,引發炎癥反應[22]。既往研究表明,AECⅡ通過TLR2/TLR4二聚體識別MTB[23]。本試驗qRT-PCR結果也顯示TLR2和TLR4的mRNA表達變化趨勢基本一致。本研究采用RFP處理AECⅡ,當濃度達為10.54μg/ml時,可抑制A549細胞TLRs受體信號通路活化。INH干預雖對A549細胞TLR2和MyD88表達的影響一致,但對NF-κB表達的影響與前兩者相反,即TLR2和MyD88表達降低的同時NF-κB表達升高,表明INH抑制TLR2信號通路的同時,可經TLR4通過MyD88非依賴性通路活化NF-κB。MyD88非依賴通路可調控樹突狀細胞成熟及免疫調節因子(如MHC、CD4+等)表達調節獲得性免疫反應[21]。因此,INH可能具有增強機體免疫功能的作用,與既往研究結論一致。INH可通過提高大鼠外周血CD3+T細胞、CD4+T細胞比例及CD4+/CD8+比值有效增強免疫功能[24]。毛澤善等[14]發現INH可增強小鼠非特異性免疫功能。兩種二線抗結核藥物AMK、PAS均可上調TLR2、TLR4、MyD88和NF-κB表達,表明AMK、PAS可激活A549細胞TLR2、TLR4信號通路。
炎癥細胞因子表達水平可反映局部炎癥損傷程度。適量細胞因子分泌有利于機體清除病原體,但過度分泌可能導致炎癥失控及加重局部組織損傷[25]。IL-8是趨化性細胞因子,其主要功能是趨化中性粒細胞、T淋巴細胞等多種免疫細胞至感染部位吞噬、殺傷病原菌,并進一步導致細胞脫顆粒,引發呼吸爆發等;此外,炎癥反應早期始發因子通過激活其他炎癥細胞因子引起炎癥反應[26]。IL-8參與慢性支氣管炎、胃潰瘍、胰腺炎等炎癥性疾病發生、發展和預后過程中,在感染控制及病理損傷形成中起重要作用。IL-12作為一種多功能細胞因子,被認為是調節免疫反應最重要的細胞因子之一,在機體抗MTB感染過程中,可誘導Th0向Th1分化,并維持Th1效應功能,刺激NK細胞、巨噬細胞和中性粒細胞增殖并提高其靶細胞殺傷活性,此外還是IFN-γ分泌的強誘導劑,是天然免疫和獲得免疫的調節因子[27-33]。從炎癥細胞因子mRNA表達和分泌情況來看,隨RFP濃度增加,IL-8、IL-12表達、分泌均有一定程度的降低,對IL-12的影響最為顯著。表明RFP可通過抑制TLRs信號通路降低炎癥因子表達,具有免疫抑制功能,與RFP可有效降低復治涂陽肺結核患者機體TNF-α、CRP、IL-6、IL-10等炎癥因子水平、促進機體免疫功能恢復的研究結論一致[34]。因此,RFP在減輕局部炎癥反應強度、減輕組織損傷方面起重要作用。INH可不同程度提高IL-8及IL-12表達與分泌,5.00μg/ml時效果最強,說明INH具有免疫激活作用。感染引起的機體炎癥反應是一把“雙刃劍”,適當調節可協助機體殺滅MTB,調節不當則會導致機體組織損傷[33]。因此,采用INH治療TB時,為減輕炎癥反應,可與具有抑制炎癥因子表達作用的抗結核藥物聯用,也從免疫調節作用的角度,解釋了為何INH與RFP聯用是目前公認的最佳治療方法。隨AMK濃度增加,IL-8、IL-12 mRNA表達及分泌先升高后降低,說明AMK在激活TLRs信號通路促進IL-8、IL-12產生的同時,還通過其他途徑調控炎癥因子表達,從而抑制炎癥反應,即AMK存在免疫激活和免疫抑制兩方面作用。AMK為第三代氨基糖苷類抗生素,關于氨基糖苷類抗生素免疫調節作用的報道存在免疫激活和免疫抑制兩方面,如慶大霉素既可抑制CD3+、CD56+、CD8+等免疫細胞數量,起免疫抑制作用,又可通過抑制中性粒細胞的凋亡提高機體免疫功能[35-36]。蔡少華等[37]研究證實AMK具有免疫抑制作用,但AMK通過何種信號轉導通路抑制炎癥因子合成有待進一步研究。PAS可促進IL-8和IL-12表達與分泌,具有免疫激活功能,但隨濃度增加,免疫激活作用減弱,IL-12含量降低,提示PAS的免疫調節作用可能具有多重性。PAS為磺胺類抗菌藥物及其增效劑,磺胺類藥物屬于抑制免疫功能的抗菌藥物,但多呈現出對中性粒細胞吞噬作用的抑制作用,如在炎癥性腸病研究中發現PAS對黏膜IL-1β合成與釋放無影響[38]。本研究中PAS呈現免疫激活作用。目前對PAS的研究較少,其對機體免疫功能的影響及機制有待進一步研究。
綜上所述,4種抗結核藥物對AECⅡA549細胞的免疫功能具有一定影響。RFP通過抑制TLRs信號通路降低炎癥因子表達,減輕炎癥反應;INH可通過激活MyD88非依賴性通路促進炎癥反應及增強免疫功能;AMK在激活TLRs通路的同時,又通過其他通路抑制炎癥因子表達,呈現出免疫調節功能的多重性;PAS可激活TLRs信號通路促進IL-8、IL-12表達,促進A549細胞免疫應答,為TB臨床治療提供了理論參考。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06