煙草bZIP基因家族的鑒定及其在不同成熟度煙葉中的表達分析
2021-06-05 07:46:28楊佳翰張炳輝顧鋼楊玄松秦明月謝小芳
中國煙草學報 2021年2期
關鍵詞:煙草
楊佳翰,張炳輝,顧鋼,楊玄松,秦明月,謝小芳*
1 福建農林大學生命科學學院,福州 350002;
2 福建省煙草專賣局煙草科學研究所,福州 350003;
3 福建省作物設計育種重點實驗室,福州 350003
轉錄因子是一類重要的調控蛋白,它能與特定DNA序列(順式作用元件)結合,從而激活或抑制特定基因的轉錄表達[1]。堿性亮氨酸拉鏈轉錄因子(Basic region/leucine zipper,bZIP)是真核生物中廣泛存在的一個重要基因家族[2]。bZIP轉錄因子具有一個長度約為60~80個氨基酸的保守bZIP結構域,該結構域由堿性區(qū)和亮氨酸拉鏈區(qū)組成[3],其中,堿性區(qū)高度保守,長約16個氨基酸,包含一個核定位信號和一個識別并結合啟動子特定核酸序列的保守N-X7-R/K基序;亮氨酸拉鏈區(qū)的保守性較低,靠近堿基區(qū)的C端,它由多個七肽重復序列組成,每七個氨基酸的第七位含有一個亮氨酸,亮氨酸具有被其他疏水性氨基酸所取代的特性,如異亮氨酸、纈氨酸、蛋氨酸等。該區(qū)域通過兩個bZIP轉錄因子之間的疏水力纏結形成同源或異源二聚體疏水性氨基酸,參與bZIP蛋白與DNA結合前的二聚反應。
迄今為止,已有許多真核生物的bZIP基因家族成員在全基因組范圍內被成功鑒定。例如:擬南芥(75個)[4],水稻(89個)[3],玉米(125個)[5],蓖麻豆(100個)[6],蘋果(114個)[7],甘薯(41個)[8],番茄(69個)[9]和大豆(131個)[10]等植物。已有研究表明:bZIP轉錄因子參與種子萌發(fā)過程、幼苗的產生和形成、花芽分化和花器誘導等[11-13]。與此同時,bZIP轉錄因子廣泛參與了鹽脅迫,干旱脅迫,冷脅迫等非生物脅迫響應和生物脅迫防御等生物學過程。例如,擬南芥AtbZIP17對鹽脅迫響應明顯,可以直接或間接地激活鹽脅迫應答基因的表達,對提高植物的耐鹽性具有重要作用[14];在水稻中,干旱處理下OsbZIP20和OsABA45的表達都大幅上調,過表達OsbZIP71可顯著提高水稻的抗旱性[15-16];在低溫脅迫中,Liu等[17]在水稻中鑒定出8 個與苗期低溫抗性相關的bZIP 基因(OsbZIP08、OsbZIP35、OsbZIP38、OsbZIP46、OsbZIP63、OsbZIP72、OsbZIP73和OsbZIP76);此外,小麥TabZIP1[18],辣椒CabZIP1等基因參與了病原菌侵染的防御響應[19-20]。在煙草中,Kusano等鑒定了一個受低溫誘導的bZIP基因(tbz17)[21],Yang等[22]研究發(fā)現(xiàn)tbz17及其家族成員tbzF與煙葉的衰老緊密相關,這兩個基因在衰老的煙葉中大量表達,其轉錄本主要位于衰老葉片中的保衛(wèi)細胞和微管組織中。另外,Kuhlmann等[23]發(fā)現(xiàn)煙草bZIP 轉錄因子BZI-1在煙草的病原防御響應和生長素介導的信號轉導中具有重要作用。
煙草是我國的重要經濟作物之一。煙草收獲的主要器官是煙葉,因此葉片的生長發(fā)育、葉色轉換等非常重要。煙葉的成熟衰老常常被當作一種逆境,其實質是各類細胞的程序性死亡,從而適應外界環(huán)境改變帶來的生存脅迫,在基因調控下細胞自主選擇終結生命歷程的過程[24-25]。一些轉錄因子會激活或抑制逆境基因的表達,從而影響植物的抗逆性和生長發(fā)育[26-29]。大量研究證明:bZIPs家族廣泛參與植物的生長發(fā)育、果實成熟和顏色轉化、植物正常衰老過程和逆境脅迫響應[23,30],因此,在煙草中系統(tǒng)研究bZIP轉錄因子對研究煙葉的生長發(fā)育成熟具有重要意義。本研究擬通過生物信息學方法在全基因組范圍內對煙草bZIP轉錄因子家族成員進行系統(tǒng)分析,結合不同成熟度煙葉的轉錄組數(shù)據,探究該家族成員在不同成熟度煙葉中的表達情況,研究結果將為相關bZIP轉錄因子的功能研究提供參考。
1 材料和方法
1.1 煙草bZIP基因家族成員的鑒定和理化性質預測
從擬南芥公共資源TAIR(http://www.arabidopsis.org/)中下載擬南芥75個bZIP基因的蛋白質序列[4],以擬南芥bZIP的蛋白質序列作為母序列,利用blast程序在茄科植物基因組數(shù)據庫(https://solgenomics.net/)搜索煙草的bZIP候選蛋白序列,將相似度≥30%,并且E值≤-10的序列確定為最初的候選蛋白。進一步利用NCBI(https://www.ncbi.nlm.nih.gov/)的CDD程序分析煙草bZIP候選蛋白序列的結構域。將包含bZIP轉錄家族特有保守結構域的序列確定為最終的候選蛋白,利用ExPASy軟件(https://www.expasy.org)對煙草bZIP候選蛋白相關的理化性質進行預測分析,將對應的基因號,基因的染色體定位信息和蛋白質信息構建列表。
1.2 煙草bZIP基因進化分析
運用Clustal X軟件[31]對最終確認的候選bZIP蛋白序列進行多序列比對,運用軟件MEGA-X[32]構建煙草bZIP蛋白家族的進化樹,使用的算法為鄰接法(Neighbor-joining model;NJ),Bootstrap值設定為1000。
1.3 煙草bZIP基因結構和蛋白結構域分析
利用Edwards等人發(fā)布的普通煙草K326的基因組數(shù)據庫(2017)(https://solgenomics.net/organism/Nicotiana_tabacum/genome),下載煙草bZIP基因結構的相關信息文件(包括GFF 文件、核酸序列文件以及蛋白序列文件),利用在線的軟件工具Gene
Structure Display Server(GSDS)(http://gsds.cbi.pku.edu.cn/)[33]分析煙草bZIP基因外顯子/內含子(intron/exon)的分布情況并繪制基因結構圖。利用軟件DNAman 6.0(Lynnon Biosoft,Quebec,Canada)對煙草bZIP家族成員的bZIP保守域氨基酸組成進行可視化分析,默認參數(shù)。進一步通過MEME(http://meme-suite.org/tools/meme)[34]在 線工具分析煙草bZIP蛋白的保守基序(motif),設置參數(shù)為:20個motif,最小和最大長度分別設定為5和200 bp。
1.4 不同成熟度煙葉bZIP基因的表達分析
以福建三明尤溪大田生長的普通煙草品種翠碧一號的中部葉(從下往上數(shù)8~10葉位)為材料,在同一田塊且種植間隔7天的兩批植株中選擇長勢相對均勻一致的煙株,由三位熟悉煙葉成熟度判定的專業(yè)人員根據葉色,葉脈和絨毛等外觀特征[35]共同衡量后,在同一時間段分別采收5個成熟度(M1、M2、M3、M4和M5)的葉樣。每3個葉片為一個樣品,3個生物學重復。采用打孔器沿著葉片主脈均勻打孔取樣,迅速置于-80℃液氮保存?zhèn)溆谩2捎冒偬┛酥参锟俁NA提取試劑盒(離心柱型)分別提取各檔成熟度煙葉樣品總RNA。采用Illumina HiSeqTM 2000平臺開展煙葉樣品轉錄組測序分析,其中,文庫構建和測序工作由百邁克生物科技有限公司完成,利用基因的FPKM值衡量基因表達水平[36]。從轉錄組數(shù)據中獲得煙草bZIP家族各成員的FPKM值(log2轉換),利用R語言gplots包的heatmap功能[37]繪制不同成熟度煙葉的bZIP基因表達熱圖。采用SYBR Premix Ex Taq(Takara)兩步法進行qRT-PCR測定,分析煙草部分bZIP基因成員在不同成熟度煙葉中的表達情況,qRT-PCR擴增引物序列和煙草Actin內參基因引物見表1。每個樣品3次技術重復和3次生物學重復。相對表達水平衡量采用2-△△Ct方法[38],以M1成熟度的基因表達量為參照,計算bZIP基因家族成員在不同成熟度煙葉中的相對表達量,根據 3次生物學重復結果計算標準差。
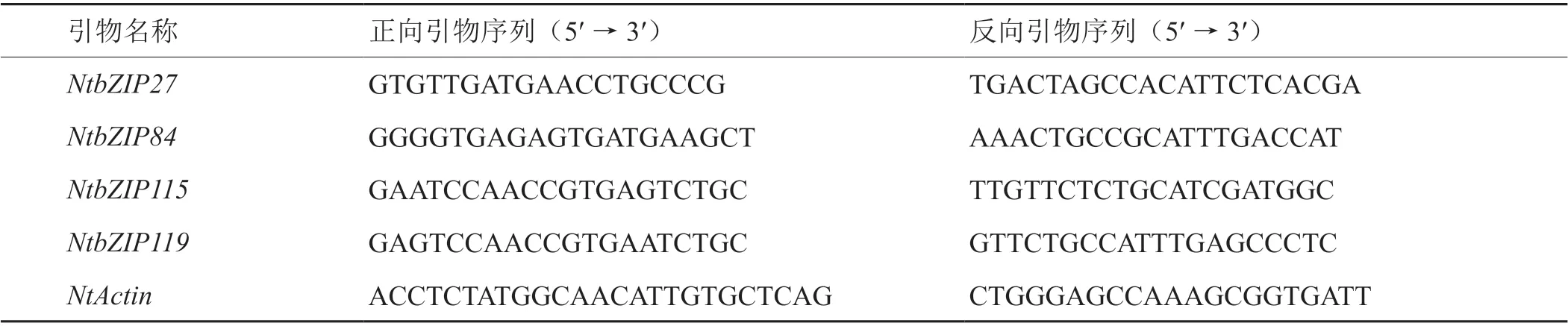
表1 煙草NtbZIP基因qRT-PCR分析特異引物Tab. 1 NtbZIP gene-specific primers of tobacco used for qRT-PCR analysis
2 結果與分析
2.1 煙草bZIP基因家族成員理化性質及其染色體定位
通過相似性搜索和結構域判定,在煙草中共鑒定出126個bZIP基因,依次重新編號命名為NtbZIP1~NtbZIP126,基因的具體信息列于表2,包括基因的ID,物理位置,外顯子數(shù)目,蛋白長度、分子量和等電點等。由表2可知:煙草NtbZIP基因的外顯子數(shù)目、蛋白長度和分子量等差異較大。其中,NtbZIPs外顯子數(shù)目最小的僅為1,最大達到13個,含有1個和4個外顯子的基因數(shù)量最多,分別達到26和33個;編碼蛋白質長度最大達812 aa(NtbZIP22),最小僅為112 aa(NtbZIP117);相對分子質量最小為12913.63 Da(NtbZIP117),最 大 的 為87319.33(NtbZIP22);理 論 等 電 點在5.03(NtbZIP123)~9.79(NtbZIP6)之間。染色體定位結果顯示,126 個NtbZIP基因中有58個NtbZIP定位于染色體中,且基因在各染色體上的分布不均勻,其中第19號和第24號染色體最多,各含7個NtbZIP基因,而第8、14、18和20號染色體上未發(fā)現(xiàn)NtbZIP基因。另外的68個NtbZIP基因定位到Scafford上,目前尚無法確定這些基因在染色體上的具體位置。

表2 煙草bZIP基因家族成員信息Tab. 2 The characters of bZIP gene family members of Nicotiana tabacum

表2(續(xù)1)

表2(續(xù)2)

表2(續(xù)3)
2.2 煙草bZIP基因的結構與進化
利用126個NtbZIPs基因的蛋白序列構建系統(tǒng)進化樹,結果發(fā)現(xiàn)(圖1a):煙草的bZIP基因家族可分為11個亞族(bootstrap≥80%),分別為A、B、C、D、E、F、G、H、I、J和S亞族,其中,由于bootstrap值太低,NtbZIP123未能歸到任何一個亞族。在11個亞族中,S是最大的亞族,包含28個成員。與此同時,D、A、I和G的成員也較多,分別含有23,20,17和11個成員,這5個亞族成員的總和為NtbZIP家族成員總數(shù)的78.6%。此外,B亞族的成員最少,僅有2個,亞族J和H也僅含有3個成員。基因結構分析表明(圖1b):NtbZIP基因序列的長度、內含子/外顯子的數(shù)量和分布差異較大。綜合結構和進化分析結果(圖1)可以發(fā)現(xiàn):在同一亞族中的大多數(shù)成員具有相似的基因結構。例如:在最大的S亞族成員中,大多數(shù)基因的長度較短,在28個成員中有23個僅含有1個外顯子,占該亞族總數(shù)的82.1%;而在D亞族中,NtbZIP家族成員的基因長度較大,外顯子數(shù)目較多(7-12個)。由此可以推測:該家族基因的結構與進化關系密切,家族成員的擴張與內含子的插入或缺失有關。
2.3 煙草bZIP蛋白保守結構
為分析煙草bZIP家族成員蛋白的結構域組成,進一步利用DNAman軟件對126個NtbZIP蛋白中的結構域進行多序列比對分析,并將其可視化(圖2)。結果發(fā)現(xiàn):煙草bZIP結構域非常保守,在其N端都含有一個堿性區(qū)域(N-X7-R/K),隨后具有保守的RK或KK以及亮氨酸拉鏈區(qū)域,該結構參與形成bZIP蛋白與 DNA 結合之前的二聚體化過程,此外,發(fā)現(xiàn)部分亮氨酸被其他疏水殘基(異亮氨酸、纈氨酸核甲硫氨酸等)取代的現(xiàn)象,該結果與前人的研究一致[4, 39]。

圖2 煙草bZIP家族蛋白保守結構域序列比對Fig.2 Alignment of conserved domain of the bZIP family proteins in Nicotiana tabacum
利用在線工具MEME分析NtbZIPs基因的蛋白序列中Motif組成的多樣性,結果表明(圖3):在126條煙草的bZIP蛋白序列中共鑒定出20個Motif(表3),同一亞族的成員大多含有相同的基序組成和排列順序。例如:D亞族成員都含有Motif1、Motif6、Motif2、Motif5、Motif15和Motif3;E亞族和I亞族,大多數(shù)成員含有Motif1和Motif4,該結果與基因結構的分析情況(圖1)一致。與此同時,不同亞族中的蛋白序列Motif組成和分布存在較大差異,有些Motif只出現(xiàn)在特定的亞族中,例如Motif2、Motif3和Motif5只出現(xiàn)在D亞族中,Motif13和Motif7只出現(xiàn)在F亞族中,Motif12和Motif17只存在于A亞族,Motif9只存在于I亞族;而一些基序是幾個亞家族共有的,例如:Motif4存在于E和I亞族,Motif18存在于A、G和I亞族等。由這些結果可以推測:NtbZIPs基因在進化過程中出現(xiàn)了功能分化。

圖1 煙草bZIP家族基因結構與進化Fig. 1 Gene structure and evolution of bZIP family in Nicotiana tabacum

表3 煙草bZIP蛋白的20個預測保守基序Tab. 3 Sequences of 20 predicted motifs of NtbZIP proteins
進一步利用CDD對20個基序進行鑒定,其中,Motif1注釋為bZIP結構域組成部分,在126條NtbZIPs蛋白中,除NtbZIP38和NtbZIP124外,其余124條蛋白序列均含有Motif1。此外,Motif2注釋為DOG1結構域組成部分,該結構域與種子的休眠有關[40-41]。Motif16被注釋為MFMR結構域,Motif10被注釋為bZIP_C結構域的組成部分。
2.4 煙草bZIP基因家族的系統(tǒng)進化分析
為了探究煙草bZIP轉錄因子的進化歷程,整合126條煙草的bZIP蛋白序列和75條擬南芥的bZIP蛋白序列構建系統(tǒng)發(fā)育樹,并對照Jakoby 等在擬南芥的分組規(guī)則分類[4],將bZIP蛋白共聚成11個分支(圖4),每個分支中都含有煙草和擬南芥的bZIP成員,而且每個亞族基因數(shù)量分布比例相似,說明這兩個物種的bZIP 轉錄因子家族成員并未因物種差異而單獨聚類。在這11個分支中,S分支是含有成員最多的亞族,共有43個的成員,其中煙草28個,擬南芥15個,而H包括的bZIP成員最少(5個),其中煙草2個,擬南芥3個。與擬南芥相比[4],本研究新增加的一個亞族J,該亞族共有6個成員,包括煙草3個成員和擬南芥3個成員。除J亞族外,擬南芥bZIP的聚類與前人研究結果基本一致,只有少數(shù)成員在聚類中分到不同的分支,說明bZIP基因家族在進化上相對保守。在整個系統(tǒng)發(fā)育樹中共發(fā)現(xiàn) 77對同源姐妹對,其中4對為直系同源蛋白,73對為旁系同源蛋白(煙草53 對,擬南芥20對),由此可見,煙草的旁系同源蛋白數(shù)目是擬南芥的兩倍多,該結果可能是由于普通煙草源于絨毛煙草和林煙草的染色體加倍進化而來,因此存在大量的相似序列。

圖 3 bZIP蛋白質保守基序組成與分布Fig. 3 Conserved motifs for bZIP proteins in Nicotiana tabacum

圖4 煙草與擬南芥bZIP基因家族系統(tǒng)進化分析Fig. 4 Phylogenetic analysis of Nicotiana tabacum and Arabidopsis thaliana bZIP genes
2.5 不同成熟度煙葉中NtbZIP基因的表達情況
為分析NtbZIPs在不同成熟度煙葉中的表達變化,本研究采集了5個成熟度(M1-M5)煙葉。從5檔成熟度煙葉的轉錄組數(shù)據中獲取NtbZIPs基因的表達值,剔除部分未表達或低表達的基因(PFKM<0.5),進一步繪制NtbZIPs基因(77個)的表達熱圖。由圖5可知:不同NtbZIP基因的表達量差異較大,其中,分支Ⅲ中的27個NtbZIP基因在5個成熟期表達量較低,而分支Ⅱ的10個NtbZIP基因表達量較高,包 括NtbZIP82,NtbZIP27,NtbZIP111,NtbZIP76,
NtbZIP32,NtbZIP120,NtbZIP115,NtbZIP119,NtbZIP123和NtbZIP31等基因。值得注意的是:NtbZIP84,NtbZIP120,NtbZIP119和NtbZIP115等基因的表達量隨著成熟度的提高呈現(xiàn)規(guī)律性下降趨勢。這些結果暗示了煙草NtbZIP家族成員間在功能上可能具有多樣性。
進一步挑選表達變化比較明顯的NtbZIP27,NtbZIP84,NtbZIP115和NtbZIP119等4個 基 因 開展RT-PCR驗證,由結果可知(圖6):NtbZIP27基因在M1-M4等4個成熟期的表達變化不大,在M5成熟期表達量下降近5倍。與M1成熟期相比,NtbZIP84,NtbZIP115和NtbZIP119等基因的表達隨著成熟度的提高呈現(xiàn)明顯下降趨勢,這3個基因在M1和M5成熟期的表達量差異近10倍。此外,隨著煙葉成熟度提高,這4個NtbZIP基因的表達在qRTPCR分析(圖6)和轉錄組測序分析(圖5)中表現(xiàn)出類似的變化趨勢,說明轉錄組測序結果較可靠。

圖5 NtbZIPs基因在不同成熟度煙葉中的表達情況Fig. 5 The expression of NtbZIPs in tobacco leaves at different stage of maturity

圖6 NtbZIPs基因在不同成熟度煙葉中的表達情況Fig. 6 Relative expression level of 4 NtbZIPs in tobacco leaves at different stage of maturity
3 討論
在全基因組測序完成的前提下,全基因組范圍內的基因鑒定為基因功能研究提供了一條便捷有效的途徑。bZIP轉錄因子是真核生物中普遍存在的一個大基因家族,許多物種在全基因組水平上開展了bZIP基因家族的鑒定。本研究對煙草中bZIP基因家族的成員進行了全基因組水平上鑒定和分析,最終獲得了126個NtbZIP基因。由于煙草基因組主要通過基因富集區(qū)測序獲得,且其全基因組測序尚未完成,可能存在未被鑒定的NtbZIP基因。因此,煙草中實際存在的bZIP將超過本研究鑒定的126個。顯然,煙草中bZIP基因數(shù)量高于擬南芥(75個)[4],水稻(89個)[3],甘薯(41個)[8]和番茄(69個)[9]等物種中的bZIP基因數(shù)量,該結果主要源于普通煙草是異源四倍體,其存在兩套異源基因組,分別來自于S基因組供體林煙草和T 基因組供體絨毛煙草兩個祖先種,大約在 20 萬年前經過一次種間雜交產生[42]。因此,在本研究鑒定的126個bZIP基因中存在大量的旁系同源基因(53對)。研究表明,基因倍增后,最初的結果是出現(xiàn)兩個完全相同的基因,在選擇壓作用下,其中一個基因保持原有的功能,而另一個基因更容易產生突變積累,從而變成假基因或產生新的功能[43]。因此,多倍化可能是產生新基因的主要途徑,這可能是導致煙草中bZIP基因擴張的重要原因。
基因的表達模式是基因功能的重要體現(xiàn)。已有的研究表明,bZIPs家族廣泛參與植物的生長發(fā)育、顏色轉化和植株的正常衰老過程[30]。本研究中不同成熟期煙葉NtbZIPs基因的表達分析發(fā)現(xiàn):NtbZIPs基因在不同成熟度中廣泛表達,而且大多NtbZIPs基因在煙葉的不同成熟期表現(xiàn)出差異性表達,其中NtbZIP84,NtbZIP115,NtbZIP119和NtbZIP120等基因表達量較高,且隨成熟度提高表現(xiàn)出規(guī)律性變化下降趨勢,暗示這些基因隨著煙葉成熟度的提高,物質轉化和降解的完成,表達量逐漸下降。與此同時,部分基因隨著成熟度提高表達量呈現(xiàn)上升趨勢(NtbZIP123),前人研究中也發(fā)現(xiàn)類似現(xiàn)象,例如Yang等[22]研究發(fā)現(xiàn):煙草bZIP家族成員tbzF和tbz17基因的表達量與煙葉中的葉綠素含量呈負相關。此外,在不同成熟度NtbZIP基因表達的分析發(fā)現(xiàn)(圖5):分支Ⅱ的10個NtbZIP基因表達量很高,值得注意的是:這10個基因中,除了NtbZIP82,NtbZIP27和NtbZIP123外,其余7個均分布在S亞族中(圖1),暗示該亞族中的這些基因可能在不同成熟度中煙葉中發(fā)揮重要作用,并且可能具有類似的功能,但具體生物學功能還需要進一步的驗證。
通常認為,聚類中關系越密切則其相似性就越大,因此,聚在同一亞族中的基因在結構與功能上往往有一定的相似性。已有研究表明:擬南芥中的AT2G35530基因主要通過調控光、GA及 ABA等基因的應答[44];擬南芥中的 AT5G06950與AT5G06960為bZIP轉錄因子家族的 TGA 亞族,參與擬南芥的抗逆調控[45-46]。因此,我們推測在系統(tǒng)樹G亞族中(圖4)與擬南芥AT2G35530聚類距離較近的NtbZIP96,NtbZIP97,NtbZIP98和NtbZIP99等基因可能參與調控光、GA與ABA激素的代謝;D亞族中與擬南芥AT5G06950和AT5G06960聚類距離較近的NtbZIP45,NtbZIP46,NtbZIP49和NtbZIP50等基因可能在逆境響應中發(fā)揮作用,具體有待進一步的實驗驗證。本文所提供的NtbZIPs基因的表達和進化信息,可為NtbZIPs基因的進一步功能研究奠定基礎。
4 結論
本研究在煙草基因組中共鑒定了126個NtbZIP基因。NtbZIP成員基因結構多樣,蛋白質長度差異較大,共分為11個亞族。進化分析顯示NtbZIP基因分化的時間早于擬南芥與煙草分化的時間,異源四倍體的普通煙草中存在大量的NtbZIP旁系同源基因對(53對)。本研究中NtbZIPs在不同成熟度煙葉中廣泛表達,其中NtbZIP84,NtbZIP115,NtbZIP119和NtbZIP120等基因表達量隨成熟度提高而逐漸降低。系統(tǒng)分析基因家族各成員的序列特征、進化關系和表達模式等可為基因的功能研究提供指導。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛(wèi)生(2019年5期)2019-05-21 01:08:34
浙江中西醫(yī)結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
