陸地棉纖維發育相關基因GhEXPs的分析及表達研究
2021-06-19 08:39:36呂麗敏左東云王省芬張友平程海亮王巧連宋國立馬峙英
棉花學報 2021年3期
呂麗敏,左東云,王省芬,張友平,程海亮,王巧連,宋國立,4*,馬峙英*
(1.河北農業大學/棉花生物學國家重點實驗室河北基地,河北 保定071001;2.中國農業科學院棉花研究所/棉花生物學國家重點實驗室,河南 安陽455000;3.河北農業大學/教育部華北作物種質資源研究與利用重點實驗室,河北 保定071001;4.鄭州大學/棉花生物學國家重點實驗室鄭州基地,鄭州450001)
擴展蛋白(Expansin)是一類重要的細胞壁蛋白,在酸性條件下它能松弛細胞壁,促使細胞增大[1-2]。擴展蛋白在細菌、真菌和高等植物中以多基因家族的形式廣泛存在,在促進細胞伸長、莖尖和根伸長、葉片的形態建成、花和果實的發育、胚胎發生、花粉發育和花粉管生長等過程中發揮著重要作用[3-6]。McQueen-Mason團隊最早從黃瓜幼苗中分離出有活性的擴展蛋白,研究表明該蛋白在酸性、離體條件下能夠促進下胚軸細胞的伸長[7]。
近年來,隨著分子生物學的快速進步和高通量測序技術的發展,科學家從植物和微生物中鑒定了多個物種的擴展蛋白基因家族。通過對擬南芥[9]、大豆[10]、水稻[9]、大白菜[11]、楊樹[12]、煙草[13]等植物擴展蛋白的研究發現,植物類擴展蛋白家族包括4個亞家族,分別為α-擴展蛋白(EXPA)、β-擴展蛋白(EXPB)、類α-擴展蛋白(expansin-like A,EXLA)和類β-擴展蛋白(expansin-like B,EXLB)[8]。該家族蛋白通常包含250~275個氨基酸殘基,氮端有包含20~30個氨基酸的信號肽,屬于分泌蛋白[3]。擴展蛋白包括2個典型的結構域,分別是位于氮端的DPBB(Double-psi betabarrel)和碳端的CBM 63(Fam ily-63 carbohydrate binding module),其中DPBB是1個六鏈雙“ψ”β-桶形結構,CBM 63是碳水化合物結合模塊家族-63結構域[4]。結構分析表明,2個擴展蛋白分子中間夾1個細胞壁多糖形成“三明治”式的結構,該結構是擴展蛋白促使細胞壁發生松弛的作用模型[3]。
棉花纖維是胚珠表皮細胞分化伸長的單細胞結構,它的特殊伸長機制一直是生物學研究的熱點,棉花纖維是研究單細胞伸長的模式系統[14]。棉花纖維發育過程分為5個時期,分別是起始期、伸長期、轉換期、次生壁合成期和成熟期,各個時期之間存在交互重疊,沒有嚴格的界限[15]。擴展蛋白參與棉花纖維細胞伸長發育的生物學過程已取得了一些進展[16-18],科學家利用棉花纖維的cDNA(Complementary DNA,互補DNA)文庫、基因芯片、SSH(Suppression subtractive hybridization,抑制差減雜交)文庫等技術從棉花中篩選出在纖維優勢表達的擴展蛋白基因[19-20],發現EXPA亞家族的2個基因GhExp1和GhExp2在棉花纖維中高表達[21],后來證明這2個基因是位于棉花A亞組和D亞組的1對同源基因[22]。在陸地棉中過表達GhEXPA8基因能顯著促進纖維的伸長[23];序列比對發現GhEXPA8和上述的GhExp2是同一個基因。通過對陸地棉擴展蛋白全基因組分析,采用系統命名法將該基因命名為GhEXPA2,染色體定位分析發現它位于D亞組[22]。此外,從海島棉A亞組獲得了1個擴展蛋白基因GbEXPATR,屬于EXPA亞家族,過表達該基因能夠使棉纖維變得更長、更細,強度更強。與陸地棉GhEXPA2相比,GbEXPATR缺少CBM 63結構域,這可能與它在海島棉中的特殊功能有關[16]。近來,對陸地棉擴展蛋白基因家族的研究鑒定了93個GhEXPs基因,發現一些基因在纖維發育中上調表達[22]。這些研究表明,擴展蛋白在纖維發育過程中發揮重要作用。
早期科學家認為擴展蛋白通過減弱多糖和纖維素微纖絲之間的連接作用,從而使細胞壁松弛[2],也可能是通過清除纖維素和多糖之間的氫鍵使細胞壁松弛[8]。隨著研究的不斷深入,科學家發現細胞壁中的纖維素微纖絲呈束狀分層排列,并把纖維素微纖絲交叉的位置稱為“生物力學熱點(biomechanical hotspots)”,該區域存在的細胞壁多糖使纖維素微纖絲連接在一起[24]。目前,科學家已經獲得了細菌擴展蛋白 EXLX1(Expansin-Like X)和玉米EXPB亞家族的擴展蛋白EXPB1(Zea m1)的晶體結構。研究表明,芳香族氨基酸苯丙氨酸、色氨酸和酪氨酸是擴展蛋白和多糖底物相互結合的關鍵位點,并且十分保守[25-26]。擴展蛋白晶體結構的解析為我們深入了解其生物學功能和機制奠定了堅實基礎。
為進一步闡釋棉花纖維伸長的作用機制,解析擴展蛋白在棉花纖維發育過程中發揮的作用。本文從陸地棉轉錄組數據中篩選了4個在纖維發育過程中優勢表達的擴展蛋白基因GhEX PA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a,對這4個GhEXPs基因的進化關系、蛋白結構及表達模式等方面進行了研究。
1 材料與方法
1.1 材料
試驗材料陸地棉(Gossypium hirsutumL.)標準系TM-1是來源于中國農業科學院種質資源中期庫的資源,本課題組種植于中國農業科學院老所部試驗基地(河南省安陽縣,36°06′84.44″N,114°49′61.5″E)保存使用,田間常規管理。在盛花期對試驗材料進行掛牌,分別于開花當天(0 day post anthesis,0 DPA)、3 DPA、5 DPA、7 DPA、10 DPA、15 DPA、20 DPA和30 DPA的胚珠和纖維進行取樣,5 DPA及之前時間點的樣品是胚珠和纖維未分離的材料,其余時間點的樣品均是從胚珠上剝離的纖維材料。冰上取材后立即放入液氮速凍,于-80℃超低溫冰箱保存備用。
1.2 方法
1.2.1 總RNA的提取和c DNA制備
RNA提取按照多糖多酚植物總RNA提取試劑盒(貨號:DP441,北京天根生化科技有限公司)說明書及參考相關文獻進行操作[22]。
提取的RNA放置-80℃保存備用或直接進行反轉錄,獲得的cDNA用超純水稀釋10倍后,-20℃冰箱保存,用于后面實驗。步驟參考Takara公司的PrimeScriptTMII 1st Strand cDNA Synthesis Kit(貨號:6210A)的說明書及相關文獻[22]。
1.2.2 棉花GhEXPs基因的克隆
從棉花基因組數據庫(https://cottonfgd.org/)[27]獲得4個GhEXPs的開放閱讀框,分別為Ghir_D05G018790.1(GhEXPA1d)、Ghir_A04G009030.1(GhEXPA4b)、Ghir_A05G038370.1(GhEXPA23b)和Ghir_A12G014930.1(Gh-EXPB3a)。設計引物擴增每個基因的開放閱讀框(Open read frame,ORF),然后把序列連到pMDTM18-T載體上進行測序驗證。基因克隆所用的引物序列見表1。

表1 基因開放閱讀框擴增的PCR引物Table 1 The primers for GhEXPs gene open read frame am plification
1.2.3 棉花GhEXPs基因的進化分析及三維結構預測建模
利用MEGA 6.0軟件對4個GhEXPs蛋白以及其它物種的擴展蛋白進行多重序列比對并構建系統進化樹[28]。構建進化樹所用擬南芥蛋白序列來自擬南芥TAIR網站(https://www.arabidopsis.org/),陸地棉擴展蛋白序列來自相關報道[22],其他物種的擴展蛋白序列來自于NCBI數據庫,用4個GhEXPs蛋白在NCBI數據庫進行BLAST。選擇了包括單子葉、雙子葉植物以及擴展蛋白不同亞家族成員,包括草莓:FaExp2(AF159563),歐洲甜櫻桃:PraExp1(AF297521)、PraExp2(AF297522),黃瓜:CsExp1(U30382)、CsExp2(U30460),番茄:LeExp1(U82123)、LeExp2(AF096776)、LeExp3(AF059487)、LeExp4(AF059488)、LeExp5(AF0-59489)、LeExp8(AF184232),水稻:OsExp1(Y07782)、OsExp4(U85246),獨 腳 金:SaExp3(AF291659),百日草:ZeExp3(AF230333),歐洲油菜:BnExp1(AJ000885),煙草:NtExp1(AF04-9350.1),豌豆:PsExp1(X85187),小麥:TaEXPA2(AAS48871.1),玉米:EXPB1_Zea m1(AAO45-608.1)。
將4個GhEXPs蛋白序列去掉信號肽后,利用 在 線 軟 件 Phyre2(http://www.sbg.bio.ic.ac.uk/phyre2/htm l/page.cgi?id=index)進行三維結構模型預測[29],然后利用軟件PyMOL V2.3.4對蛋白模型進行可視化處理,再用Adobe Illustrator CS3 13.0.0編輯圖片標注關鍵氨基酸信息。
1.2.4 轉錄表達分析
從0 DPA、3 DPA、5 DPA、7 DPA、10 DPA、15 DPA、20 DPA和30 DPA纖維和胚珠樣品中提取的總RNA進行轉錄組測序。此外,從Cotton FGD(Cotton Functional Genom icsDatabase,https://cottonfgd.org/)[27]數據庫中下載了其它組織樣品的RNA-Seq數據,樣品包括:根、莖、葉、苞葉、花瓣、雄蕊、雌蕊和花托。測序結果的基因表達量用每千個堿基的轉錄每百萬映射讀取的片段數(Fragments per kilobase of m illion mapped reads,FPKM)表示。
1.2.5 目的基因的表達分析
通過qRT-PCR(Quantitative real-time polymerase chain reaction,實時定量PCR)檢測0 DPA、3 DPA、5 DPA、7 DPA、10 DPA、15 DPA、20 DPA和30 DPA不同時期的棉花胚珠和纖維樣品中4個GhEXPs基因的表達情況。以UBQ7(GenBank登錄號:AY189972)為內參基因,引物見表2。試驗所用試劑盒為Takara公司生產的TB GreenTMPrem ix Ex TaqTMII Kit(貨號:RR820A),采用美國ABI的QuantStudioTM5熒光定量PCR儀。采用2-ΔΔCT方法計算基因的相對表達量[30]。利用Excel 2010進行差異顯著性分析。
2 結果與分析
2.1 陸地棉4個GhEXPs基因的克隆及進化分析
本研究中4個GhEXPs基因(GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a)開放閱讀框分別為768 bp、795 bp、798 bp和804 bp。將GhEXPA1d、GhEXPA4b、GhEXPA23b和Gh-EXPB3a與棉屬及其他物種的擴展蛋白一起構建系統進化樹(圖1),發現GhEXPA4b、EXPA1d和GhEXPA23b分別屬于EXPA亞家族的第Ⅳ、Ⅴ和Ⅻ亞組,即EXPA-Ⅳ、EXPA-Ⅴ和EXPA-Ⅻ;GhEXPB3a屬于EXPB亞家族的第Ⅱ亞組,即EXPB-Ⅱ(圖1)。

表2 定量PCR的引物Table 2 The primers for qRT-PCR
2.2 陸地棉4個GhEXPs蛋白的三維建模及關鍵氨基酸位點分析
目前植物中僅有玉米EXPB1和細菌中的EXLX1兩個擴展蛋白解析了晶體結構,位于EXPB1和EXLX1三維結構表面的芳香族氨基酸是它們和細胞壁多糖結合的關鍵位點。為尋找棉花纖維細胞壁中擴展蛋白的關鍵氨基酸位點,本研究把陸地棉的4個GhEXPs蛋白與上述2個蛋白進行序列比對(圖2),鑒定關鍵的氨基酸位點。結果發現,在DPBB結構域中,棉花4個擴展蛋白GhEXPs與玉米EXPB1的色氨酸W 26和酪氨酸Y27對應的位點也是2個保守的芳香族氨基酸,其中GhEXPB3a與玉米EXPB1對應位點的氨基酸完全相同,其余3個GhEXPs與W 26位點對應的氨基為苯丙氨酸(F),4個Gh-EXPs與Y27對應的位點沒有發生變異。在CMB63結構域中,玉米擴展蛋白EXPB1的關鍵位點是Y 160和W 194,4個GhEXPs蛋白與Y160對應的位點變異為W或F(GhEXPA23b除外),與W 194對應的位點完全相同,沒有發生變異(圖2)。同時,4個GhEXPs與微生物擴展蛋白EXLX1也進行了比對,EXLX1中兩個連續的色氨酸W 125、W 126是該蛋白的關鍵氨基酸位點,4個GhEXPs與W 125、W 126對應的位點變異為YF(GhEXPA23b與W 126對應的位點除外)2個芳香族氨基酸(圖2)。

圖1 4個GhEXPs蛋白與其它植物擴展蛋白的聚類分析Fig.1 Phylogenetic tree of the four GhEXPs and other plant expansins
利用在線軟件建立了陸地棉4個擴展蛋白的三維結構模型,參考模型是玉米EXPB1(模板編號:c2hczX)的結構,建模結果如圖3所示。GhEXPA1d、GhEXPA4b、GhEXPA23b和Gh-EXPB3a與玉米EXPB1的序列一致性分別為31%、34%、32%和42%,模型可信度均為100%。從三維結構模型的預測結果來看,GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a蛋白均包含DPBB和CBM 63兩個結構域,其中DPBB是典型的六鏈雙“ψ”β-桶結構,2個結構域上分布有芳香族氨基酸苯丙氨酸、酪氨酸和色氨酸。我們把序列比對得到的關鍵芳香族氨基酸標注在三維結構圖中(圖3),在DPBB結構域中,Gh-EXPA1d的F13、Y14,GhEXPA4b的F18、Y19,GhEXPA23b的F22、Y23和GhEXPB3a的W 19、Y20位于結構的表面;而GhEXPA1d的W 183、F150,GhEXPA4b的W 187、F154,GhEXPA23b的W 190,Y155和GhEXPB3a的W 188、Y154位于CBM 63結構域的表面(圖3)。

圖2 4個GhEXPs與EXLX1和EXPB1的序列比對Fig.2 Multiple sequence alignment of 4 GhEXPs and EXLX1 and EXPB1
從三維結構圖可以看出,每個擴展蛋白均有4個芳香族氨基酸分布在同一平面上,GhEXPA1d包括F13、Y14、F150和W 183,GhEXPA4b包括F18、Y19、F154和W 187,GhEXPA23b包括F22、Y23、W 190和Y155,GhEXPB3a包括W 19、Y20、W 154和W 188(圖3),但GhEXPA1d、Gh-EXPA4b和GhEXPB3a中與EXLX1的W 125對應的芳香族氨基酸和它們并不在一個平面上(圖3)。而這些位于同一平面的保守氨基酸在細胞壁中與多糖結合可能是松弛細胞壁的關鍵。
2.3 陸地棉中4個GhEXPs基因的組織表達分析
根據轉錄組測序結果分析4個GhEXPs基因在棉花纖維發育不同時期及其他組織的表達情況(圖4)。總體來看,GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a在胚珠和纖維中優勢表達,在其他組織包括根、莖、葉、苞葉、花瓣、雄蕊、雌蕊和花托表達水平很低。其中GhEXPA1d和GhEXPA23b這兩個基因在纖維中特異表達,在其他組織中基本不表達;GhEXPA4b和Gh-EXPB3a在纖維之外的其它組織如葉片、雄蕊、雌蕊等有較低水平的表達。轉錄組結果初步分析表明,4個GhEXPs是在纖維發育中明顯優勢表達的基因。

圖3 陸地棉4個GhEXPs蛋白的三維結構預測Fig.3 The predicted three-dimensional structures of 4 GhEXPs
2.4 qRT-PCR分析驗證4個GhEXPs基因的表達情況
為進一步驗證4個陸地棉擴展蛋白基因的表達情況,利用qRT-PCR技術對4個GhEXPs基因的表達進行了驗證。從圖5可以看出,4個GhEXPs基因在纖維發育不同時期的表達模式和轉錄測序的結果基本一致(圖4和圖5)。GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a均在纖維發育的起始期和伸長期上調表達。其中GhEXPA1d和GhEXPB3a在開花當天(0 DPA)表達量相對較高,他們分別在開花后10 d和20 d的表達水平急劇下降(圖5);GhEXPA4b和Gh-EXPA23b在花后3~7 d表達量高,然后表達量開始下降(圖5)。此外,EXPA亞家族的3個基因在纖維中的表達情況也不相同,GhEXPA1d和GhEXPA4b的表達水平明顯高于GhEXPA23b。
3 討論
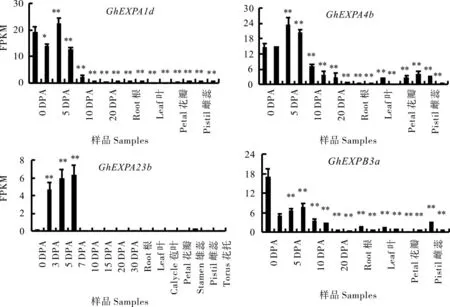
圖4 陸地棉4個GhEXPs基因在不同纖維樣品和組織中的轉錄組分析Fig.4 The transcriptome analysis of 4 GhEXPs from fiber and other tissues
本研究鑒定了4個在棉花纖維中特異表達的擴展蛋白基因,其中3個屬于EXPA亞家族,1個屬于EXPB亞家族,這與陸地棉擴展蛋白基因EXPA亞家族占優勢的結果一致[22]。蛋白的一級結構是正確行使功能的基礎。在蛋白序列一級結構的DPBB結構域上,對比棉花4個GhEXPs與玉米EXPB1、細菌EXLX1的關鍵保守位點發現,在這些關鍵位點也都是芳香族氨基酸,暗示了它們可能與玉米EXPB1和細菌EXLX1具有相似的功能,但個別氨基酸的差異,又凸顯了物種之間的不同。
最近利用固態核磁共振(Solid-state nuclear magnetic resonance,ss-NMR)技術揭示了EXPB1蛋白結合的底物是半纖維素葡萄糖醛阿拉伯聚糖(GAX,glucuronoarabinoxylan)[31],點突變試驗表明了EXLX1的結合底物主要是纖維六糖[3,32]。在本研究中,根據同源序列比對和建模分析,我們發現陸地棉4個GhEXPs與已報道的解析出晶體結構的玉米擴展蛋白EXPB1的關鍵氨基酸位點基本相同,并與細菌中已經解析結構的EXLX1蛋白中在關鍵的芳香族氨基酸位點也有較高的一致性。由此推測本研究發現的棉花擴展蛋白可能具有類似的功能。另有研究發現,細胞壁纖維素在聚合粘連位置形成“生物力學熱點”,在這些位點起黏合作用的就是半纖維素成分,擴展蛋白就是通過結合這些半纖維分子把它們從纖維素上解鎖從而松弛細胞壁的[24]。棉花纖維細胞壁的半纖維素成分有木葡聚糖、甘露聚糖和葡萄糖醛阿拉伯聚糖(GAX)等[15]。我們推測棉花擴展蛋白也可能是通過與棉花纖維細胞中的半纖維成分結合而使細胞壁發生松弛,從而在內部膨壓的驅動下使纖維細胞不斷擴展。然而,棉花擴展蛋白在纖維細胞壁內的生物機制以及具體結合的半纖維種類仍需要進一步深入研究。

圖5 陸地棉4個GhEXPs基因的qRT-PCR分析Fig.5 qRT-PCR analysis of 4 GhEXPs in upland cotton
擴展蛋白廣泛分布在植物和微生物的細胞壁中,通過松弛細胞壁促進細胞的擴展[3,24,33]。研究表明,外加細胞壁蛋白的提取物能夠促進細胞伸長[7]。擬南芥根毛特異基因AtEXPA7的RNAi(RNA interference,RNA干擾)試驗可明顯降低根毛細胞的長度,與野生型對照相比,RNAi株系根毛長度減少了25%~48%[6]。擴展蛋白在參與棉花纖維發育也有諸多報道[16-18],GhEXPA1(即GhExp1[21])在開花后6~28 d的纖維中特異表達且是表達水平高的基因,Xu等[18]研究表明,GhEXPA1和GhRDL1相互作用共同調控纖維的伸長。Li等[16]獲得海島棉擴展蛋白基因GbEXPATR,過表達該基因能明顯促進棉花纖維的伸長。擴展蛋白基因具有明顯的時空和組織表達特異性。在棉花纖維發育中已經鑒定了位于A亞組和D亞組上的一對同源擴展蛋白基因GhExp1和GhExp2[21],RNA印記實驗已經證明二者在纖維發育不同時期高表達,而在根、莖、葉等組織中不表達,超表達GhEXPA8(即GhExp2)基因可以明顯促進棉纖維的伸長[23,25]。本研究篩選獲得的陸地棉擴展蛋白基因GhEXPA1d、GhEXPA4b、GhEXPA23b和GhEXPB3a也是在棉花纖維中特異表達,在根、莖、葉等組織中表達量低(圖4),推測這些擴展蛋白基因在促進纖維伸長發育過程中也可能發揮著重要的作用。
4 結論
本文從陸地棉中獲得了4個在纖維中特異表達的擴展蛋白基因,分別是GhEXPA1d、Gh-EXPA4b、GhEXPA23b和GhEXPB3a,前3個基因屬于EXPA亞家族,GhEXPB3a屬于EXPB亞家族。其編碼的蛋白均含有DBPP和CBM 63兩個保守結構域。三維結構建模分析發現,芳香族氨基酸苯丙氨酸、色氨酸和酪氨酸分布在擴展蛋白2個結構域表面的4個位點,這些氨基酸處于同一平面上,可能是底物結合的關鍵位點。4個基因在根、莖、葉、苞葉等組織中表達量低,在棉花纖維組織發育的前期優勢表達。本研究為闡明擴展蛋白基因對纖維發育的調控提供了理論依據。
