水稻MKKs基因在發育過程中對非生物脅迫處理的表達特征分析
2021-07-06 17:24:42余舜武張小慶陳守俊葉水烽羅利軍
上海農業學報 2021年3期
余舜武,張小慶,陳守俊,葉水烽,張 余,羅利軍
(上海市農業生物基因中心,上海201106)
植物的生長發育和逆境脅迫反應是在復雜的信號調控網絡下完成的,其中蛋白質磷酸化過程具有重要的信號傳遞功能。研究表明:細胞分裂原激活的蛋白激酶(MAPK,mitogen-activated protein kinase)級聯磷酸化過程在酵母、植物、動物等真核生物中普遍存在,參與細胞分裂、生長、激素信號傳遞及對外界環境的適應[1-2]。MAPK級聯途徑在真核生物中高度保守,一般存在3種類型的蛋白激酶:MAPKKKs(MAPKs激酶的激酶)、MKKs(MAPKs激酶)和MAPKs。這3種激酶從上到下逐級磷酸化,將信號放大并傳遞下去,最后通過MAPKs磷酸化不同底物,如轉錄因子、蛋白激酶、細胞骨架蛋白等,特異性調節下游基因的表達,使細胞、組織、器官及整個生物體作出相應反應[1-2]。
植物中的MAPK級聯途徑錯綜復雜,涉及發育和脅迫響應信號的傳導與整合。不同的發育或脅迫信號可能通過同一條激酶鏈傳遞,同一來源的信號又有可能通過不同的激酶鏈傳導,在錯綜復雜的信號網中MAPK級聯系統將信號巧妙有序地傳遞[1-3]。在三級級聯系統中,MKK家族基因位于中間,成員數最少,已知擬南芥、水稻和楊樹中只有8—10個成員,但第一級MAPKKK家族成員在擬南芥中多達80個,第三級MAPK家族也有20個成員[4]。在復雜的MAPK級聯中,MKKs在MAPK級聯各條途徑中起到了樞紐作用,能夠有效地協調上游MAPKKKs和下游MAPKs之間的信號傳遞。
MAPK家族基因最開始被發現與酵母細胞分裂和分化相關[5],該系統在植物中也是保守存在的,如AtMKK6基因參與擬南芥細胞分裂[6-7]。此外,MAPK也被發現參與植物激素調控的生長,如AtMKK7基因過量表達抑制生長素的極性運輸,導致擬南芥畸形發育[8]。還有報道認為水稻MKK4級聯系統調控谷粒大小[9]。但進一步研究認為,MAPKK家族基因與各種抗逆信號相關,在抗病性和非生物脅迫中扮演更重要角色。如油菜BnMKK1基因能響應多個非生物脅迫和激素處理,過量表達該基因改變了煙草株型,也改變了轉基因煙草的抗旱性和抗病能力[10];水稻OsMKK3和OsMK K10.2基因正調控植物的抗病性,其中OsMK K10.2基因還能改善水稻的抗旱性[11-12]。
全基因組范圍內研究MAPKK家族基因的表達模式已在擬南芥、棗、甜椒、西瓜和黃瓜等物種中開展,研究發現MAPKK家族基因與生長發育和逆境脅迫相關,水稻中也有MKKs基因的相關研究,但并不充分[3,13-16]。為全面了解水稻MKKs基因在水稻發育進程、激素和非生物逆境脅迫下的表達特點,本試驗利用不同發育時期不同器官組織的芯片表達數據和模擬非生物脅迫處理的定量PCR分析研究水稻中MKK家族基因在發育內部信號和環境外部逆境信號下的表達模式,以期為進一步解析水稻MKKs基因功能奠定基礎。
1 材料與方法
1.1 試驗材料和處理
試驗材料為水稻品種‘日本晴’,在正常的光照培養條件下培養至水稻幼苗三葉一心期,分別用PEG6000(聚乙二醇6000)、NaCl、冷、熱和植物生長調節劑ABA(脫落酸)處理這些材料。
PEG6000滲透脅迫是將水稻幼苗由標準水稻營養液移入含20%PEG6000的水稻營養液中;NaCl處理是將幼苗由標準營養液移入含150 mmol∕L NaCl的營養液中;熱處理是將植株放在42℃生長箱中生長;冷處理是將植株放入4℃生長箱中生長;ABA處理是用100μmol∕L ABA噴灑水稻葉片。在處理前(即0 h)、處理后3 h、6 h、12 h、24 h以及恢復正常培養后24 h分別剪取葉片,迅速放入液氮中保存以備后續試驗使用。
1.2 OsMKK蛋白序列分析
根據國家水稻數據中心(http:∕∕www.ricedata.cn∕gene∕index.htm)水稻基因數據,搜索TIGR Rice Genome Annotation(http:∕∕rice.plantbiology.msu.edu∕)數據庫并下載水稻OsMKK蛋白序列,從NCBI(https:∕∕www.ncbi.nlm.nih.gov∕gene∕)數據庫下載擬南芥MKK蛋白質序列,利用Clustal X對水稻和擬南芥MKK蛋白序列進行比對分析,然后利用MEGA 4軟件構建Neighbor-joining進化樹。
1.3 OsMKK基因芯片表達譜分析
從CREP(http:∕∕crep.ncpgr.cn)芯片表達數據庫中提取水稻OsMKK基因的表達數據,分別提取了‘珍汕97’‘明恢63’和‘汕優63’3個水稻品種27個發育時期和組織的表達信號值和三葉幼苗期經赤霉酸(GA3)、萘乙酸(NAA)和激動素(KT)等處理的表達信號值,具體數據見參考文獻[2]。每個樣品編號對應的發育時間和所取的組織見表1。
1.4 反轉錄定量PCR(qRT-PCR)分析
水稻葉片總RNA抽提采用Trizol法,具體使用方法參照中國TianGen公司TRNzol-A+Total RNA Reagent試劑盒說明書。以水稻總RNA為模板反轉錄合成無基因組DNA的第一鏈cDNA(北京全式金生物公司TransCript One-step gDNA Removal and cDNA Synthesis SuperMix試劑盒);然后以cDNA為模板,用定量PCR的方法進行表達量分析(TaKaRa SYBR?Premix Ex TaqTM試劑盒,PCR儀為ABI PRISM 7000 Real-Time PCR System)。用于定量PCR的MK K基因引物見表2。
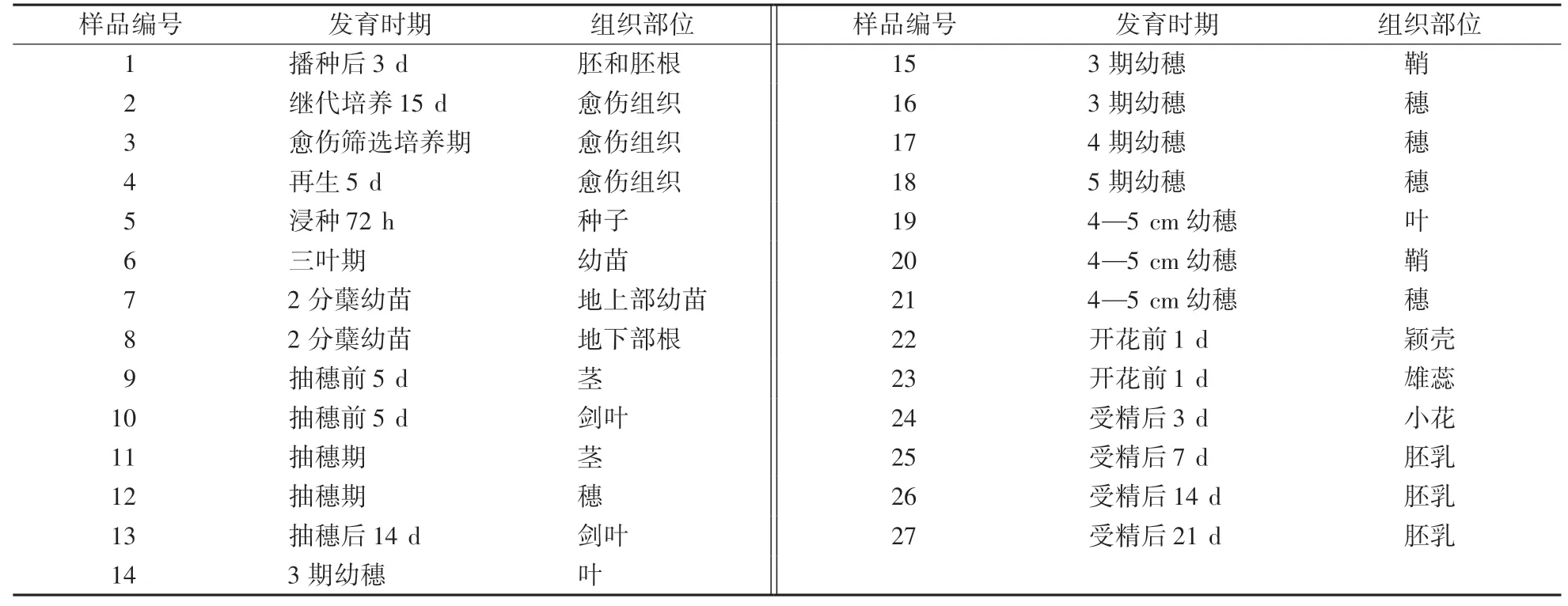
表1 水稻不同發育時期芯片表達譜分析的組織樣品Table 1 Tissue samples for microarray expression profile spectrum analysis of rice at different development stages
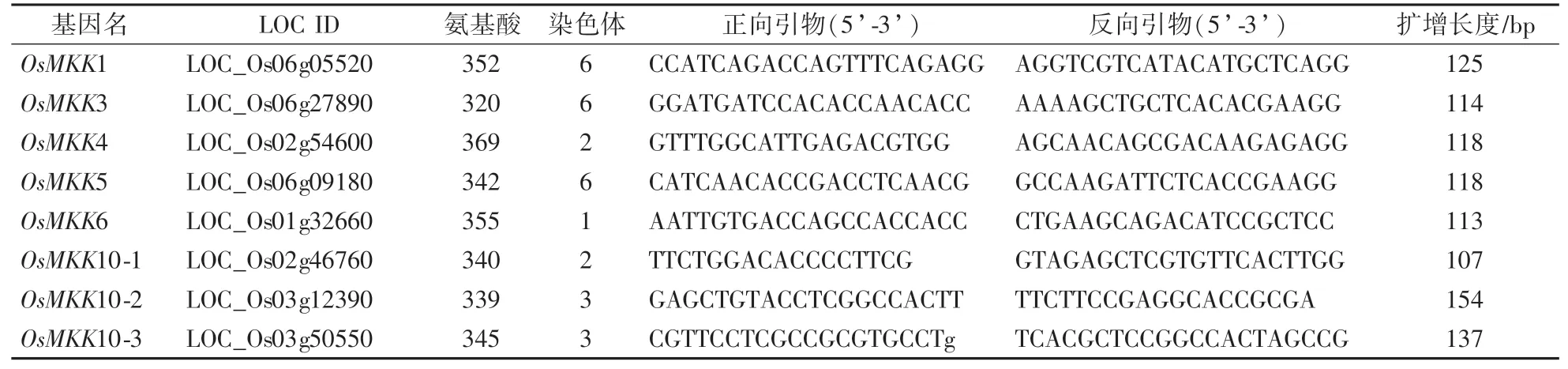
表2 水稻MKK家族基因成員和定量用PCR引物Table 2 Members of rice MKK gene family and primers used for qRT-PCR
2 結果與分析

圖1 水稻和擬南芥MKK蛋白家族的進化樹Fig.1 Phylogenetic tree of MKKs protein family in rice and Arabidopsis thaliana
2.1 OsMKK蛋白家族的進化樹
參照TIGR Rice Genome Annotation(http:∕∕rice.plantbiology.msu.edu∕)數據庫和國家水稻數據中心(http:∕∕www.ricedata.cn∕gene∕index.htm)相關數據,發現水稻中有8個OsMKK基因(表2),下載這些基因編碼的蛋白序列進行比對分析,發現OsMKK蛋白均具有在激活環中保守的天冬氨酸和賴氨酸殘基D(L∕I∕V)K和植物特異的磷酸化靶位點S∕TxxxxxS∕T。以擬南芥AtMKK蛋白序列作為參照,對OsMKK蛋白序列進行比對,構建Neighbor-joining進化樹。如圖1所示,OsMKK蛋白分為4個亞家族,其命名與擬南芥同源蛋白保持一致,A亞族包括OsMKK1和OsMKK6蛋白;C亞族包含OsMKK4和OsMKK5蛋白,但水稻中缺少AtMKK8—9蛋白;D亞族中與AtMKK10蛋白同源性最高的OsMKK10蛋白有3個;B亞族只有1個OsMKK3蛋白。
2.2 OsMKK基因在不同組織中的表達分析
CREP數據庫公開了Affymetrix芯片檢測數據,利用基因ID號搜索水稻OsMKK基因在‘珍汕97’‘明恢63’和‘汕優63’3個水稻品種27個不同發育時期組織器官的表達信號值。由于OsMKK10-3基因沒有對應芯片探針信號值,故OsMKK10-3基因未納入統計分析。結果表明:OsMK K基因表達模式在不同時期不同組織中表達存在差異,且品種間也存在差異(圖2)。OsMKK1基因在不同組織中均能檢測到表達,在葉片表達最高,幼穗中表達偏低,3個品種中‘珍汕97’表達最低。OsMKK4和OsMKK10-2基因表達與OsMK K1基因類似,葉片表達最高,幼穗中表達偏低。OsMKK3和OsMKK6基因表達模式聚在一起,OsMK K3基因整體表達量低,在幼穗中相對較高,胚乳中表達低,3個品種中‘珍汕97’表達相對偏高;OsMK K6基因整體表達量比OsMKK3基因偏高,其在愈傷組織、幼穗和胚乳中表達較高。OsMKK5與OsMK K10-1基因表達類似,雄蕊和幼穗中表達最低,但OsMKK10-1基因整體表達量很低,僅在胚乳中表達量高。表達模式聚類結果(圖2)與蛋白同源性聚類結果(圖1)不一致。

圖2 OsMKK家族基因表達模式的聚類分析Fig.2 Cluster analysis of OsMKK family genes expression patterns
2.3 OsMKK基因在激素處理后的表達分析
OsMKK基因對3種激素(GA3、NAA和KT)處理的表達數據值來源于CREP芯片數據。在不同激素處理下,OsMKK家族基因在3葉期的3個品種中表達量均呈現一定變化。以‘珍汕97’為例,OsMKK10-1、OsMK K3、OsMKK4和OsMK K6基因在激素處理后,除了個別處理稍有降低外,表達量均有一定提高(圖3)。OsMK K1、OsMKK5和OsMKK10-2基因表達量在激素處理后略有降低(圖3)。不同激素處理在品種間稍有變化,表明遺傳基礎的差異對激素反應存在一定影響。如GA3處理后,‘明恢63’的OsMKK5和OsMK K6基因表達量上升,而其他2個品種降低(圖3)。

圖3 NAA、GA3和KT處理后水稻MKK基因的表達譜Fig.3 Expression profiles of OsMKK genes treated with NAA,GA3 and KT
2.4 OsMKK基因在非生物逆境下的表達模式分析
對水稻幼苗模擬非生物逆境脅迫處理,取處理不同時間的葉片抽提總RNA,采用qRT-PCR的方法分析OsMKK家族基因的表達模式。如圖4所示,20%PEG6000處理能誘導OsMKK家族基因的表達量上升。在處理的24 h內,OsMKK1、OsMKK3和OsMK K5基因在12 h時表達量最高,OsMKK4基因在處理24 h時最高。150 mmol∕L NaCl處理也能誘導OsMKK家族基因表達量上升,其中OsMK K1、OsMKK3、OsMK K4和OsMKK5基因在處理的24 h內表達量持續升高,但NaCl處理對OsMKK10-2上調表達影響不顯著。低溫處理時,OsMK K1、OsMKK4和OsMK K6基因表達量上升。高溫能誘導OsMKK1和OsMKK3基因在早期表達量上升,9 h后逐漸下降恢復常量表達,OsMKK6和OsMK K10-2基因表達量呈下降趨勢,但不顯著。ABA處理能明顯誘導OsMKK1、OsMKK3和OsMKK5基因表達。綜上所述,在水稻MAPKK 4個亞家族中,A和C亞家族的OsMKK1、OsMKK6、OsMKK4和OsMK K5基因與非生物脅迫緊密相關,受干旱、鹽、高溫等脅迫和植物激素ABA誘導表達;B亞家族的OsMKK3基因與干旱、鹽脅迫和ABA處理相關;D亞家族的OsMK K10-2基因在PEG6000和冷處理下略有上升,其他應答不明顯,均與對照沒有顯著差異。以上結果說明,A∕B∕C亞家族的OsMKK基因均受非生物脅迫正調控。
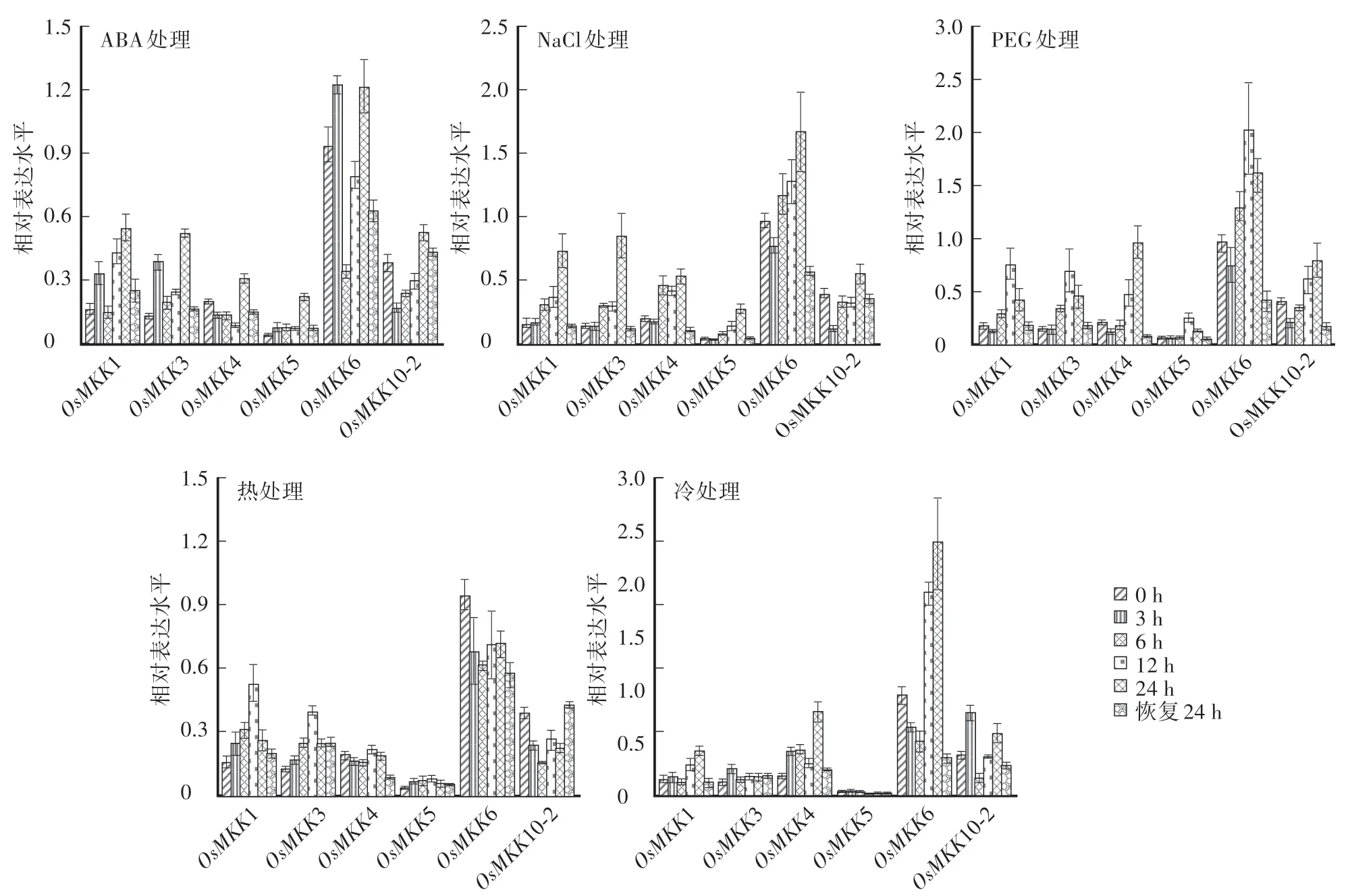
圖4 不同逆境處理下‘日本晴’幼苗OsMKK基因的表達量分析Fig.4 Analysis of OsMKK genes expression in‘Nipponbare’seedlings under different stress treatments
3 討論
MKK家族基因是植物生長和逆境反應中重要的信號傳導基因,其通過有限的幾個MKK成員將多個功能途徑共享到相同的MAPK級聯信號系統中。從擬南芥MK K成員的時空表達特異性和復合信號系統的形成來看,MKK所參與的MAPK級聯信號系統參與了植物的生長發育和防衛反應[17]。本研究中,從不同發育時期的不同組織器官中的表達來看,OsMKK家族基因在水稻生長發育中的不同時期均有表達,OsMK K1、OsMKK4和OsMK K10-2基因在營養器官中表達量偏高,而OsMKK3、OsMK K5和OsMKK10-1基因則在生殖器官中表達偏高,表明OsMKK家族基因在水稻生長發育的各個時期都扮演了重要的角色。OsMK K4基因所在的MAPK級聯系統能調控細胞分化和細胞增殖來影響穗的發育[8]。OsMKK4和OsMK K5基因同是水稻OsMKK家族C組成員,OsMKK5基因在‘明恢63’‘珍汕97’和‘汕優63’3個品種的苗期表達量相對較低,但在穗及雄蕊中表達較高,說明OsMKK5可能與OsMKK4基因相似,參與調控花器官的發育。本研究還發現,OsMKK基因對生長激素也存在一定的響應,這與擬南芥中的研究結果相似。擬南芥同源基因AtMKK3參與了植物激素GA3信號的調控,對擬南芥白天和黑夜的轉換起到重要作用[18]。此外,非生物脅迫處理也能誘導OsMKK4和OsMKK5基因表達明顯上調,表明OsMKK基因家族C組成員在水稻非生物逆境和生長發育中均具有重要功能。值得注意的是,OsMKK基因家族A和B組成員對非生物逆境脅迫有響應,它們同時也在水稻生長發育過程中的各個組織中有表達,這些結果進一步表明植物中的MAPK級聯控制信號系統錯綜復雜,涉及植物整個發育階段和脅迫響應信號的傳導和整合[1-3]。
本研究表明,OsMK K基因參與多種非生物脅迫,在滲透脅迫、鹽、高低溫等脅迫處理時表達量呈上升趨勢。OsMK K1基因的表達能夠被多種非生物逆境脅迫和ABA誘導,其同源基因AtMK K1∕2通過AtMEK K1-AtMKK1∕2-AtMPK4基因級聯途徑參與活性氧(ROS)平衡維持,使擬南芥在脅迫環境下正常生長,并能誘導與低濃度的活性氧有關的20個轉錄因子表達來應對環境脅迫[19]。除B亞家族只包含一個OsMK K基因成員外,其他亞家族都有2個以上成員,表明除B家族外的其他成員之間可能存在功能冗余。如A亞族中還存在一個OsMKK6基因成員,也參與鹽脅迫響應,在鹽處理下表達量上升。D亞家族的OsMK K10-2基因對非生物脅迫不敏感,這與水稻MKK家族其他的大部分基因不一樣,因為它們的表達能被多種非生物脅迫誘導表達,因此水稻MKK家族絕大多數基因都參與逆境脅迫反應。這表明水稻MK Ks基因能夠介導由不同非生物脅迫刺激的信號轉導,但基因功能是否存在冗余或者是每一個MK K基因鏈接的上下游是否一致,還需進一步研究。
MKKs基因參與的信號調控網絡是復雜的系統,除了參與多種非生物脅迫外,OsMK K基因也參與了生物脅迫響應。已有報道表明,水稻OsMKK3基因可激活下游MAPK和轉錄因子途徑來抵抗水稻白葉枯病和褐飛虱侵害[12,20]。更重要的發現是OsMKK10-2基因不僅能提高水稻的抗病性,同時能提高水稻的抗旱性[11]。以上結果表明:MKKs基因參與的級聯系統具有多功能性,處于中間的核心位置并扮演著核心作用,不僅調控植物的生長發育,而且還對植物在生物逆境和非生物逆境下的自我保護作用發揮重要作用,因此有必要充分挖掘和理清水稻MKKs基因的調控網絡。MKK家族基因在不同品種中的表達差異可能會影響到水稻的表型和農藝性狀,深入研究水稻種質資源中MKKs基因的功能差異,有助于解釋品種間農藝性狀的差異,為水稻的現代化育種提供理論參考。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
軍事文摘(2021年22期)2021-11-26 00:43:51
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
電子制作(2018年11期)2018-08-04 03:25:42
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
