苦蕎蛋白磷酸酶2C家族的鑒定及表達分析
2021-07-15 09:10:48劉耀東肖書雅王安虎李小意劉志斌李旭峰王健美
四川大學學報(自然科學版) 2021年4期
劉耀東, 肖書雅, 王安虎, 劉 宇, 方 陽, 李小意, 劉志斌, 李旭峰, 王健美, 楊 毅
(1.四川大學生命科學學院生物資源與生態環境教育部重點實驗室, 成都 610065; 2.西昌學院, 西昌 615000)
1 引 言
蛋白磷酸酶2C(protein phosphatases 2C, PP2C)依賴于Mg2+/Mn2+離子,是蛋白磷酸酶中極其重要的一支[1]. 從原核生物到真核生物,PP2C參與各種有意義的生命調控活動,在植物中,PP2C是蛋白磷酸酶最大的一個分支[2]. 植物PP2C被認為是蛋白信號級聯反應中的負調控因子,也在激素信號轉導過程中扮演重要角色[3]. 此外,植物PP2C還能調控植物的生長發育,包括種子萌發和根系發育[4-5]. 植物PP2C基因家族之前有很多研究,例如擬南芥(Arabidopsisthaliana)、水稻(Oryzasativa)和大豆(Glycinemax)[6-8]. 但是苦蕎PP2C基因家族的研究未見報道.
苦蕎(Fagopyrumtataricum)是一種藥食兼用的糧食,與甜蕎(Fagopyrumesculentum)相比,它更富含維生素B和蘆丁,這些成分有助于降低高血壓和緩解動脈硬化[9],并且苦蕎對惡劣環境有很強的耐受力,而PP2C的A亞族是ABA信號通路中的核心成員,參與植物逆境脅迫應答[10-12]. 苦蕎的基因組測序已經完成,加快了苦蕎抗逆的機制研究. 本研究參考擬南芥PP2C基因家族信息,通過生物信息學方法在苦蕎中全基因組中鑒定出81個PP2C成員,并分析了理化性質、基因結構、保守結構域、保守基序分布、染色體定位、進化情況等. 同時,通過qRT-PCR對苦蕎PP2C的A亞族基因組織特異性表達和ABA處理后受誘導情況進行了分析. 這些結果對苦蕎PP2C基因家族的后續研究奠定了基礎.
2 材料和方法
2.1 材 料
實驗使用的苦蕎種子為西蕎1號,由西昌學院王安虎老師提供. 所有的實驗材料在四川大學生命科學學院陽臺上的無菌土壤中進行培養,培養土壤為蛭石和土壤的混合物(1∶3, v/v). 在相同生長條件下選擇生長情況相同的樣品進行處理. 將收集到的植物樣品用液氮處理后保存在-80 ℃的冰箱中用以進一步實驗.
苦蕎的基因組完整信息下載自TBGP數據庫(http://www.mbkbase.org/Pinku1/). 作為參考的擬南芥PP2C家族成員從已發表文獻中獲取,蛋白質序列和CDS序列從Ensembl Plants數據庫(http://plants.ensembl.org/index.html)下載.
2.2 方 法
2.2.1 苦蕎中PP2C家族成員的鑒定 通過參考擬南芥PP2C家族成員的蛋白質序列和苦蕎其他基因家族的鑒定方法及參數選擇[7,13],使用BLAST軟件[14]搜索苦蕎中PP2C家族的候選序列(score≥100 and e-value≤10-10),并通過HMMER[15]軟件搜索PP2C結構域輔以搜索候選序列(e-value≤10-2). PP2C結構域的隱馬爾可夫模型文件(HMM)下載自PFAM數據庫(http://pfam.xfam.org/):PF00481. 候選序列通過搜索PFAM數據庫驗證是否含PP2C結構域以進一步篩選. 篩選后的序列中去掉基因的可變剪切,最終確認了81個苦蕎PP2C家族成員. 理論等電點(pI)和理論分子量(MW)等信息通過ExPasy網站(https://web.expasy.org/compute_pi/)上傳蛋白質序列進行分析.
2.2.2 基于系統發育分析對苦蕎PP2C家族進行分類 通過構建苦蕎和擬南芥PP2C的系統發育樹,并參考擬南芥PP2C家族的分類,根據它們在樹上的相對位置,將苦蕎PP2C分為不同的亞族. 其中,通過使用MUSCLE軟件(默認參數)進行序列比對,用RAxML-NG軟件[16](--all --msa input_file --model VT+I+G4+F --prefix at_ft --seed 678 --threads 4 --bs-tree autoMREs)構建極大似然(ML)系統發育樹. 建樹的模型通過Modeltest-NG軟件分析,選擇“VT+I+G4+F”模型. 系統發育樹的可視化通過iTOL網站(https://itol.embl.de/)完成.
2.2.3 苦蕎PP2C家族的基因結構分析和序列特征分析 苦蕎PP2C家族的基因結構分析通過GSDS2.0網站(http://gsds.gao-lab.org/)完成,蛋白質保守基序分析通過使用MEME工具(http://meme-suite.org/tools/meme)完成,MEME的參數調整為數目20,基序長度參數調整為6至200.
2.2.4 苦蕎PP2C家族的染色體定位和基因重復事件分析 苦蕎PP2C家族的染色體定位基于苦蕎基因組信息完成. 苦蕎PP2C家族的基因重復事件分析使用BLAST軟件進行序列比對,MCScanX軟件[17]進行線性分析(默認參數),得到苦蕎PP2C家族的串聯重復事件和區段重復事件.
2.2.5 實時熒光定量PCR 為了分析苦蕎PP2C的9個A亞族基因的組織特異性表達,通過qRT-PCR對它們在苦蕎根、莖、葉、花、果實五個器官中的表達進行分析,并進行三組生物學重復. 為了分析苦蕎PP2C的A亞族基因的受ABA誘導情況,用50 μmol/L ABA處理7日齡的苦蕎幼苗后通過qRT-PCR進行分析,其中,處理時長分別為0 h、0.5 h、2 h、4 h、8 h、12 h、24 h,用吸水紙干燥表面后使用RNA提取試劑盒(Tiangen)的方法從植物材料中提取總RNA,并通過使用Prime Script RT試劑盒(Takara)的方法合成cDNA. 在qRT-PCR實驗中,引物通過Primer3程序(http://frodo.wi.mit.edu/)獲取(表1),使用FtH3基因作為內部參考基因. 之后通過2-(ΔΔCt)方法分析獲得的mRNA的表達情況.
3 結 果
3.1 苦蕎81個PP2C家族成員的鑒定
用BLAST軟件[14]和HMMER軟件[15]篩選出600多個候選序列,通過是否含PP2C結構域進一步篩選出88個序列,在去除可變剪切的情況后,確認了81個苦蕎PP2C家族成員. 根據它們在染色體上的位置,從上往下依次命名為FtPP2C01至FtPP2C81,相關結果提供下載(https://github.com/felixlyd/ftpp2c),其中,苦蕎PP2C的A亞族序列特征信息如表2所示. 在苦蕎PP2C家族中,蛋白質序列長度從125(FtPP2C06)至1 098(FtPP2C15),平均長度為400;理論等電點(pI)分布在4.34(FtPP2C31)至9.6(FtPP2C60)之間;理論分子量在13 975.72 kD(FtPP2C06)至123 818.42 kD(FtPP2C15)之間.
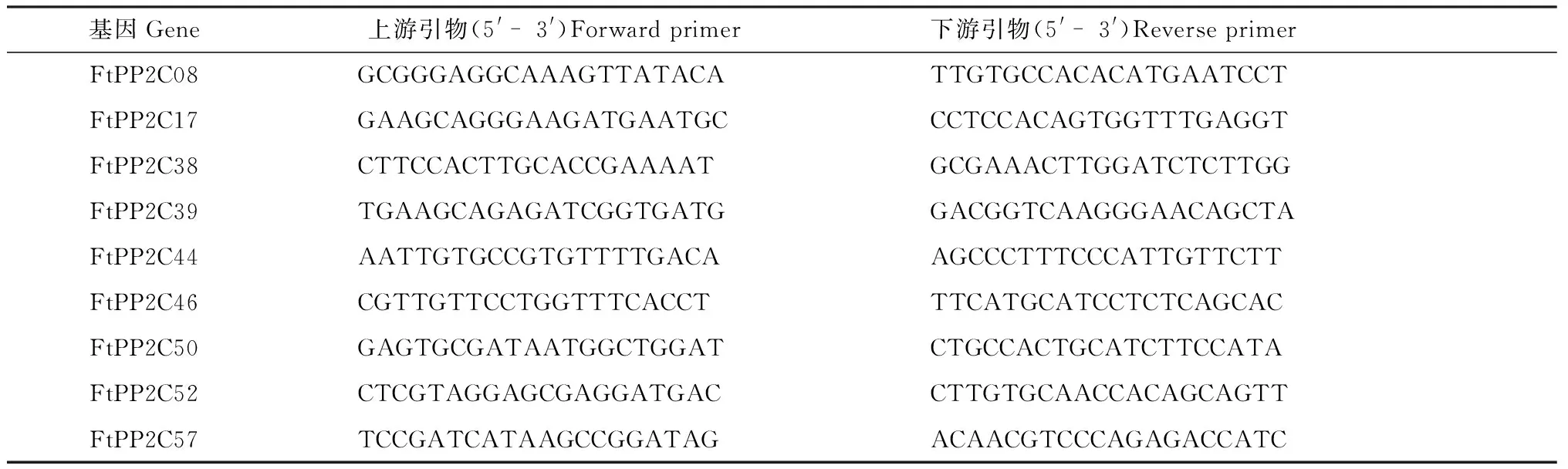
表1 苦蕎PP2C的A亞族基因表達分析的實時熒光定量引物

表2 苦蕎PP2C家族的A亞族基因的序列特征信息
3.2 苦蕎PP2C家族基因的分類
161個PP2C基因的系統發育樹通過ML方法構建(圖1),其中,苦蕎PP2C基因數目為81,擬南芥PP2C基因數目為80. 根據擬南芥PP2C基因的分類[6],將苦蕎PP2C基因分為從A至K共11個亞族,標識“other”表示那些未分類的PP2C基因,其中,苦蕎PP2C的A亞族有9個基因. Bootstrap值用來標識分類的可靠性. C亞族和D亞族的bootstrap值為99%,被分類為2個亞族;F亞族的兩個子亞族bootstrap值為92%,但這些基因被分類至F亞族. 這樣的分類方法是依據擬南芥PP2C的分類推導出的. 在進化樹中,最大的亞族是E亞族,有13個苦蕎PP2C基因和14個擬南芥PP2C基因;最小的亞族是J亞族,只有1個苦蕎PP2C基因和2個擬南芥PP2C基因. 此外,結果表明苦蕎和擬南芥PP2C基因在各亞族的數目分布上趨近相同.

表3 苦蕎PP2C的保守基序信息
3.3 苦蕎PP2C家族的基因結構和保守基序分析
基因結構分析主要為外顯子和內含子分布分析(圖 2b). 結果表明苦蕎PP2C家族基因中,內含子的數目最小為0(FtPP2C25),最大為19(FtPP2C15),但大多數分布在3至5之間,其中有27個苦蕎PP2C基因內含子數目為3,占比三分之一;外顯子的數目分布為1(FtPP2C25)至20(FtPP2C15),但大多數分布在4至6之間,其中有37個苦蕎PP2C基因外顯子數目為4,占比45.7%. 此外,在不同亞族間,外顯子和內含子的分布有一定差異,但在同亞族中,苦蕎PP2C基因之間的外顯子和內含子分布接近.
在苦蕎PP2C中找到了20個保守基序(motif)(圖 2a,表3). 除FtPP2C78、FtPP2C07、FtPP2C60、FtPP2C12、FtPP2C24外,其他苦蕎PP2C均含motif4. 除D亞族和K亞族外,其他亞族均含motif6. 苦蕎PP2C的A亞族的保守基序分布模式基本為motif14、motif4、motif18、motif1、motif7、motif6、motif2、motif20、motif10(除FtPP2C39和FtPP2C52不含motif14而FtPP2C44含額外的motif17). 其他亞族中的保守基序分布也極為接近,例如G亞族保守基序均為motif14、motif4、motif1、motif7、motif6、motif2、motif20、motif10;D亞族保守基序分布模式基本為motif 8、motif 4、motif 9、motif 1、motif 5、motif 15、motif 2、motif 3(FtPP2C45、FtPP2C48、FtPP2C41不含motif15;FtPP2C03不含motif8;FtPP2C54不含motif2和motif3).
基因結構分析結果和保守基序分析結果可驗證苦蕎PP2C家族分類的正確性,根據這些結果可以推測出苦蕎PP2C不同亞族間的基因功能有一定差異.

圖2 81個苦蕎PP2C基因的保守基序分布和基因結構分析 (a) 苦蕎PP2C的保守基序, (b)苦蕎PP2C基因的基因結構.Fig.2 The conserved protein motifs and gene structures of the 81FtPP2C genes (a)The conserved motifs of the FtPP2C proteins, (b)the gene structures of the FtPP2C genes.
3.4 苦蕎PP2C家族的染色體定位和基因重復事件分析
基于苦蕎基因組信息對苦蕎PP2C基因進行染色體定位,結果表明苦蕎PP2C基因在苦蕎的8個染色體上均有分布,并且分布情況相對均勻(圖 3a). 苦蕎PP2C基因在染色體上的分布數量分別為:15(Ft1)、10(Ft2)、9(Ft3)、9(Ft4)、4(Ft5)、8(Ft6)、10(Ft7)、16(Ft8). 基因重復事件分析結果表明苦蕎PP2C家族沒有串聯重復事件,但是有14次分布在苦蕎不同染色體上的區段重復事件(圖3b). 這表明區段重復事件是苦蕎PP2C基因數量增多的重要途徑.

圖3 苦蕎PP2C基因的染色體分布及線性分析
3.5 苦蕎PP2C的A亞族基因的組織特異性表達分析
通過qRT-PCR分析了苦蕎PP2C的A亞族基因在苦蕎根、莖、葉、花、果實五個器官中的表達模式,結果表明它們在所有器官中均有表達. 其中,除FtPP2C44外,8個基因在苦蕎生殖器官尤其是果實中,表達量比較高(圖4a),例如FtPP2C08主要在花和果實中表達,FtPP2C38在果實中的表達量是在葉中的1 715倍. 此外,FtPP2C39在根中表達量比較高,其他基因在根中表達量較低. 據此推測苦蕎PP2C家族的A亞族基因主要在苦蕎生殖器官中表達并發揮一定作用,這與之前擬南芥[6]、水稻[7]、大豆[8]中的研究一致.
3.6 苦蕎PP2C的A亞族基因受ABA誘導的表達情況
通過qRT-PCR分析了苦蕎PP2C的A亞族基因在ABA處理下的表達情況,其中,處理條件分別為0 h、0.5 h、2 h、4 h、8 h、12 h、24 h(圖 4b). 結果表明除FtPP2C08外,8個基因均受ABA誘導表達量顯著上調. 其中,在ABA誘導下4 h時,8個基因受ABA誘導表達量最高;FtPP2C39在所有處理條件下表達量都較高. 從而推測,苦蕎PP2C的A亞族基因在ABA處理后表達量上調具有一定生物學意義,而FtPP2C39發揮的作用可能較為重要.

圖4 苦蕎PP2C的A亞族基因的表達分析
4 討 論
過去的研究表明PP2C基因家族在植物中具有關鍵作用,但是目前有關苦蕎PP2C基因家族的研究很少. 本文從苦蕎全基因組中鑒定出81個PP2C家族成員,與擬南芥80個PP2C家族成員在數量上非常接近[7]. 對苦蕎和擬南芥PP2C進行系統發育分析,將苦蕎PP2C分類為從A-K的11個亞族,其中苦蕎PP2C的A亞族有9個成員. 結果還表明除AT4G11040外,擬南芥PP2C其他分類結果與以前的研究一致. 這可能是因為構建系統發育樹的方法不同,以前的研究[7]使用了鄰接法(NJ)而此處使用了極大似然法(ML). 此外,與擬南芥PP2C家族相比,苦蕎PP2C家族中C、D、K亞族的成員更多,而A、B、E、G、J亞族的成員更少. 基因重復事件分析結果表明苦蕎PP2C家族有14次區段重復事件,而擬南芥中則是13次,數值非常接近. 基因重復事件與基因家族的擴增有關,從而推測這可能是兩者PP2C數量上十分接近的原因. 出現這種情況的原因可能是植物PP2C的基因功能在進化過程中相對保守[18].
保守基序分析結果表明苦蕎PP2C家族在保守基序分布上有一定差異,但它們均含保守的PP2C結構域. 而且在相同亞族中,蛋白質保守基序的組成相近. 特定的保守基序分布可以代表特定的功能,例如擬南芥中PP2C的A亞族成員參與調控ABA應答過程[7],而它們的保守基序組成類似. 苦蕎PP2C的A亞族也具有相似的保守基序組成,與擬南芥PP2C的A亞族親緣性較高,從而推測苦蕎PP2C的A亞族也參與調控ABA應答過程. 而qRT-PCR分析結果表明苦蕎PP2C的A亞族基因在ABA處理后除FtPP2C08外均受誘導表達顯著上調,上調的原因可能是因為ABF結合PP2C啟動子調控其表達[19]. 因此推測苦蕎PP2C的A亞族基因在苦蕎ABA應答過程中發揮一定作用.
qRT-PCR分析結果還表明苦蕎PP2C的9個A亞族基因在根、莖、葉、花、果中均有一定表達,其中,FtPP2C08、FtPP2C17、FtPP2C38在花和果實中的表達量較高;FtPP2C39在根中表達量很高. 此外,FtPP2C39與擬南芥AT4G11040(又名DOG18)基因同源性較高,而擬南芥種子休眠和萌發受到DOG18的調控[20],據此推測FtPP2C39在苦蕎種子休眠和萌發過程中發揮一定作用.
綜上所述,本文通過生物信息學方法對苦蕎PP2C家族進行分析,并通過qRT-PCR對苦蕎PP2C的A亞族基因進行了組織特異性表達和ABA處理后受誘導情況分析,這些結果為進一步挖掘和研究苦蕎PP2C家族的基因功能奠定了基礎,有利于進一步對苦蕎抗逆的機制研究.
猜你喜歡
農產品加工(2023年14期)2023-08-17 10:32:28
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
青年歌聲(2018年5期)2018-10-29 03:18:40
山東工業技術(2016年15期)2016-12-01 05:31:22
廣東飼料(2016年3期)2016-12-01 03:43:12
家庭百事通·健康一點通(2016年4期)2016-04-22 09:16:57
林業與生態(2016年2期)2016-02-27 14:24:01
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
