棉花核酸外切酶基因GhWRN的克隆及功能驗(yàn)證
2021-06-19 08:39:20薛羽君魏恒玲王寒濤馬亮程帥帥郝蓬勃顧麗姣付小康蘆建華喻樹迅
棉花學(xué)報(bào) 2021年3期
關(guān)鍵詞:植物
薛羽君,魏恒玲,王寒濤,馬亮,程帥帥,郝蓬勃,顧麗姣,付小康,蘆建華,喻樹迅
(中國農(nóng)業(yè)科學(xué)院棉花研究所/棉花生物學(xué)國家重點(diǎn)試驗(yàn)室,河南安陽455000)
作為1種水解磷酸鍵的酶,核酸酶可以剪切核酸中的3′或者5′磷酸二酯鍵,它對(duì)DNA和RNA的剪切催化功能是各種生命體中必不可少的。例如,5′-3′核酸外切酶和核酸內(nèi)切酶參與DNA復(fù)制過程,同時(shí)也是移除RNA引物的必需組分;3′-5′核酸外切酶是DNA校正修復(fù)過程的必需組分,因此核酸酶的活性是DNA完整復(fù)制的保證[1-4]。核酸酶突變的植物,在種子發(fā)育、管狀細(xì)胞及維管組織發(fā)育中都顯現(xiàn)出不同程度的生長發(fā)育障礙。核酸酶功能缺失的高等植物會(huì)因核酸降解失衡從而出現(xiàn)各種畸形發(fā)育,也會(huì)影響植物幼苗期的光形態(tài)建成,從而造成植株的生長停滯與死亡。如百日草(Zinnia elegans)的核酸內(nèi)切酶ZEN1在不影響液泡形態(tài)的前提下調(diào)節(jié)細(xì)胞核中DNA的降解,從而特異地調(diào)控管狀細(xì)胞的衰 老 和 死 亡[5]。杜 仲(Eucomm ia ulmoides)中 的EuCaN1和EuCaN2基因編碼Ca2+依賴的核酸內(nèi)切酶,定位于細(xì)胞核,具有降解DNA的功能。這2個(gè)基因經(jīng)RNA干擾后,其轉(zhuǎn)基因植株的次級(jí)木質(zhì)部出現(xiàn)分化異常的情況[6]。對(duì)擬南芥(Arabidopsis thaliana)核酸內(nèi)外切酶研究發(fā)現(xiàn),核酸內(nèi)外切酶突變體出現(xiàn)植株發(fā)育畸形的表型。在菜豆(Phaseolus coccineus)相關(guān)研究中發(fā)現(xiàn),DNA降解對(duì)各器官的平衡和發(fā)育起到了重要的作用[7-8]。PARN(Poly A-specific 3′-5′ribonuclease,多聚腺苷酸特異性核酸酶)能高效降低真核生物mRNA的聚腺苷酸尾,缺乏PARN的擬南芥胚發(fā)育明顯受阻,并且在光形態(tài)建成時(shí)期子葉會(huì)彎曲并停止生長,表明RNA的降解影響植物的生長發(fā)育過程[9]。擬南芥植物體中存在1種小干擾RNA(Small interfering RNA,siRNA)合成及微小RNA(M icroRNA,m iRNA)降解相關(guān)的RNA結(jié)合蛋白,該蛋白的缺失突變體出現(xiàn)m iRNA積累和植株生長發(fā)育缺陷[10]。
核酸酶不僅參與植物生長發(fā)育,也參與非生物脅迫應(yīng)答調(diào)控[11]。相關(guān)試驗(yàn)證明,大麥(Hordeum vulgareL.)中的Bnuc1核酸酶在鹽脅迫或者外源脫落酸(Abscisic acid,ABA)處理下誘導(dǎo)表達(dá),但在種子萌發(fā)階段外源施加赤霉素(Gibberellin,GA)時(shí)該酶無響應(yīng)[12]。擬南芥中RNase通過調(diào)控mRNA的降解來影響HSP70蛋白的活性和豐度,從而影響植株對(duì)高溫脅迫的響應(yīng)[13]。試驗(yàn)表明擬南芥RNase突變體中miRNA積累,對(duì)鹽脅迫更敏感[14]。以上研究表明,擬南芥、大麥等植物中一些核酸酶基因參與植物的生長發(fā)育及非生物脅迫應(yīng)答。
基于不同熟性陸地棉品種轉(zhuǎn)錄組數(shù)據(jù)[15],通過生物信息學(xué)分析所得差異表達(dá)基因,篩選出與棉花早熟相關(guān)的基因,從中發(fā)現(xiàn)含有WRN-exo結(jié)構(gòu)域的DnaQ核酸外切酶。通過查閱文獻(xiàn),發(fā)現(xiàn)在人和動(dòng)物及多種植物中均報(bào)道過該核酸酶,而棉花中的WRN-exo未見報(bào)道。因此,本研究通過克隆得到1個(gè)棉花核酸外切酶基因GhWRN,分析該基因在不同棉花品種花芽分化中的特點(diǎn)及組織特異性的表達(dá)特征,克隆獲得該基因的啟動(dòng)子序列并進(jìn)行順式作用元件分析,在第四片真葉展平期對(duì)幼芽噴施外源激素,通過qRT-PCR(Quantitative real-time polymerase chain reaction,實(shí)時(shí)熒光定量-聚合酶鏈?zhǔn)椒磻?yīng))分析GhWRN基因響應(yīng)植物激素時(shí)的表達(dá)特征,構(gòu)建植物的過表達(dá)載體得到轉(zhuǎn)基因擬南芥并觀察其表型,從而為深入研究核酸外切酶參與棉花生長發(fā)育及調(diào)控植物在激素處理后的應(yīng)答模式奠定基礎(chǔ)。
1 材料與方法
1.1 試驗(yàn)材料
本試驗(yàn)以陸地棉早熟品種鹽早2號(hào)和晚熟品種國欣11為試驗(yàn)材料,種植于中國農(nóng)業(yè)科學(xué)院棉花研究所東場試驗(yàn)田(河南省安陽縣),大田常規(guī)管理。
取鹽早2號(hào)和國欣11幼苗子葉展平期、第一至第五片真葉展平期共6個(gè)時(shí)期頂芽樣品。取鹽早2號(hào)盛花期的根、莖、葉、苞片、萼片、花瓣、雄蕊、雌蕊,所有材料于-80℃保存?zhèn)溆谩1驹囼?yàn)所用野生型擬南芥為哥倫比亞型(Col-0生態(tài)型),由中國農(nóng)業(yè)科學(xué)院棉花研究所棉花早熟育種課題保存,培養(yǎng)在溫度為22℃、光周期為16 h光照/8 h黑暗的溫室。
1.2 試驗(yàn)方法
1.2.1GhWRN基因的克隆。采用北京天根生化科技公司植物總RNA提取試劑盒提取鹽早2號(hào)第五片真葉展平期頂芽中的RNA,利用超微量分光光度儀檢測總RNA的濃度和純度。利用PrimeScript RT reagent kit with gDNA Eraser(Perfect Real Time)試劑盒(寶生物,中國大連)將RNA反轉(zhuǎn)錄合成第1鏈cDNA,以該cDNA為模板,利用Oligo 7軟件設(shè)計(jì)特異引物GhWRN-F/R(表1),擴(kuò)增GhWRN基因的全長cDNA序列。將目的條帶切膠回收后,與pGEM-T Easy載體連接,轉(zhuǎn)化至大腸桿菌DH5α感受態(tài)細(xì)胞(諾唯贊,中國南京),挑取單克隆,經(jīng)菌液PCR鑒定正確后,送由尚亞技術(shù)生物(鄭州)有限公司測序。
1.2.2GhWRN基因的生物信息學(xué)分析。將GhWRN基因的DNA序列提交到Gene Structure Display Server online software(http://gsds.cbi.pku.edu.cn./)網(wǎng)站預(yù)測基因結(jié)構(gòu)。使用DNAMAN軟件進(jìn)行序列比對(duì),利用MEGA7軟件的鄰接法(Neighbor-Joining,NJ)構(gòu)建進(jìn)化樹。使用ExPASy在線工具(http://www.expasy.org/tools/)分析GhWRN的蛋白質(zhì)分子量和等電點(diǎn)。用CBSTMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)和CBS-SignalP(http://www.cbs.dtu.dk/services/SignalP/)分析蛋白質(zhì)的跨膜結(jié)構(gòu)和信號(hào)肽。利用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.htm l)和ExPASy-SW ISS-MODEL(https://www.sw issmodel.expasy.org/)預(yù)測蛋白質(zhì)的二級(jí)和三級(jí)結(jié)構(gòu)。在Cotton-FGD上獲得GhWRN(Gh_D12G1228)基因轉(zhuǎn)錄起始位點(diǎn)上游1 500 bp的核苷酸序列,使用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/htm l/)進(jìn)行在線分析和功能預(yù)測。
1.2.3激素處理棉花幼苗。根據(jù)GhWRN基因啟動(dòng)子的分析結(jié)果,對(duì)正常生長的第四片真葉展平期的鹽早2號(hào),分別用以下激素葉面噴施:100μmol·L-1的生長素(Auxin,IAA)、200μmol·L-1的脫落酸(Abscisic acid,ABA)和5 mmol·L-1的水楊酸(Salicylic acid,SA),同時(shí)將600 m L蒸餾水加200μL無水乙醇(各激素溶解時(shí)使用200μL無水乙醇溶解)作為對(duì)照,取處理前(0 h)及處理后0.5 h、1.5 h、3 h、6 h、9 h、12 h、24 h的頂芽,各處理重復(fù)3次,各重復(fù)取3個(gè)樣品,置于液氮中速凍,保存于-80℃?zhèn)溆谩?/p>
1.2.4轉(zhuǎn)基因擬南芥的遺傳轉(zhuǎn)化及表型鑒定。采用農(nóng)桿菌介導(dǎo)的花序浸染法[16],將構(gòu)建的含35S∷GhWRN的pBI121表達(dá)載體轉(zhuǎn)化野生型擬南芥,使用含有卡那霉素的1/2MS培養(yǎng)基篩選轉(zhuǎn)基因苗,將每一代的陽性植株移栽種植并檢測,至T3開始表型觀察、鑒定工作。取生長4周的T3陽性擬南芥和野生型擬南芥的葉片,采用CTAB法[17]提取DNA,以野生型擬南芥(WT)作為陰性對(duì)照。利用35S啟動(dòng)子序列的上游引物At35S-F(表1)及GhWRN基因下游引物GhWRN-R進(jìn)行轉(zhuǎn)基因檢測,使用1%瓊脂糖凝膠電泳檢測。根據(jù)T3植株表型觀察,提取生長28 d的已開花的T3轉(zhuǎn)基因擬南芥葉片RNA,選取擬南芥花發(fā)育相關(guān)的關(guān)鍵基因AtFT、AtSOC1、AtAP1、AtFUL進(jìn)行基因表達(dá)量分析,熒光定量引物見表1。
1.2.5GhWRN基因的qRT-PCR分析。使用Oligo 7設(shè)計(jì)GhWRN的熒光定量引物qRT-PCRF/R (表1),以GhHistone3(Accession No.:AF024716)作為棉花的內(nèi)參基因,Atactin2為擬南芥內(nèi)參基因,分析GhWRN基因在不同陸地棉品種、鹽早2號(hào)不同處理下的表達(dá)模式,每個(gè)樣本做3次生物學(xué)重復(fù)。qRT-PCR反應(yīng)體系和反應(yīng)程序按照SYBR Prem ix ExTaq(DRR041A)熒光定量試劑盒說明書進(jìn)行操作,所用儀器為ABI 7500Real Time PCRSystem(ABI,美國)。每個(gè)樣品重復(fù)3次,目的基因的相對(duì)表達(dá)量采用2-ΔΔCT方法[18]計(jì)算,所得數(shù)據(jù)利用GraphPad Prism5軟件繪圖。

表1 本研究中使用的引物Table 1 Primers used in this study
2 結(jié)果與分析
2.1 GhWRN基因克隆及特征分析
從轉(zhuǎn)錄組數(shù)據(jù)[15]中獲得1個(gè)在不同熟性陸地棉品種中表達(dá)存在明顯差異的Coding sequence(CDS),在CottonFGD網(wǎng)站上進(jìn)行Blast序列比對(duì),根據(jù)南京農(nóng)業(yè)大學(xué)測序基因組[19]獲得該基因(Gh_D12G1228)的開放閱讀框(Open reading frame,ORF)序列,發(fā)現(xiàn)該基因是含有WRN-exo結(jié)構(gòu)域的DnaQ核酸外切酶,所以將該基因命名為GhWRN。以cDNA模板進(jìn)行PCR擴(kuò)增,經(jīng)檢測條帶正確后,送測序分析。該GhWRN基因ORF為798 bp,無內(nèi)含子。
通過National Center for Biotechnology Information(NCBI,https://www.ncbi.nlm.nih.gov/)的非冗余蛋白數(shù)據(jù)庫搜索GhWRN蛋白同源序列,發(fā)現(xiàn)可可(Theobroma cacao)XP_007051231.2、哥倫比亞錦葵(Herrania umbratical)XP_021279751.1、辣椒(Capsicum annuum)KAF3629001.1、煙草(Nicotiana attenuate)OIT08451.1與GhWRN蛋白相似性分別為68%、68%、51%、48%,擬南芥(A.thaliana)At5g06450蛋白為含DnaQ-like的核酸外切酶[20]。對(duì)以上6個(gè)蛋白進(jìn)行多序列比對(duì),使用MEGA7軟件構(gòu)建系統(tǒng)發(fā)育樹,表明GhWRN與可可、哥倫比亞錦葵的同源蛋白的親緣關(guān)系最近(圖1)。
GhWRN蛋白包含265個(gè)氨基酸,預(yù)測相對(duì)分子量是30.23 kDa,等電點(diǎn)是9.58。序列進(jìn)行分析,發(fā)現(xiàn)GhWRN蛋白中富含賴氨酸(13.6%)、亮氨酸(9.8%)和丙氨酸(7.9%),含少量色氨酸(0.8%),不含吡咯賴氨酸、硒半胱氨酸;蛋白整體呈堿性且?guī)д姾桑渲袔ж?fù)電荷的殘基31個(gè),帶正電荷的殘基49個(gè);預(yù)測半衰期約為30 h;蛋白不穩(wěn)定指數(shù)為28.58,指數(shù)較低為穩(wěn)定蛋白。蛋白的親疏水性分析發(fā)現(xiàn),親水性氨基酸明顯多于疏水性氨基酸,親水性平均系數(shù)為-0.451,因此該蛋白屬親水性蛋白。
CBS-TMHMM在線預(yù)測表明,GhWRN蛋白含跨膜螺旋的氨基酸期望值為17.20876,推測不存在跨膜螺旋結(jié)構(gòu);且CBS-SignalP在線預(yù)測其不存在信號(hào)肽,綜合推測GhWRN是非分泌蛋白。由SOPMA軟件對(duì)該蛋白的氨基酸序列二級(jí)結(jié)構(gòu)進(jìn)行預(yù)測。結(jié)果表明,該蛋白主要由α-螺旋構(gòu)成。用ExPASy-SWISS-MODEL對(duì)蛋白進(jìn)行三維結(jié)構(gòu)預(yù)測,結(jié)果顯示該蛋白的三級(jí)結(jié)構(gòu)多為α-螺旋和無規(guī)則卷曲。
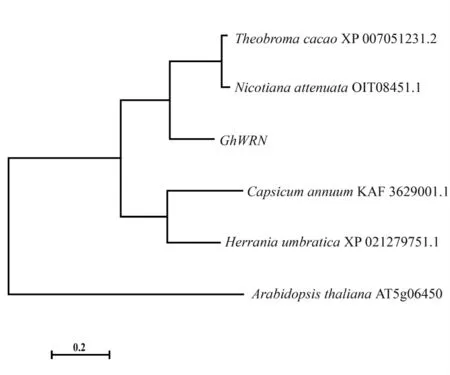
圖1 GhWRN進(jìn)化樹分析Fig.1 Phylogenetic tree analysis of GhWRN proteins
2.2 GhWRN基因的啟動(dòng)子生物信息學(xué)分析
對(duì)GhWRN基因上游1 500 bp啟動(dòng)子序列進(jìn)行分析,結(jié)果表明:該啟動(dòng)子包含多個(gè)典型的核心元件,如TATA框和CAAT框。此外,該啟動(dòng)子還包含豐富的順式調(diào)控元件:環(huán)境脅迫響應(yīng)元件,如光響應(yīng)元件(Box4、G-Box、GT1-motif、MRE、TCCC-motif等)、逆境脅迫響應(yīng)元件(TC-rich repeats)等;激素響應(yīng)元件,如脫落酸響應(yīng)元件(ABRE)、生長素響應(yīng)元件(AuxRR-core、TGA-element)等(表2)。表明GhWRN基因的表達(dá)可能與非生物脅迫及植物激素誘導(dǎo)相關(guān)。
2.3 GhWRN基因的表達(dá)分析
通過qRT-PCR分析GhWRN基因在不同熟性陸地棉真葉期(True leaf stage,TLS)頂芽及各組織的表達(dá)特征。結(jié)果顯示,在早熟材料鹽早2號(hào)的2片真葉期(兩葉期)至五葉期該基因表達(dá)量顯著增加;在晚熟材料國欣11各時(shí)期表達(dá)量無顯著變化(圖2A),與其在鹽早2號(hào)兩葉期表達(dá)量相當(dāng)。提取鹽早2號(hào)四葉期的根、莖、葉、花瓣、雄蕊、雌蕊、苞片、副萼,進(jìn)行組織特異性表達(dá)分析。結(jié)果表明,GhWRN基因在不同組織中均有表達(dá),在苞片、雄蕊、葉中表達(dá)量較高,在根中表達(dá)量最低(圖2B)。

表2 GhWRN基因啟動(dòng)子調(diào)控元件預(yù)測Table 2 Prediction of cis-elements of GhWRN promoter
2.4 GhWRN基因響應(yīng)植物激素的表達(dá)分析

圖2 GhWRN基因的特異性表達(dá)分析Fig.2 Expression specificity analysis of GhWRN gene
通過qRT-PCR分析在不同植物激素處理下棉花頂芽中GhWRN基因表達(dá)量的變化。結(jié)果顯示(圖4):ABA處理后0.5 h,該基因表達(dá)量較對(duì)照極顯著升高;處理后12 h、24 h該基因表達(dá)量較對(duì)照顯著下降(圖3A)。IAA處理后除1.5 h、6 h外,其他檢測時(shí)期該基因表達(dá)量較對(duì)照均顯著升高(圖3B)。SA處理后9 h、12 h該基因表達(dá)量存在顯著差異(圖3C)。不同激素誘導(dǎo)后不同時(shí)間GhWRN基因表達(dá)水平不同,脫落酸、生長素極顯著誘導(dǎo)GhWRN基因上調(diào)表達(dá),表明GhWRN基因的表達(dá)受外源激素的調(diào)控,推測這些調(diào)控與啟動(dòng)子中的激素響應(yīng)元件相關(guān)。
2.5 轉(zhuǎn)基因擬南芥的鑒定
觀察野生型擬南芥與轉(zhuǎn)基因T3擬南芥的表型,發(fā)現(xiàn)生長至28 d時(shí),轉(zhuǎn)基因擬南芥T3陸續(xù)開花而野生型擬南芥剛剛完成蓮座葉生長(圖4A)。選取野生型擬南芥及3個(gè)轉(zhuǎn)基因擬南芥株系,每個(gè)株系各挑選4株植株混取葉片,對(duì)Gh-WRN基因表達(dá)進(jìn)行定量檢測。試驗(yàn)結(jié)果顯示,3個(gè)陽性株系均能檢測到目的條帶;轉(zhuǎn)基因擬南芥株系GhWRN基因在3個(gè)陽性株系中的表達(dá)量都很高且顯著高于野生型(圖4B、4C),說明已獲得穩(wěn)定的轉(zhuǎn)基因擬南芥陽性株。為了確定開花時(shí)間,對(duì)野生型擬南芥和T3轉(zhuǎn)基因擬南芥開花時(shí)的蓮座葉數(shù)和開花天數(shù)進(jìn)行了數(shù)據(jù)統(tǒng)計(jì),發(fā)現(xiàn)轉(zhuǎn)基因擬南芥各株系的開花時(shí)間比野生型的擬南芥提前約5.5 d。其中,野生型擬南芥從抽薹(莖長1 cm)至首次開花為32.8 d,蓮座葉平均數(shù)為11.9;T3轉(zhuǎn)基因擬南芥株系從抽薹至首次開花平均為27.3 d,蓮座葉平均數(shù)為9.3(表3)。綜合T3陽性株基因表達(dá)量、開花時(shí)間及提前開花的表型判斷,GhWRN基因與植株開花時(shí)間有關(guān)。
2.6 GhWRN影響花發(fā)育信號(hào)通路相關(guān)基因的表達(dá)
為了研究過表達(dá)GhWRN在擬南芥中促進(jìn)開花的分子機(jī)制,選取花發(fā)育相關(guān)的關(guān)鍵基因進(jìn)行熒光定量分析。結(jié)果發(fā)現(xiàn),與花發(fā)育相關(guān)的基因AtFT、AtSOC1和AtFUL的表達(dá)量在T3轉(zhuǎn)基因擬南芥中表達(dá)量極顯著高于野生型,AtAP1表達(dá)量與野生型無顯著差異(圖5)。推測GhWRN在AtSOC1、AtFT、AtFUL基因組成的相關(guān)開花調(diào)控通路中發(fā)揮功能。

圖3 GhWRN基因響應(yīng)植物激素的表達(dá)分析Fig.3 Expression analysis of GhWRN gene in response to plant hormones
3 討論
GhWRN蛋白富含賴氨酸、亮氨酸和丙氨酸,色氨酸含量較少,核酸酶由高度親水性氨基酸賴氨酸等組成,這種親水性可能使該酶更易于同底物(水)發(fā)生反應(yīng)[11]。二級(jí)結(jié)構(gòu)預(yù)測顯示GhWRN蛋白主要由α-螺旋組成,前人研究報(bào)道指出DnaQ-like核酸外切酶有1個(gè)絕對(duì)保守的DEDD功能域,該功能域中的保守酸性氨基酸殘基多數(shù)位于α-螺旋中,GhWRN蛋白具有相同的結(jié)構(gòu),推測行使相同的功能[22]。聚類分析表明,GhWRN蛋白與可可、哥倫比亞錦葵的WRN蛋白的同源性較高。
在植物體中大部分DNA酶和RNA酶受生物脅迫影響,并且部分核酸酶的功能缺失與植物逆境抗性相關(guān)[11]。黃瓜為單性花,CsCAN核酸酶基因在其雌花發(fā)育過程特異表達(dá)。在施加外源乙烯前體物質(zhì)后,收集處理后4、8、12、16、20和24 h幼苗子葉,結(jié)果顯示隨著時(shí)間的延長CsCAN核酸酶基因上調(diào)表達(dá)從2倍增加到4倍,表明乙烯和CsCAN活性之間的關(guān)系并不簡單[23]。水稻和擬南芥中的BBD(Bifunctional nuclease in basal defense response,堿基防御反應(yīng)中的雙功能核酸酶)蛋白是1種DNA酶和RNA酶雙功能核酸酶,水稻OmBBD基因在100μmol·L-1外源ABA處理后1、2、3、4 h表達(dá)量明顯增加,表明該基因受ABA的誘導(dǎo)[24]。本試驗(yàn)中外源IAA或ABA處理后,GhWRN基因表達(dá)量有明顯升高的趨勢,推測該基因與調(diào)控植物應(yīng)對(duì)外源激素等環(huán)境變化相關(guān)。

圖4 轉(zhuǎn)基因擬南芥的表型及基因表達(dá)情況Fig.4 The phenotypes of transgenic A.thaliana p lants and GhWRN gene expression pattern
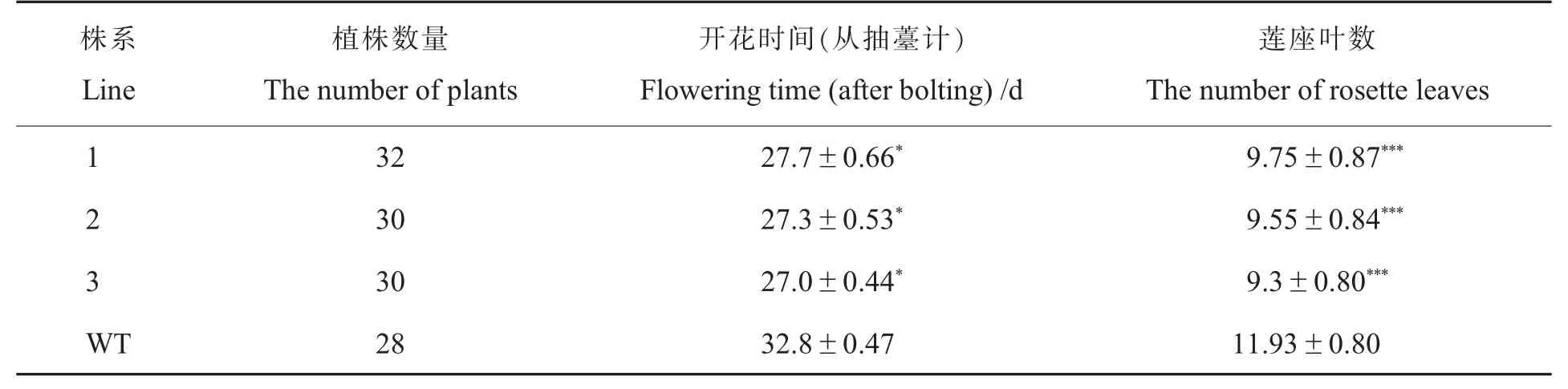
表3 抽薹時(shí)蓮座葉片數(shù)Table 3 Rosette leaf number at bolting
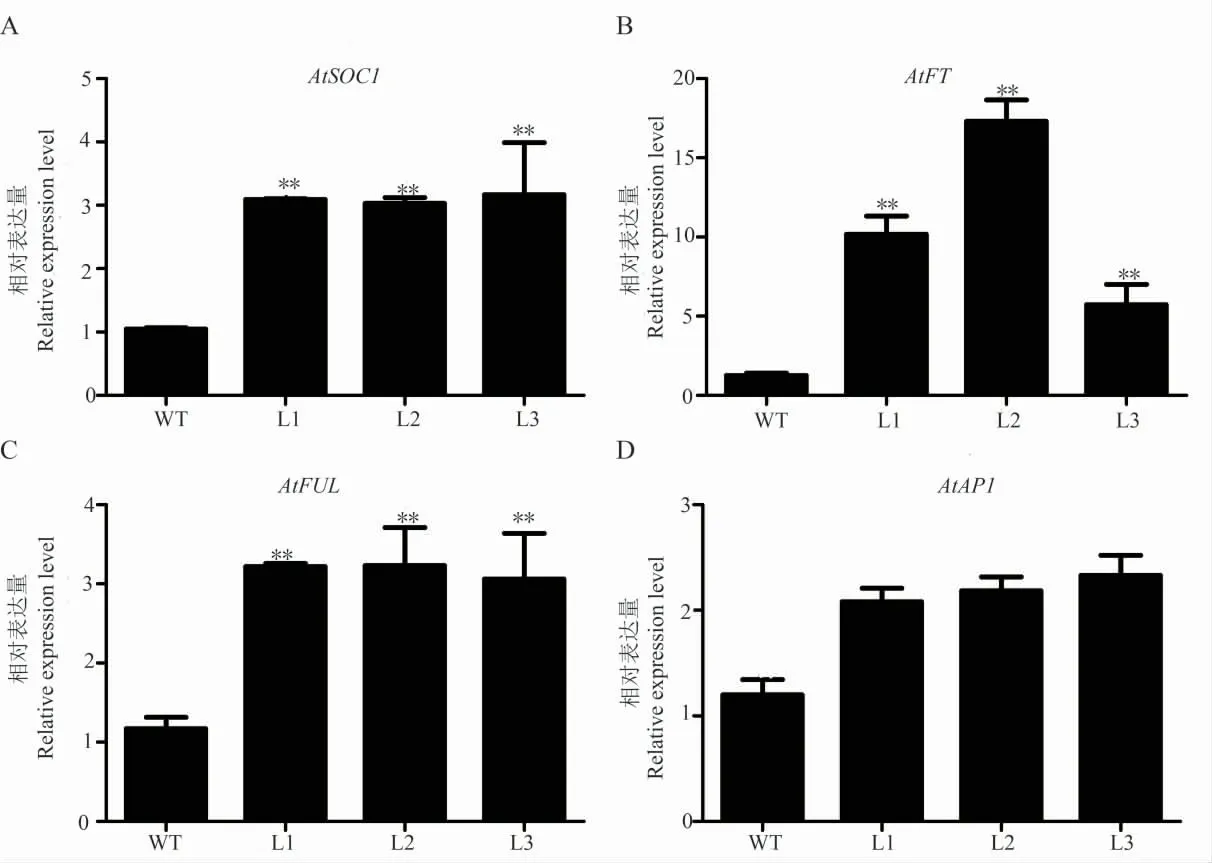
圖5 轉(zhuǎn)基因擬南芥內(nèi)源開花基因表達(dá)分析Fig.5 Expression analysis of endogenous flowering-related genes of transgenic A.thaliana
花芽分化是植物由營養(yǎng)生長向生殖生長轉(zhuǎn)化開始的標(biāo)志,石蠟切片證明早熟品種是在二葉期開始花芽的分化[21]。我們對(duì)不同熟性棉花品種的子葉、不同真葉期的葉片進(jìn)行GhWRN基因表達(dá)檢測,GhWRN基因在早熟棉二葉期的表達(dá)量升高且在花器官中表達(dá)量較高,推測該基因可能與棉花熟性及花芽分化有關(guān)。擬南芥是一種十字花科植物,廣泛用于植物遺傳學(xué)、發(fā)育生物學(xué)和分子生物學(xué)的研究,已成為一種典型的模式植物,對(duì)于植物開花時(shí)間相關(guān)的基因調(diào)控有較深入研究。已有研究表明FT作為1種成花素,在光周期途徑調(diào)控下轉(zhuǎn)移到頂芽中,與轉(zhuǎn)錄因子FD一起激活開花途徑的整合因子SOC1,該整合因子以及其他的調(diào)控因子最后直接或間接地激活A(yù)P1,促使花器官形成[21]。但是,在煙草、擬南芥等植物中發(fā)現(xiàn),F(xiàn)UL在控制花序形態(tài)和花分生組織方面與AP1功能重疊,F(xiàn)UL同樣具有促進(jìn)開花的功能[24-27]。棉花研究中,發(fā)現(xiàn)開花相關(guān)基因Gh-FLP1超表達(dá)擬南芥會(huì)促進(jìn)其內(nèi)源開花基因SOC1、FT等不同程度的上調(diào)[28]。從熒光定量結(jié)果來看,轉(zhuǎn)GhWRN基因擬南芥中AtFT、AtSOC1和AtFUL基因表達(dá)量極顯著升高,而AtAP1表達(dá)量無顯著變化。總之,GhWRN在早熟棉花品種花芽分化時(shí)期及花器官中表達(dá)量較高,且過表達(dá)GhWRN的擬南芥植株開花提前,因此推測GhWRN基因在植物中過量表達(dá)能促使AtFT升高進(jìn)而激活整合因子AtSOC1,最后激活促開花因子AtFUL而非AtAP1,從而促使轉(zhuǎn)基因擬南芥植株早開花。但對(duì)于該基因如何具體參與調(diào)控棉花開花機(jī)制仍需要進(jìn)一步深入的研究。
4 結(jié)論
本研究從陸地棉鹽早2號(hào)中克隆出GhWRN基因,開放閱讀框長度為798 bp,編碼265個(gè)氨基酸。GhWRN基因無跨膜結(jié)構(gòu),為非分泌蛋白,包含高度保守的WRN-exo功能域。GhWRN基因在早熟棉花品種花芽分化時(shí)期及雄蕊、苞片等花器官中表達(dá)量較高。啟動(dòng)子中包含多個(gè)脅迫響應(yīng)元件和激素響應(yīng)元件,外源ABA及IAA誘導(dǎo)GhWRN基因顯著上調(diào)表達(dá)。在擬南芥中過表達(dá)GhWRN基因發(fā)現(xiàn)GhWRN在植物開花時(shí)間方面發(fā)揮重要作用,推測該核酸外切酶基因通過參與花發(fā)育過程調(diào)控植物開花。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
