牦牛HDAC2基因克隆及其在睪丸中的表達
2021-07-01 08:07:40曲尼拉姆楊柳青袁鈺潔
華北農學報 2021年3期
關鍵詞:小鼠
殷 實,王 斌,曲尼拉姆,楊柳青,袁鈺潔,李 鍵,2,3
(1.西南民族大學 青藏高原動物遺傳資源保護與利用教育部重點實驗室,四川 成都 610041; 2.西南民族大學 青藏高原動物遺傳資源保護與利用四川省重點實驗室,四川 成都 610041; 3.西南民族大學 畜牧獸醫學院,四川 成都 610041;4.西南民族大學 現代生物技術國家民委重點實驗室,四川 成都 610041)
組蛋白乙酰化是細胞內染色質修飾的一種重要方式,正常情況下細胞內的組蛋白乙酰化平衡由組蛋白去乙酰化酶(Histone deacetylases,HDACs)和組蛋白乙酰基轉移酶(Histone acetylases,HATs)共同維持,HDACs能夠移除組蛋白賴氨酸上的乙酰化基團,使組蛋白所帶的正電荷增加,進而促進其與帶負電的DNA分子間的結合變的緊密,使染色質由松弛轉變成凝集狀態并促使基因沉默[1-2],而HATs則執行相反的過程,由此可見,組蛋白的乙酰化調控對于細胞內基因轉錄調控以及染色質構象的改變等過程至關重要。
HDACs分為Ⅰ類 (HDAC1、HDAC2、HDAC8)、 Ⅱ類 (HDAC4-7、HDAC9、HDAC10)、 Ⅲ類 (Sirtuins) 及Ⅳ類 (HDAC10)[3]。Histone deacetylases 2 (HDAC2) 屬于Ⅰ類HDAC,已有文獻報道HDAC2參與了多個重要的生物學過程,例如HDAC2對小鼠的胚胎及神經系統發育至關重要。Hdac2突變的小鼠具有較高的胚胎致死率,存活下來的小鼠體質量、大腦質量及生育力與野生型小鼠相比均明顯下降[4-5],而過表達Hdac2則會損害小鼠突觸的可塑性及記憶的穩定性[6];HDAC2在腫瘤的發生過程中也扮演著重要角色,在大腸癌及胰腺癌等腫瘤細胞中敲除HDAC2會導致細胞生長停滯并導致細胞凋亡[7-8]。
精子發生的順利進行是維持雄性動物生殖能力的重要條件,在這一過程中大量基因的表達發生了變化,與此同時,染色體構象以及組蛋白的結構也發生了劇烈變化[9-10]。在小鼠等模式動物中的研究表明,精子發生的過程需要組蛋白去乙酰化的精確調控,例如在雄性小鼠腹腔內注射HDACs抑制劑苯丁酸鈉后圓形精細胞形態異常[11];生殖細胞缺失組蛋白去乙酰化酶SIRT1的小鼠無法繁育,其睪丸與正常小鼠相比明顯變小,減數分裂進程延遲,且出現大量形狀異常的精子[12]。牦牛(Bosgrunniens)是一種主要在我國青藏高原上活動的特有動物,是牧民重要的交通運輸工具,也是其重要的經濟來源[13]。然而野生牦牛的初次交配成功率不到40%[14],雄性牦牛直到4~5歲才發育至性成熟[15]。 因此,對牦牛的繁殖性能進行研究對于青藏高原人民生活水平的提高和經濟的發展具有推動作用。本試驗將通過基因克隆的方法獲得牦牛的HDAC2基因,對基因的保守性及其編碼蛋白的結構和功能進行分析,并檢測HDAC2的mRNA在牦牛睪丸中的表達及定位。本研究對后續研究HDAC2在牦牛精子發生中的作用提供了一定的基礎數據。
1 材料和方法
1.1 樣本收集
采樣地點為成都市青白江區屠宰場。依據齒齡將牦牛劃分為幼年組(0.5~1歲)、成年組(3~5歲)及老年組(7~9歲)。用無菌剪刀采集斷頸剖腹后的成年組牦牛肝臟、腎臟、肺、胃、脾臟、腦、心臟及卵巢組織,并采集各時間段牦牛睪丸組織(n=3)。分離下的組織用無菌剪刀剪成1.0 cm×1.0 cm×0.5 cm大小,立即轉移至-80 ℃液氮罐中保存。對于要進行原位雜交的組織,立即放于4%多聚甲醛(無RNA酶)中進行固定。
1.2 主要試劑及儀器
TRIzol(Invitrogen);PrimeScriptTMRT Reagent Kit反轉錄試劑盒(TaKaRa);Premix TapTMDNA聚合酶(TaKaRa);SYBR?Premix Ex TaqTMⅡ試劑盒(TaKaRa);pMD19-T載體(TaKaRa);DNA膠回收試劑盒(TIANGEN);DEPC H2O(TIANGEN);感受態細胞DH5α(TIANGEN);Real-time PCR Master (ROX)(Roche);CISH試劑盒(上海歌凡生物有限公司);紫外分光光度計(NanoDrop);熒光定量PCR儀(Applied Biosystems)。
1.3 總RNA的提取和反轉錄
各組織總RNA通過TRIzol進行提取,通過紫外分光光度計測定的260,280 nm 吸光值的比值在1.8~2.0的RNA樣品可進行后續試驗。反轉錄體系20 μL:RNA樣品1 μL,gDNA Purge 1 μL,NovoScript Plus All-in-one 1st Strand cDNA Synthesis SuperMix 10 μL,RNase Free Water 8 μL。反應條件:50 ℃孵育30 min,75 ℃孵育5 min,反轉錄產物置于4 ℃冰箱儲存。
1.4 基因克隆測序
依據參考GenBank中黃牛HDAC2基因序列(登錄號:NM_001075146.1)(表1)設計克隆引物,以牦牛肝臟cDNA為模板,通過普通PCR進行牦牛HDAC2基因的克隆。反應體系如下:2×LA Taq Master Mix 12.5 μL;上、下游引物及模板cDNA各1 μL;ddH2O 9.5 μL。反應條件:95 ℃ 4 min;95 ℃ 30 s,62 ℃ 45 s,72 ℃ 45 s,38個循環;72 ℃ 7 min。將產物與pMD19-T載體在16 ℃條件下連接15 h,之后與感受態細胞混合并在LB液體培養基中培養1 h,離心后將沉淀菌液均勻涂到含有0.1%氨芐的LB固體培養基上,37 ℃培養12 h,挑取白色單菌落接種于液態LB培養基中,搖床振蕩培養8 h,隨后選陽性菌株交由南京金斯瑞生物科技有限公司測序。
1.5 生物信息學分析
牦牛HDAC2基因及氨基酸序列的預測,以及同源性比對分析由DNAMAN 8軟件完成。通過MEGA軟件構建系統進化樹,通過ProtParam、ProtScale、NetPhos 3.1、PSIPRED、Pfam 以及SWISS MODE分別預測牦牛HDAC2蛋白的基本理化性質和結構。
1.6 熒光定量PCR(Real-time PCR)
牦牛各組織及不同時期睪丸的cDNA為模板,GAPDH為內參基因。PCR反應體系為15 μL:模板cDNA及上、下游引物各1 μL(10 μmol/L),SYBR Premix Ex TaqTM Ⅱ 7.5 μL,ddH2O 5.5 μL。反應條件:95 ℃ 預變性2 min;95 ℃ 變性10 s,60 ℃ 退火30 s,72 ℃延伸 30 s,其中變性、退火和延伸共持續40個循環。每個樣本重復3次。
1.7 反轉錄PCR (Reverse transcription PCR,RT-PCR)
以牦牛不同時期睪丸的cDNA為模板,GAPDH為內參基因。 反應體系:2×LA Taq Master Mix 12.5 μL;上、下游引物各1 μL(10 μmol/L);模板1 μL(800~1 000 ng/μL);ddH2O 9.5 μL。反應條件:95 ℃預變性4 min;95 ℃變性30 s,62 ℃退火45 s,72 ℃延伸45 s,72 ℃ 7 min。其中變性、退火及延伸共持續38個循環。每個樣品重復檢測3次,引物序列見表1。
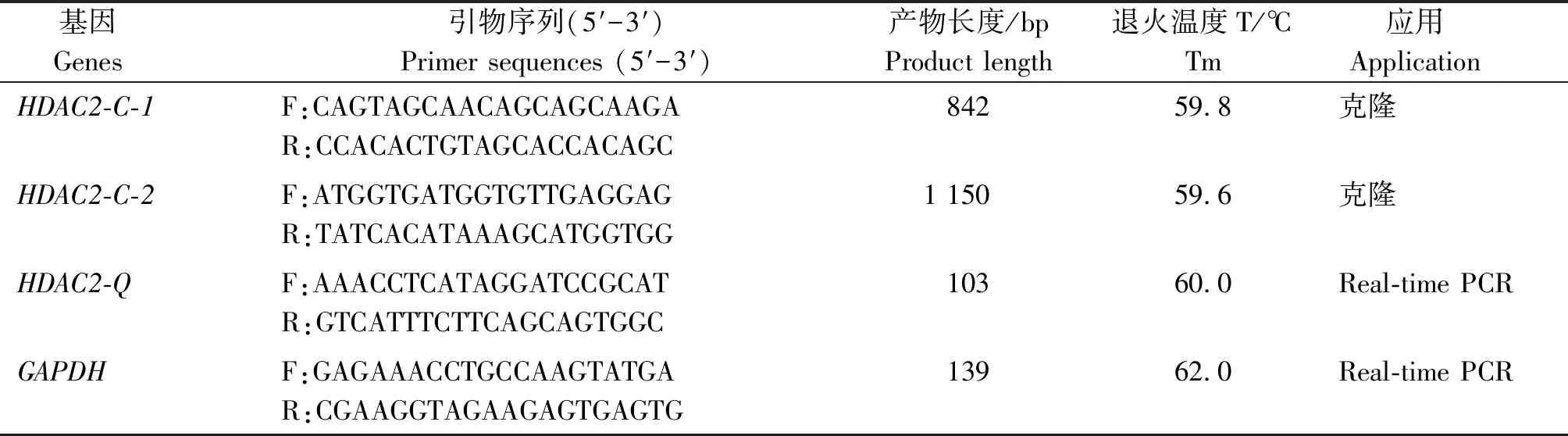
表1 PCR引物信息Tab.1 The information about PCR primers
1.8 色素原位雜交(Chromogenicin Situ Hybridization,CISH)
原位雜交的步驟參照CISH試劑盒說明書進行,主要步驟如下:石蠟包埋后的組織切片常規脫蠟至水洗,隨后用30% H2O2及純甲醇按照1∶9的比例配制混合液,室溫處理10 min,玻片置于濕盒內,滴加0.25%鹽酸,常溫孵育15 min;蛋白酶 K 覆蓋組織并于37 ℃處理 20 min,0.1 mol/L 甘氨酸洗1 min;4% PFA 固定10 min,分別用乙酸酐(pH值8.0)及5×SSC(Saline Sodium Citrate,檸檬酸鈉)(pH值7.5)清洗,預雜交液覆蓋組織并于65 ℃預雜交 1 h,用連接了地高辛的探針(500 ng/mL)覆蓋切片,65 ℃暗中雜交48 h,封閉液覆蓋切片,室溫反應30 min;滴加生物素地高辛抗體,37 ℃反應60 min;滴加 SABC(Strept Avidin-Biotin Complex,鏈霉親和素-生物素復合物),37 ℃反應20 min;滴加生物素化過氧化物酶,37 ℃反應20 min,DAB(3,3′-diaminobenzidine,二氨基聯苯胺)顯色,蘇木精染核。HDAC2的探針序列:5′-CACAAGCTATTCGTTTGTCTGATGCTCGAATAGAAATTCTCTTGT-3′,一個線蟲的非特異性序列探針作為陰性對照(Negative control,NC),其序列為5′-UUGUACUACACAAAAGUACUG-3′。
1.9 統計學分析
采用One-way ANOVA (Tukey′s 多重比較檢驗)分析數據的顯著性差異。結果以平均值標準誤形式表示,P>0.05為差異不顯著,P<0.05為差異顯著。
2 結果與分析
2.1 牦牛HDAC2基因的序列分析及蛋白結構預測
對克隆得到的HDAC2序列進行分析表明,牦牛HDAC2基因的ORF(Open reading frame,開放閱讀框)一共為1 467 bp,編碼488個氨基酸(圖1),序列的同源性分析結果表明,其與黃牛(Bostaurus)和野牛(Bisonbison)的基因及氨基酸序列相似度最高,均為100%,與馬(Equuscaballus)、野豬(Susscrofa)、白鰭豚(Lipotesvexillifer)、羊駝(Vicugnapacos)和人(Homosapiens)中HDAC2的序列相似度也均在90%以上 (表2、圖2)。

表2 牦牛與部分物種HDAC2核苷酸及氨基酸的序列同源性比對Tab.2 Alignment of HDAC2 nucleotide and amino acid sequences between yak and some species
氨基酸組成分析表明,HDAC2 蛋白由20種氨基酸構成,其中所占比例最高的為甘氨酸(Gly,9.0%)、賴氨酸(Lys,8.2%)、天冬氨酸(Asp,8.0%)及谷氨酸(Glu,8.0%)(表3)。在所有氨基酸中,帶正電荷的氨基酸(Arg+Lys)有63個,帶負電荷的氨基酸(Asp+Glu)有78個,由此可得,HDAC2蛋白整體上帶負電。HDAC2的最大疏水性指數為2.967,最小疏水性指數為-3.367,分別位于第161及454位氨基酸,其平均親水指數為-0.713(圖3),由此表明,HDAC2具有疏水脂溶性蛋白的特點。蛋白修飾位點分析表明,牦牛HDAC2 蛋白中一共含有50個磷酸化修飾位點,其中絲氨酸、酪氨酸和蘇氨酸磷酸化位點分別為21,14,15個(圖4)。牦牛HDAC2蛋白二級結構主要由α-螺旋、β-轉角及無規卷曲構成,其比例分別占38.73%,9.22%,52.05%,同時該蛋白不含有跨膜結構及信號肽(圖5-A)。保守結構域預測結果表明,HDAC2含有一個組蛋白去乙酰化酶結構域(圖5-B),HDAC2蛋白的三級結構預測的結果與二級結構相吻合(圖5-C)。
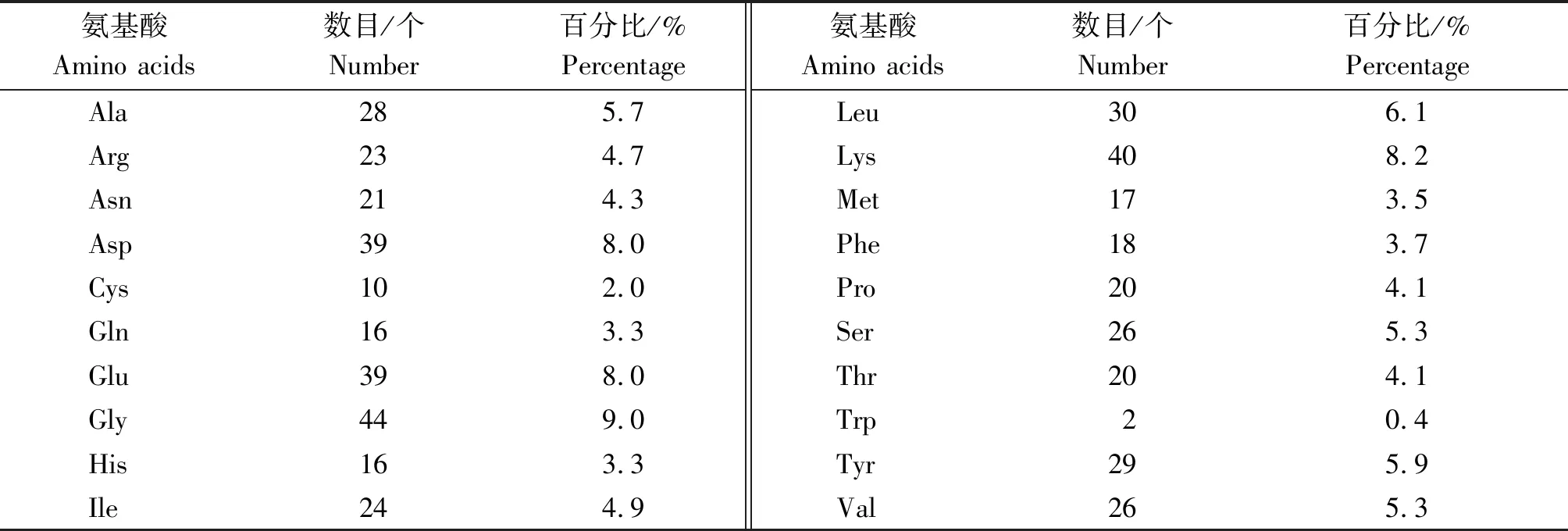
表3 牦牛HDAC2蛋白氨基酸組成Tab.3 The amino acids composition of yak HDAC2 protein
2.2 牦牛HDAC2 mRNA在各組織中的表達情況
實時熒光定量PCR的結果表明,在所檢測的9個牦牛組織中均發現有HDAC2mRNA的表達,其中在卵巢和睪丸中表達顯著高于其他組織,在腦、心臟和脾臟中的表達次之,在胃、肺、腎臟及肝中表達顯著低于其他組織(圖6)。
2.3 HDAC2在牦牛不同發育時期睪丸中的表達和定位
利用RT-PCR檢測了HDAC2在不同發育階段(幼年期、成年期及老年期)睪丸中的表達,結果表明,HDAC2在幼年期牦牛睪丸中的表達量最高,在成年期及老年期表達量較低(圖7)。原位雜交結果表明,HDAC2mRNA 在牦牛睪丸的非生殖類細胞支持細胞及睪丸間質細胞的胞質和胞核中均有所表達,在生殖類的精原細胞及精母細胞中也發現了HDAC2的表達。然而HDAC2不在精細胞中表達(圖8)。
3 討論與結論
HDAC2所屬的組蛋白去乙酰化酶家族是一類影響細胞內組蛋白乙酰化水平的重要酶類[2,16],已有報道HDAC2參與了人和小鼠等模式動物的胚胎發育、神經系統發育及腫瘤生成等生物學過程[4-8],然而有關牦牛HDAC2的信息知之甚少。本研究克隆了牦牛HDAC2的CDS序列,并分析了該序列的保守型及其編碼蛋白的結構,同時檢測了HDAC2的mRNA在牦牛睪丸中的表達及定位。
生物信息學的分析結果表明,牦牛HDAC2基因序列的保守性較高。HDAC2蛋白結構域的預測剛好證實了這一點。在牦牛HDAC2蛋白中發現了一個保守的組蛋白去乙酰化酶催化結構域,該結構域廣泛存在于Ⅰ型去乙酰化酶的不同成員中,通過與鋅離子結合催化賴氨酸進行去乙酰化[17-19]。此外,牦牛HDAC2的蛋白結構中含有多個潛在的磷酸化修飾位點,暗示該蛋白可能受到磷酸化調控。已有研究表明,HDAC2的磷酸化參與了多個重要的生物學過程,例如在大鼠中熱休克蛋白70(Heat Shock Protein 70,HSP70)能夠通過磷酸化HDAC2導致心肌肥大[20],在小鼠中HDAC2 能夠通過其第394位絲氨酸的磷酸化與叉頭轉錄因子O亞型3a(Forkhead box O3,FOXO3A)形成復合體,在氧化應激誘導的小鼠小腦顆粒神經元凋亡中發揮重要作用[21]。本研究預測的牦牛HDAC2蛋白結構、保守結構域及轉錄后修飾位點為后續進一步研究牦牛HDAC2的功能提供了一定的研究基礎。
在不同組織中的表達檢測表明,HDAC2的轉錄本在卵巢中的水平較高,提示HDAC2可能參與了牦牛卵巢發育的調控。研究表明,在小鼠的卵母細胞中敲除Hdac2會導致卵母細胞的著絲粒出現異常,導致其無法完成減數分裂并進而引起小鼠不孕等[22];在家貓卵母細胞成熟過程中HDAC2會從細胞核的核質向核仁轉移,這一過程對于GV期卵母細胞轉錄水平的沉默及卵子的成熟是必需的[23]。而HDAC2是否參與牦牛卵母細胞的成熟依然有待進一步研究。
HDAC2 的轉錄本在牦牛睪丸的除精細胞和精子之外的各類細胞中均有表達,這與小鼠睪丸中HDAC2的表達分布類似[24]。研究表明:精母細胞向精子轉變的過程中組蛋白需要被魚精蛋白所替代,這一過程需要核心組蛋白H3及H4 多個賴氨酸位點的高度乙酰化[25]。推測精細胞中缺少HDAC2很可能是為了保證組蛋白H3及H4 高度乙酰化的進行,進而保證組蛋白能夠被順利的替換。HDAC2蛋白在牦牛睪丸中的表達隨年齡的增長不斷降低,推測這可能與不同階段睪丸本身的細胞組成有關,幼年期的牦牛睪丸中以精原干細胞和精原細胞為主,缺少成熟的精細胞,而隨著個體年齡的增加,精細胞所占比例持續增加[26],因此,成年期中HDAC2mRNA水平的下調很可能是由于該階段睪丸中精細胞比例的增高所致;而在老年期牦牛睪丸中各類生精細胞的數量急劇下降[27],這可能是導致該階段HDAC2表達下降的一個重要原因。
牦牛HDAC2基因的開放閱讀框包含1 467個堿基,編碼488個氨基酸。牦牛HDAC2基因在哺乳動物中高度保守,預測的牦牛HDAC2蛋白是一個疏水脂溶性蛋白,包含一個組蛋白去乙酰化酶結構域和多個磷酸化位點,在幼年期睪丸組織中牦牛HDAC2mRNA的表達較高,其定位除精細胞之外的各類睪丸細胞中。本研究對于未來深入探討HDAC2對牦牛雄性生殖系統發育的調控機制提供了一定的參考數據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
