海島棉新海35花藥發(fā)育及花粉受精的研究
2021-06-19 08:39:34楊靜馬益贊王為然閔玲寧新民孔杰
棉花學報 2021年3期
楊靜,馬益贊,王為然,閔玲,寧新民,孔杰*
(1.華中農(nóng)業(yè)大學作物遺傳改良國家重點實驗室,武漢430070;2.新疆農(nóng)業(yè)科學院經(jīng)濟作物研究所,烏魯木齊830091)
作為雌雄同株的植物,生長環(huán)境、成花數(shù)量、花芽發(fā)育等都會影響棉花纖維品質(zhì)。花藥的正常發(fā)育作為花粉粒成熟及受精的必備條件,同樣影響棉纖維產(chǎn)量和品質(zhì)。對花藥發(fā)育的研究需從花器官發(fā)育過程的細胞結構[1]及其具體的細胞特征進行探究。自1999年,Sanderszai等[2]以擬南芥為研究對象,對花藥發(fā)育過程進行了詳細的時期劃分后,人們研究其他開花植物花發(fā)育過程時,均以擬南芥花藥內(nèi)部細胞組成特征作為參照來確定花藥發(fā)育時期。雖然花藥發(fā)育不同時期,環(huán)境條件的改變能夠決定花藥發(fā)育早期細胞的命運,但花藥細胞的特殊結構對花器官的發(fā)育及部分功能的行使具有識別作用[1,3-4]。不同的開花時間、各個花器官對環(huán)境條件的敏感程度[5]、植物本身的部分氧化物及相關基因都會影響開花植物花芽的正常發(fā)育[6],因此,棉花花芽發(fā)育時期、花蕾大小及其花器官的細胞發(fā)育特征成為研究棉花花發(fā)育的關鍵。其中花藥的發(fā)育過程不僅包括多種細胞形態(tài)建成,也是后期雙受精完成的先決條件。近年,部分學者研究了陸地棉的生育期[7]、花期[8]和高溫脅迫對花藥發(fā)育的影響[9],而針對花藥發(fā)育過程中細胞結構、相關發(fā)育時期[10]及受精過程的研究并不多見。其中海島棉花蕾發(fā)育與花藥細胞的形態(tài)建成、發(fā)育時期以及花粉的雙受精過程尚未報道。本研究利用半薄切片技術與顯微鏡觀察方法,確定新海35花蕾發(fā)育過程中各發(fā)育時期的細胞特征,建立花發(fā)育時期與花蕾大小的對應關系,同時對花粉萌發(fā)及受精過程進行了探究,以期為海島棉花發(fā)育時期的判定及相關細胞發(fā)育的研究提供參考。
1 材料及方法
1.1 試驗材料
試驗材料為新海35,由新疆農(nóng)業(yè)科學院經(jīng)濟作物研究所提供。2017—2018年分別種植于新疆維吾爾自治區(qū)巴音郭楞蒙古自治州庫爾勒市阿瓦提鄉(xiāng)新疆農(nóng)業(yè)科學院經(jīng)濟作物研究所育種家基地(86.08°E,41.68°N)和華中農(nóng)業(yè)大學作物遺傳改良國家重點實驗室的棉花溫室。庫爾勒大田白天最低氣溫為16℃,最高氣溫37℃;溫室最低氣溫20℃,最高氣溫36℃,溫室采用花盆單株種植,所用土壤為礫石與基質(zhì)1∶2混合,常規(guī)水肥、防蟲管理。田間管理同常規(guī)大田,定期施肥澆水。根據(jù)長期的監(jiān)測觀察,新海35號從播種到開花為61~67 d,總生育期為135~140 d。
1.2 試驗方法
1.2.1花藥取樣。在花芽發(fā)育早期進行掛牌標記,以便后期對應相同生育期取樣。2017年7月2日和2018年6月15日,分別取大田材料和溫室材料制作切片。從花芽出現(xiàn)開始,待花芽發(fā)育3~6 d后,根據(jù)花蕾發(fā)育長度,每增加1 mm取樣1次,5個重復。取回的花蕾去掉苞葉,用游標卡尺從底端蜜腺處至花蕾頂部進行長度測量、記錄、拍照,并進行解剖觀察。
1.2.2雌蕊柱頭取樣。2018年7月4-8日,在庫爾勒大田每日16:00對發(fā)育正常且長勢一致的新海35號去雄,次日10:00選取即將開放且發(fā)育一致的花蕾對前1 d去雄后留存的雌蕊柱頭進行授粉,隨后套上紙袋,避免其他花粉的污染。人工授粉后0、5、10、20、30、45 m in及1、1.5、2、4、8、10、12、14、16、18、20、22、24、26、28、30 h的雌蕊柱頭取樣;從授粉后30 h開始,之后每隔4 h對授粉的柱頭取1次,即34、38、42 h……74 h。且每次每個時間點采集5個重復,冰盒保濕帶回實驗室用于后續(xù)切片的固定。
1.2.3半薄切片制作、染色及觀察。樣品用FPA固定液(丙酸∶福爾馬林∶70%乙醇=5∶5∶90,體積比)固定;依次用30%、50%、70%、80%乙醇脫水(體積比)分別脫水15 m in,最后用無水乙醇漂洗1次。用預滲透液(無水乙醇與滲透液1∶1(體積比)混合,1 g的HarderⅠ溶于100 m L Technovit 7100制成滲透液)預滲透12 h,再用滲透液滲透24 h,然后使用包埋液(滲透液∶HarderⅡ=15∶1,體積比)進行包埋。
切片、展片、染色:將完成后熟的樹脂塊進行切片,厚度5μm。將切好的半薄切片放置在涂有多聚賴氨酸的載玻片上,吸去多余的水,將切好的花藥切片用0.5%甲苯胺藍染色。
封片、拍照:染色完成后,蒸餾水清洗3次,晾干水漬用中性樹膠封片,待樹膠完全干燥后用ZEISSAxioscope A1顯微鏡進行拍照觀察。
2 結果與分析
2.1 海島棉花蕾與花藥發(fā)育形態(tài)變化
海島棉的開花周期較長,從花芽出現(xiàn)至開花一般需要28~30 d。新海35在花芽發(fā)育3~5 d后,花蕾長度為2~3 mm(圖1B)。當花芽發(fā)育6~8 d時,花蕾長度為4~6 mm,此時萼片的發(fā)育早于花瓣,且完全包裹花瓣。花芽發(fā)育14~18 d時,花蕾長度為6~13 mm,花瓣被萼片半包裹。當花芽發(fā)育24~26 d,花蕾長度為14~21 mm,此時被萼片包裹著的花瓣發(fā)育速度明顯加快,且超過了萼片(圖1B)。隨著花蕾繼續(xù)生長,在花芽發(fā)育26~27 d,花蕾長度為21~22mm,此時萼片的發(fā)育速度較為緩慢,花瓣繼續(xù)快速生長體積變大,花瓣顏色由最初的灰白色(1~12 mm)變?yōu)榛揖G色(12~20 mm),而后為淡黃色,快開花時變?yōu)闄幟庶S色(圖1B)。花藥顏色從最初的白色(<12 mm)逐漸變?yōu)榻瘘S色(圖1B)。花絲和花柱隨著花藥的發(fā)育逐漸伸長,在開花前1天花瓣、花絲、柱頭和花藥生長迅速。在開花當天,花藥開裂,花絲快速伸長(圖1C-E),伸長的花絲在花藥開裂后將成熟的花粉粒散落至柱頭(圖1E),花粉開始萌發(fā),花粉管伸長,并進行雙受精過程。開花后1~2 d花柱不再伸長,此時花粉受精過程完成(圖1F),卵細胞“休眠”。合子時期,花瓣則變?yōu)殚偌t色(圖1F),花藥開始失水(圖1G),至此海島棉花蕾內(nèi)部花器官發(fā)育完成。

圖1 新海35花蕾及雄蕊發(fā)育形態(tài)Fig.1 The development of buds and stamen of G.barbadense Xinhai35
2.2 花蕾大小與花藥的發(fā)育時期的關系
2.2.1花原基與雄蕊原基時期。棉花的花芽分化需要經(jīng)過花原基分化、苞片分化、花萼分化、花瓣分化、雄蕊分化、雌蕊分化、藥隔形成、花粉母細胞形成、減數(shù)分裂、花粉粒形成等過程。當花芽出現(xiàn)2~3 d可以觀察到長2~3 mm的三角形苞片;花蕾發(fā)育至0.5 mm左右時,苞片生長較快,且苞片迅速增大,其生長速度明顯快于萼片。當萼片在顯微鏡下明顯可見時,萼片上部邊緣形成的突起即花瓣(圖2A),此時花瓣的頂部生長較底部迅速,且在其上部的左右兩端有可見的突起(圖2A)。當花芽出現(xiàn)3~4 d,花蕾長度0.5~1 mm時,雄蕊原基已經(jīng)出現(xiàn)(圖2B)。此時萼片生長速度比花瓣快,且萼片體積增大后包裹著其它正在發(fā)育的花器官。隨著發(fā)育的進程,花瓣向左右相互交錯覆蓋,包裹著隆起的分生組織,而分生組織內(nèi)緣上部形成的突起即雄蕊原基(圖2B)。隨著花芽發(fā)育,雄蕊上形成的突起逐漸彎向中央(圖2B)。雄蕊繼續(xù)向上生長,中央部分分化出雌蕊原基(圖2C)。
2.2.2花藥孢原細胞與花藥四分體時期。新海35花芽出現(xiàn)3~5 d,花蕾長度為2~3 mm,此時經(jīng)由雄蕊原基中間層即L2層細胞分化的孢原細胞已清晰可見(圖2D)。待花芽出現(xiàn)4~6 d,花蕾長至3~4 mm時,孢原細胞經(jīng)平周分裂形成周緣細胞與造孢細胞(圖2E)。花芽出現(xiàn)5~7 d,花蕾長至4~5 mm,此時次生周緣細胞分化出花藥的中間層和絨氈層,內(nèi)皮層出現(xiàn),裂口組織與維管組織形成,裂口區(qū)域和連接區(qū)清晰可見(圖2F)。花芽出現(xiàn)7~9 d花蕾長至5~6 mm時,經(jīng)花藥原基分化的花粉母細胞和4層花藥壁形成,花粉母細胞結構清晰,4層花藥壁從外到內(nèi)依次為:表皮層、藥室內(nèi)壁、中間層和絨氈層。此時花藥被藥隔分隔為4個藥室,花藥形態(tài)呈清晰的蝴蝶狀(圖2G)。當花芽出現(xiàn)10~12 d時,花蕾生長至6~7 mm,花粉母細胞開始進行減數(shù)分裂(圖2H),花藥中含有貯藏物質(zhì)的一層細胞開始減少,隨后細胞逐漸趨于扁平,直至發(fā)生解體。花芽出現(xiàn)12~13 d花蕾生長至7~8 mm時,花粉母細胞進入減數(shù)分裂后期。隨著花粉母細胞的逐漸成熟,可見被胼胝質(zhì)包裹、含有單倍體小孢子的四分體形成(圖2I)。當花芽出現(xiàn)13~14 d,花蕾生長至8~9 mm時,胼胝質(zhì)降解,小孢子從花粉母細胞的四分體中釋放出來,形成成熟的小孢子,至此小孢子的整個發(fā)育階段完成(圖2J)。
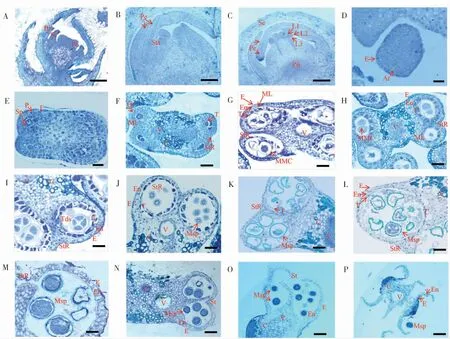
圖2 新海35花蕾長度與花藥發(fā)育時期的對應關系Fig.2 The relationship between buds length and anther development stages of Xinhai35
圖2 (續(xù))Fig.2 Continued
2.2.3花藥絨氈層發(fā)育時期。絨氈層作為種子植物花藥幼時最內(nèi)側的細胞層,能夠為小孢子的發(fā)育與外壁形成提供蛋白質(zhì)和碳水化合物等營養(yǎng)及原料,合成并分泌胼胝體酶、識別蛋白和孢粉素。當新海35的花芽出現(xiàn)5~7 d,花蕾發(fā)育至4~5 mm時,次生周緣細胞分化出絨氈層(圖2F),花芽出現(xiàn)7~9 d,花蕾為5~6 mm時,絨氈層完全形成(圖2G)。在花芽出現(xiàn)12~13 d,花蕾為7~8 mm時,絨氈層細胞出現(xiàn)液泡化,中層細胞受到擠壓,細胞質(zhì)濃度增加,細胞核體積增大,胼胝質(zhì)積累,中層細胞開始降解(圖2I)。在花芽出現(xiàn)15~17 d,花蕾長度為8~9 mm時,絨氈層合成并分泌胼胝體酶,溶解胼胝質(zhì)使小孢子從四分體中釋放。花芽發(fā)育18~22 d,花蕾9~11 mm時,來源于絨氈層的孢粉素填充在小孢子的初生外壁,小孢子體積增大,花藥膨脹生長。花蕾發(fā)育24~25 d,花蕾14~24 mm時,孢粉素積累,使花藥壁增厚,單核花粉粒經(jīng)非均等有絲分裂形成雙核花粉粒(圖2M-N),此時絨氈層完全降解。
2.2.4花粉粒的形成與釋放。海島棉花芽發(fā)育18~22 d,花蕾長度為9~11 mm時,從四分體釋放出來的小孢子只有1個細胞核,同時具有大液泡。花芽出現(xiàn)22~24 d,花蕾為11~14 mm時,絨氈層逐漸降解從而為小孢子提供營養(yǎng),同時部分蛋白在小孢子外壁積累(圖2L),形成花粉壁的前體。當花芽發(fā)育24~25 d,花蕾生長至14~24 mm時,小孢子進行有絲分裂,花藥中有豐富的內(nèi)含物,花粉粒開始膨脹生長,可以清晰地觀察花粉粒結構(圖2M)。當花蕾長度大于24 mm時,花粉粒逐漸成熟,隨后裂口組織開始降解,待花粉粒完全成熟,花藥開裂,花粉粒沿裂口區(qū)域沖出藥室,開始散粉(圖2N-O)。
2.3 花粉萌發(fā)與受精
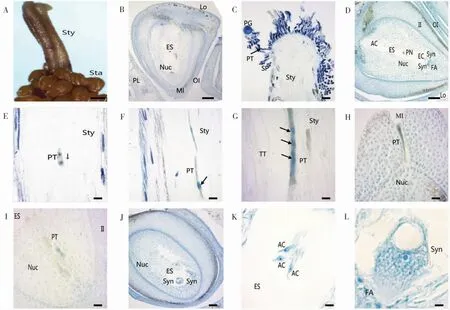
圖3 新海35花粉萌發(fā)與花粉管伸長過程Fig.3 Pollen germ ination and pollen tube elongation of Xinhai 35
2.3.1花粉粒萌發(fā)與花粉管伸長。海島棉花藥正常開裂后,花粉散出,通過人工授粉將花粉涂抹于去雄的柱頭,一定時間后取柱頭,制作切片進行觀察。經(jīng)過0.5~1 h,花粉粒開始萌發(fā),可清晰地觀察到花粉管伸出(圖3 C),并沿花柱的引導組織逐漸向下呈不規(guī)則的曲線形生長(圖3E-G)。大約在授粉后8 h,可以清晰看到花粉管中攜帶的生殖細胞發(fā)生分裂,形成前后排列的2個子細胞(圖3E)。授粉后12~14 h,花粉管伸長至花柱末端,并進入子房(圖3H)。授粉后16~20 h,花粉管已伸長到達胚珠的珠孔,并通過珠孔進入珠心(圖3H)。授粉后20~24 h,花粉管進一步到達胚囊處,并釋放出營養(yǎng)細胞和生殖細胞等內(nèi)容物(圖3 I-J)。
2.3.2雙受精過程。當人工授粉24 h后,顯微鏡觀察胚囊中清晰可見1枚精子正在與卵細胞進行受精(圖4A);另1個精子則先靠近1個極核并與之結合(圖4B),隨后與另1極核融合形成受精極核(圖4C-E),至此雙受精完成。雙受精后可以在胚囊內(nèi)清晰看見1個合子和1個助細胞(圖4E)。
2.3.3胚乳發(fā)育。海島棉的柱頭授粉后50 h,觀察到含2個游離核的胚乳(圖4D);授粉后74 h,可看見已經(jīng)形成了較多的游離核分布在胚囊內(nèi)(圖4F)。與此同時,受精卵結束休眠,胚乳進行第1次分裂。其主要特點是:基細胞大于頂細胞,基細胞靠近珠孔,細胞相對較大;而頂細胞遠離珠孔,細胞相對比較小(圖4G)。此時纖維伸長也比較明顯(圖4H)。
3 討論
3.1 棉花生長周期與花蕾形態(tài)
花器官的形態(tài)建成在植物發(fā)育過程中扮演著重要的作用,其表型不僅指示環(huán)境條件的改變,而且關系植株的生殖發(fā)育。本研究中海島棉新海35從播種到開花需要61~67 d,從現(xiàn)蕾到開花需要28~32 d,從播種到吐絮的總生育期為135~140 d。而陸地棉從現(xiàn)蕾到開花一般僅需要25~30 d[11]。從播種到吐絮的生長周期也比海島棉明顯短4~8 d[7],有研究認為,這與種質(zhì)相關基因及其棉酚含量有關[12]。新海35花蕾、萼片、花瓣和雄蕊在開花前5 d快速生長,與陸地棉的研究結果相似[13]。隨發(fā)育進程,新海35的花蕾、萼片、花瓣和雄蕊、花藥顏色隨之改變(圖1B-D),且花瓣和花藥顏色變化比陸地棉更為明顯[14]。這可能與次生代謝過程中花青素、類黃酮和花色素等相關物質(zhì)的積累有關[15]。海島棉生殖發(fā)育過程中,光合作用產(chǎn)生的干物質(zhì)以及氮素等營養(yǎng)物質(zhì)均用于生殖生長前期,導致后期積累較少,不能滿足快速發(fā)育需要,所以生育期較陸地棉長[16]。除此之外,海島棉對光照時間、光照強度及溫度有著嚴苛的要求,管理措施不當會導致開花與結鈴時期延后[17]。因此,海島棉新海35的生長周期、花蕾和花藥顏色、發(fā)育表型變化是對自然環(huán)境條件的響應,也是種質(zhì)資源、不同地理來源和遺傳變異共同作用所產(chǎn)生的結果[8,14]。
3.2 棉花花蕾大小與花藥發(fā)育時期
棉花現(xiàn)蕾后,花藥的發(fā)育需經(jīng)歷雄蕊原基突起、孢原細胞分化、1次平周分裂形成造孢細胞,再經(jīng)有絲分裂形成花粉母細胞。花粉母細胞進行減數(shù)分裂形成小孢子,小孢子形成成熟花粉粒[18-19]。海島棉H276保持系花蕾長度3~4 mm時,為花粉母細胞時期;花蕾4~5 mm時為四分體時期;花蕾長度5~6 mm為小孢子單核早期,絨氈層開始退化;花蕾6~7 mm時絨氈層完全降解,部分小孢子已經(jīng)發(fā)育成為成熟的花粉粒。花蕾>7 mm時形成成熟花粉粒[20]。而陸地棉四分體時期花蕾長度為4.1~5.5 mm,單核時期花蕾縱徑5.6~6.0 mm,成熟的花粉粒時期花蕾縱徑為8.1~10.0 mm[21]。其它研究發(fā)現(xiàn),在棉花開花期,造孢細胞時期花蕾<3.5 mm,花粉母細胞時期花蕾為3.5~5.0 mm,花粉粒時期花蕾>5 mm[22]。本研究中新海35花蕾長度至5~6 mm、7~8 mm、8~9 mm和11~14 mm分別對應花藥發(fā)育中的花粉母細胞形成期、四分體時期、小孢子釋放期和絨氈層降解時期,均比相同發(fā)育時期陸地棉的花蕾長。可能是為海島棉生殖發(fā)育后期積累營養(yǎng)物質(zhì)的要求[16]。
3.3 棉花花粉粒的發(fā)育
花粉粒的成熟是雄蕊發(fā)育的關鍵,只有花粉粒正常發(fā)育,才能保證植株的受精和結實。在雄蕊原基中初生造孢細胞分化形成花粉母細胞[23]。次生周緣細胞分化形成包裹著花粉母細胞的三層同心圓花藥壁[24],再經(jīng)對稱分裂形成中間層與絨氈層。當小孢子形成時,絨氈層細胞為其提供所需營養(yǎng)物質(zhì),并將一些遺傳物質(zhì)傳遞給小孢子[25]。小孢子的發(fā)育會直接影響花粉粒的成熟。棉花蕾期花粉粒的發(fā)育受影響會導致花粉敗育。研究證實,陸地棉花蕾處于開花前15~16 d為四分體時期至小孢子發(fā)育早期,開花前9~11 d為單核晚期至雙核早期的發(fā)育階段[26]。而新海35花芽出現(xiàn)10~13 d,花蕾生長至6~7 mm和7~8 mm時分別為花粉母細胞減數(shù)分裂和四分體時期,該時期花蕾的花藥細胞最易受非生物脅迫影響。本研究發(fā)現(xiàn),海島棉新海35在花蕾為8~9 mm時,小孢子從四分體胼胝質(zhì)中釋放出來。當花蕾11~14 mm時,絨氈層開始降解,為小孢子的發(fā)育提供營養(yǎng)物質(zhì),而細胞殘余及脂類物質(zhì)等覆蓋在小孢子表面形成花粉包被[27]。陸地棉花蕾9~14 mm時,合成并分泌孢粉素,為花粉發(fā)育提供營養(yǎng)物質(zhì)[28],而海島棉新海35則在花蕾11~14 mm完成這一過程。新海35絨氈層降解起始時的花蕾比陸地棉1355B稍大[10],可能是由于溫度、光照及種質(zhì)資源的相關調(diào)控基因改變了絨氈層細胞程序性死亡的時間[29]。因此,海島棉花蕾長度5~6 mm和7~8 mm對應花粉母細胞和四分體時期的正常發(fā)育,在花蕾8~9 mm和11~14 mm時絨氈層能否進行正常降解關系著花粉粒成熟和花藥的正常開裂[30]。
3.4 棉花花粉萌發(fā)及受精過程
在陸地棉中雙受精是精卵融合以及與極核的融合。其中,極核融合的機制是:外膜在多個點上的直接融合,即內(nèi)質(zhì)網(wǎng)的融合;隨后是內(nèi)核膜的融合形成橋,隨后橋擴大并愈合,逆向釋放被網(wǎng)羅的細胞質(zhì)成分,最后形成新的完整的核膜,完成核的融合[16]。精卵細胞的融合與極核的融合方式基本一致[25]。本研究中,海島棉花粉粒在柱頭上經(jīng)過0.5~1 h后開始萌發(fā),花粉管開始生長。授粉后8 h,可以清晰看到花粉管中攜帶的生殖細胞發(fā)生分裂。授粉后20~24 h,花粉管到達胚囊。授粉后24 h,精子與卵細胞進行受精。授粉50 h后,形成游離胚。74 h,胚乳進行第一次分裂,此時纖維明顯伸長。有研究發(fā)現(xiàn),授粉后12 h和24 h觀察到的花粉管數(shù)量基本一致,且30℃最適于花粉管萌發(fā),花柱中花粉管的平均數(shù)量最多[31]。研究發(fā)現(xiàn),花粉萌發(fā)率可作為響應溫度的主要指標[32]。在30℃培養(yǎng)條件下花粉萌發(fā)率最高,25℃培養(yǎng)條件下的花粉管長度最長。不同溫度條件不同材料的花粉萌發(fā)率和花粉管長度不同。同時花粉萌發(fā)率和花粉管長度也反映了單株鈴數(shù)和成鈴率[33]。有研究發(fā)現(xiàn),授粉后15 h花粉管通過珠孔進入胚囊,釋放內(nèi)含物。48 h后可見胚的分裂。72 h后可以明顯觀察到分裂的早期胚[34]。但在新海35中授粉50 h后,才觀察到纖維發(fā)育,這可能是因為海島棉的受精過程中,極核的受精是精子先與1個極核結合,然后再與另1個極核進行核膜、核質(zhì)的融合,因此所需要受精時間較長。這也證實了海島棉的受精時間比陸地棉長[25]。雙受精完成之后,棉花纖維進入伸長期,與此同時纖維細胞出現(xiàn)大液泡[35]。這一時期是遺傳物質(zhì)傳遞的關鍵時期,環(huán)境條件及相關基因的作用都會影響海島棉受精時間。
4 結論
海島棉新海35花藥顏色和花藥細胞隨著花蕾發(fā)育呈現(xiàn)動態(tài)變化。在相同的花藥細胞發(fā)育階段,新海35的花蕾比陸地棉的花蕾更長,且各發(fā)育階段所需的時間更長。結合花芽發(fā)育時間、花蕾形態(tài)學及細胞學觀察,確定了新海35花粉母細胞形成期、四分體時期、小孢子釋放期和絨氈層降解時期花蕾長度分別為5~6 mm、7~8 mm、8~9 mm和11~14 mm階段。同時新海35授粉后,花粉粒萌發(fā)需要0.5~1 h,花粉管延伸到子房需要12~14 h,而受精過程則是在授粉后24 h完成,而授粉后50~74 h胚乳開始發(fā)育并伴隨著纖維的發(fā)育。
