5-羥色胺信號系統的研究概況
2021-06-29 08:00:34劉啟鴻胡劍云王寧馨陳盈胡露楠柯曉
醫學綜述 2021年11期
劉啟鴻,胡劍云,王寧馨,陳盈,胡露楠,柯曉
(福建中醫藥大學附屬第二人民醫院脾胃病科,福州350003)
5-羥色胺(5-hydroxytryptamine,5-HT)是一種重要的單胺類神經遞質,在化學上屬于吲哚胺類化合物,是人體的內源性活性物質,與人體的一系列行為相關,同時也與性格和情感有關。5-HT最早由Rapport[1]從血清中分離出來,因此又稱為血清素,其大量存在于消化系統,少量存在于中樞神經系統,主要作用為參與人和動物中樞的情緒、學習、記憶、情感、覺醒等活動,以及胃腸道的運動、分泌、免疫等功能的調節[2]。5-HT的作用多種多樣,其生成、釋放、滅活與代謝等過程中任一環節出現問題均可能導致中樞和外周功能的紊亂,不僅存在于功能性疾病,也與器質性疾病有密切聯系。近年來,5-HT受體及其亞型逐漸被發現,截至目前發現的18個基因編碼超過15種哺乳動物的5-HT受體,根據信號轉導機制不同可分為7個不同家族,每個家族根據作用機制不同又分為不同的亞型[3],5-HT與不同受體的亞型結合發揮不同的生物功能效應。現就5-HT信號系統的研究概況予以綜述,通過闡述各個受體及相關亞型與疾病的關系,研究5-HT受體及亞型之間的相互作用,從而為臨床疾病的治療與新藥研發提供理論依據。
1 5-HT的來源與合成
機體中約95%的5-HT來自消化道黏膜,其中91%位于腸黏膜的腸嗜鉻細胞,9%存在于腸道神經元[4]。5-HT合成的主要原料為色氨酸,色氨酸是人體的必需氨基酸,須從食物中攝取吸收,外周系統中的5-HT很難透過血腦屏障進入中樞,但色氨酸可進入血腦屏障,因此中樞與外周的5-HT合成互不影響[5]。具體為:①外周5-HT的合成途徑。色氨酸在腸嗜鉻細胞內經過色氨酸羥化酶(tryptophan hydroxylase,TPH)1作用產生5-羥色氨酸,5-羥色氨酸在左旋芳香氨基酸脫羧酶的作用下生成5-HT,并存在于腸嗜鉻細胞。②中樞5-HT的合成途徑。色氨酸借助載體通過血腦屏障進入大腦,在中腦神經核團(中縫背核)分布的5-HT能神經元內借助TPH2催化生成5-羥色氨酸,5-羥色氨酸經過左旋芳香氨基酸脫羧酶的脫羧作用形成5-HT,并儲存于5-HT能神經元末梢的囊泡中。TPH是5-HT合成過程中的關鍵酶,包含TPH1與TPH2兩種類型,其中TPH1主要存在于腸嗜鉻細胞和腸壁肌間神經叢中的5-HT能神經元內,TPH2主要存在于中樞神經系統。可見,5-HT的合成可在腸嗜鉻細胞、腸神經系統和中樞神經系統同時進行。詳細合成途徑見圖1。
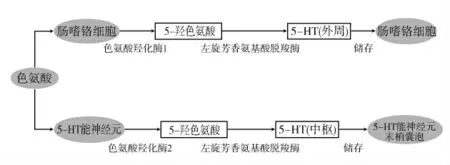
圖1 5-HT的合成途徑
2 5-HT的代謝
2.1 5-HT的釋放 5-HT的代謝動力學通過細胞水平來反映。其中,外周合成的5-HT通過轉化成囊泡顆粒的形式聚集在腸嗜鉻細胞細胞膜下,其表面的微絨毛是感受器,當感受到腸管運動擴張、迷走神經興奮及腸腔內的各種刺激時,立即將理化信息轉變為內分泌信息,從而促使5-HT的釋放[6]。而中樞神經元內的5-HT在神經沖動的驅使下,釋放到突觸間隙中與不同特異性5-HT受體結合,產生不同的生物學作用。5-HT能發揮生物學效應主要借助于G蛋白偶聯受體與離子通道偶聯受體兩大受體,G蛋白偶聯受體或離子通道偶聯受體通過與細胞或神經元上的5-HT受體結合,從而誘發細胞或神經元內反應[7]。
2.2 5-HT的滅活與再攝取 5-HT產生后進入肝門靜脈,經肝臟時部分被降解,剩余的經過肺循環時又有50%~80%被肺內皮細胞攝取,在線粒體外膜上的單胺氧化酶催化作用下形成醛類化合物,醛類化合物經過醛縮酶水解形成5-羥吲哚乙酸,通過尿液排出體外[8]。5-HT的再攝取過程是由神經元或細胞膜上的5-HT轉運體決定。5-HT轉運體是一種跨膜轉運蛋白,對5-HT具有高度選擇的親和力,負責從神經突觸間隙中重新攝取5-HT進入突觸前神經元,直接影響突觸間隙5-HT的水平,并間接反映神經纖維末梢5-HT的數量,其作用機制主要為利用Na+/K+/Cl-的跨膜梯度進行傳遞,在血小板、平滑肌細胞、肥大細胞、突觸小體等均有分布,主要分布于腸道黏膜層、肌間神經叢以及大腦邊緣系統神經元細胞膜上,但5-HT轉運體的作用可以被5-HT再攝取抑制劑抑制,因此臨床常通過研發選擇性5-HT再攝取抑制劑、5-HT和去甲腎上腺素再攝取抑制劑來治療焦慮、抑郁等精神類疾病[9]。
3 5-HT受體及其亞型
5-HT及其受體廣泛存在于腸神經系統、中樞神經系統及胃腸道、心血管和血小板等非神經組織中,調控人體的胃腸道運動、情緒等生理活動。目前發現5-HT至少存在7個受體,這7個受體又包括30多種受體亞型,研究較多的主要為5-HT1、5-HT2、5-HT3、5-HT4、5-HT5、5-HT6、5-HT7,其中5-HT1、5-HT2、5-HT3、5-HT4、5-HT7主要影響胃腸道的功能,而5-HT5、5-HT6主要與神經精神系統疾病相關,不同的受體亞型作用于效應器上會產生不同的生物學效應。
3.1 5-HT1受體 5-HT1受體包括5-HT1A、5-HT1B、5-HT1C、5-HT1D、5-HT1E、5-HT1F以及5-HT1P受體7種亞型,是5-HT信號系統中受體最多的亞型[10]。其中,5-HT1A受體主要分布在大腦邊緣系統及腸神經元的肌間或黏膜下神經叢內,通過借助G蛋白偶聯受體作用,降低腺苷酸環化酶(adenylate cyclase,AC)的活性,間接降低環腺苷酸(cyclic adenosine monophosphate,cAMP)合成能力,阻礙突觸后電位的興奮性,引起平滑肌松弛。有研究發現,5-HT1A受體參與致癌作用,如前列腺癌、膀胱癌、小細胞肺癌等,而5-HT1A受體拮抗劑能夠阻斷5-HT誘導的腫瘤生長[11]。5-HT1B和5-HT1D受體主要分布于大腦基底節與黑質,位于突觸前膜上,其作用機制可能為通過負反饋機制起到抑制神經遞質釋放的作用[12]。此外,有報道腦血管中也分布有5-HT1D受體,其發揮收縮血管的作用,該受體減少會引起血管舒張,容易導致偏頭痛,因此臨床常用5-HT1D受體激動劑治療偏頭痛,使顱內動脈血管收縮,血液重新分布,從而使偏頭痛得以改善[13]。5-HT1C受體主要分布于脈絡膜叢中的上皮細胞,作用在邊緣系統,與G蛋白偶聯進行信號連接,激動磷脂酶C使磷酸肌醇水解引發細胞反應,從而影響機體的睡眠、飲食、體溫、性行為以及神經內分泌等功能[14]。5-HT1E受體主要分布在內嗅皮質、殼核尾狀核和隱窩中,通過刺激AC的活性,增加第二信使cAMP合成,興奮突觸后電位,收縮血管平滑肌[15]。5-HT1F受體主要分布于大腦皮質、小腦、海馬,也存在于三叉神經脈管系統,其不僅在神經元上表達,也可在神經膠質細胞中表達,通過抑制降鈣素基因相關肽的釋放,阻礙三叉神經尾核內神經元信號傳遞,起到抗偏頭痛的作用[16]。5-HT1P受體主要分布在腸神經系統,其通過介導腸神經元對5-HT的慢去極化,誘發腸蠕動和分泌反射[17]。
3.2 5-HT2受體 5-HT2受體包括5-HT2A、5-HT2B和5-HT2C受體3種亞型。5-HT2A受體主要分布于血小板、平滑肌以及大腦皮質。有研究發現,5-HT2A受體可抑制前邊緣皮質部位5-HT的釋放,主要機制可能通過激活5-HT2A受體誘發γ-氨基丁酸/甘氨酸的釋放,并與單胺神經纖維的軸突形成突觸結構,從而產生抑制作用;同時突觸前膜上的5-HT2A受體能夠調節谷氨酸能的傳遞和改善機體的認知功能[18]。5-HT2B受體位于中樞神經系統、心血管系統和胃腸道中,是治療偏頭痛、肥胖和腸易激綜合征的重要靶標。有研究表明,5-HT2B受體拮抗劑(RS-127445)通過作用于結腸平滑肌上的5-HT2B受體競爭抑制結腸的運動;相反,若激動肌間神經叢上5-HT2B受體則能夠起到促進結腸平滑肌收縮的作用[19]。5-HT2C受體在大腦中高表達,主要位于大腦紋狀體和下丘腦的多巴胺能神經元和去甲腎上腺素能神經元。目前研究發現,5-HT2C受體與肥胖、疼痛、成癮、癲癇和自殺行為等有關,其機制可能是激活谷氨酸能或γ-氨基丁酸能神經元起到抑制5-HT的作用,也可能通過抑制額葉皮質和大腦邊緣系統的多巴胺能神經的活性,從而參與精神分裂癥的病理機制[20]。
3.3 5-HT3受體 5-HT3受體廣泛分布于中樞及外周神經系統,是唯一一個借助配體門控陰離子通道的亞型,是引起嘔吐、痛覺、精神分裂和腸道動力異常的重要受體[21]。在中樞神經系統中,5-HT3受體主要分布于大腦皮質、海馬、尾狀核等部位;在人的結腸中,其主要分布于黏膜下層與肌層的神經叢。5-HT3受體包括A、B、C、D、E五種亞型,其中A和B是最主要的亞型,且A亞型為離子通道的功能性受體,而其他亞型均不能形成單獨的功能性受體,D和E亞型只分布在胃腸道中。5-HT3受體是控制消化功能障礙(如厭食癥和貪食癥)的重要靶標,而5-HT3受體拮抗劑可有效抵抗飲食失調以及早期化學療法和放療引起的嘔吐[22]。5-HT3受體激活后可使細胞內的Ca2+增加,同時促進神經遞質與神經肽的釋放,起到興奮中樞、外周神經元的作用,引起嘔吐與炎癥反應。因此,臨床常通過研發5-HT3受體拮抗劑來抑制多巴胺能神經元、促使促腎上腺皮質激素水平升高起到止吐和減輕炎癥反應的作用。5-HT3受體的激活可以抑制癲癇發作,抑制5-HT3受體則會導致腦電圖尖峰波的出現,誘發癲癇[23]。
3.4 5-HT4受體 5-HT4受體主要分布在中樞與外周組織中,其主要作用為調節神經遞質的釋放與神經元離子通道的活性。在中樞神經系統中,5-HT4受體在大腦的各個組織均有分布,其中以尾狀核和腦核分布居多,其通過多種方式參與大腦功能的調節,包括睡眠-覺醒周期、食欲、情緒和心理健康等[24]。在外周組織中,5-HT4受體主要分布于胃腸道、心臟和膀胱。隨著對5-HT4受體基因的克隆定位與表達的深入研究發現,5-HT4受體包括A、B、C、D、E、F、G、N、H九種亞型,其中5-HT4B受體的基因表達情況與5-HT4受體基本一致,是分布最廣、表達最多的一個亞型,可反映5-HT4受體的作用和功能。5-HT4受體與AC偶聯,刺激cAMP水平升高并激活蛋白激酶A的活動,使得K+通道關閉,促使膜的去極化,從而開放電壓敏感性Ca2+通道,使細胞內Ca2+水平升高,促進神經遞質的釋放增加。在中樞與胃腸神經系統中,5-HT4受體能夠促進乙酰膽堿的釋放,加強大腦的學習記憶以及胃腸道的動力與分泌;同時,5-HT4受體也可作用于心臟的心房肌細胞,一旦出現Ca2+超負荷,則會引起心律失常[25]。
3.5 5-HT5受體 5-HT5受體主要分布在中樞神經系統中,其有A與B兩種亞型,其中A亞型主要分布在人和動物的海馬、下丘腦、尾狀核中,而B亞型僅存在于動物的大腦皮質與嗅球中。5-HT5受體通過與G蛋白偶聯進行信息交換,從而起到抑制AC活性的作用,最終使cAMP合成減少,抑制興奮性突觸后電位[26]。5-HT5A及5-HT5B受體的藥理學特性與5-HT1D相似,具有負反饋抑制調節神經遞質釋放的作用。研究表明,5-HT5受體對麥角酸二乙酰胺表現出較高的親和力,并對5-HT5A受體起部分激動作用,通過研究5-HT5A受體的藥理作用發現,5-HT5A拮抗劑具有抗焦慮、抗抑郁和抗精神病的作用,為新藥研發提供了理論基礎[27]。
3.6 5-HT6受體 5-HT6受體主要分布在中樞神經系統,尤以紋狀體、邊緣系統等區域分布最多。研究證明,5-HT6受體與大腦的學習、記憶、情感等活動密切相關[28]。目前,對5-HT6受體的亞型報道較少。5-HT6受體主要分布在海馬CA1區神經元突觸后膜上,與學習、情感、記憶等活動關系密切,阻斷該受體可以提高海馬中的乙酰膽堿水平,從而改善認知障礙功能。研究證實,5-HT6受體拮抗劑與中樞神經系統神經遞質關系密切,參與調節乙酰膽堿、多巴胺、γ-氨基丁酸等神經遞質[29],在學習記憶障礙、焦慮、抑郁、肥胖等方面發揮重要作用,其作用機制可能為5-HT通過借助突觸后膜上的G蛋白偶聯受體與5-HT6受體結合,刺激AC的活性,增加胞內cAMP濃度激活蛋白激酶A,活化的蛋白激酶A進入核內,磷酸化cAMP反應元件結合蛋白,從而誘發下游靶基因的轉錄,影響機體的學習記憶功能[30]。
3.7 5-HT7受體 5-HT7受體在中樞神經系統主要位于下丘腦、腦干、中縫核等腦區,控制機體的體溫、睡眠和精神障礙等功能,其已被證明與減輕壓力、抑制抑郁和傷害感受有關[31]。在消化系統,5-HT7受體主要分布于腸肌間神經叢、黏膜下神經叢。近年研究發現,5-HT7受體在大鼠組織中包括A、B、C、E四種亞型,在人體組織中則包括A、B、D三種亞型,主要通過在碳末端長度和磷酸化位點來區別這些亞型[32]。5-HT7受體在腸道中表達,有研究發現腸道免疫細胞上5-HT7受體表達的證據,并證明5-HT7受體在炎癥性結腸炎的產生中具有關鍵作用[33]。另有研究發現,5-HT7受體可調控人和豚鼠的回腸,促使平滑肌松弛,其機制可能為5-HT7受體激活下丘腦前部神經元,并激活下行神經元,傳至肌肉效應器,使得平滑肌松弛[34]。抗精神病藥物阿立哌唑對5-HT7受體有較高的親和力,可通過聯合用藥來增強5-HT重攝取抑制劑類抗抑郁藥的治療效果[35]。
4 小 結
5-HT信號系統極其復雜,隨著對5-HT受體及其亞型分布和功能研究的深入,不同受體亞型在各個系統中的生理、病理及治療中的作用及意義不斷得以驗證。因此,了解各個受體及其亞型的結合位點有助于藥物的合理設計,提示5-HT在人體疾病的發生發展中扮演重要角色。但是5-HT信號系統與人體疾病的關系仍有待進一步探索,未來可以運用轉基因與基因篩查等新技術對5-HT各個受體及其亞型進行深入研究,以助于進一步認識疾病的發病機制,并為臨床疾病的預防與治療以及新藥的研發提供理論依據。
