2018—2020年福建省稻瘟病菌無毒基因型鑒定與分析
2021-07-16 02:11:12鄧云
福建農業學報 2021年5期
關鍵詞:水稻
鄧 云
(福建省南平市農業科學研究所,福建 南平 354200)
0 引言
【研究意義】由稻瘟病菌(Magnaporthe oryzae,無性世代為Pyricularia oryzae)引起的稻瘟病是水稻上最具毀滅性的真菌病害,也是福建省水稻生產中最嚴重的真菌性病害之一[1?6]。目前,全國因稻瘟病引起的水稻產量損失已經達到每年10億kg,給我國糧食安全帶來了嚴重隱患[7?8]。培育和種植抗病品種不僅能有效解決因農藥造成的殘毒和環境污染問題,同時能夠遏制稻瘟病流行,是防治稻瘟病最安全、經濟和有效的途徑。然而,稻瘟病菌生理小種的專化性、遺傳復雜性和頻繁變異,導致抗病水稻品種在連續推廣3~5年后抗性降低甚至喪失。【前人研究進展】優質抗原主要來源于稻瘟病多發區域內經長時間自然選擇留下的品種及野生稻和栽培稻等稻種資源中表現出抗稻瘟病的品種[6]。根據Flor[9]的基因對基因學說,水稻在抵抗稻瘟病菌侵染過程中,寄主的抗病基因與稻瘟病菌的無毒基因間編碼產物可互作識別,引發過敏性壞死反應(HR),抑制稻瘟病菌在植株中的擴展,從而產生抗病效應[10],李宏宇等[11]認為水稻與稻瘟病菌之間的特異互作符合“基因對基因”假說,即水稻品種有一種抗病基因R,稻瘟菌中就會有一個與之對應的無毒基因。福建省是稻瘟病多發地區,國內已有不少學者對福建省的稻瘟病菌小種組成、分布和消長動態、稻瘟病菌無毒基因在地區和年份間差異情況進報道[12?21]。【本研究切入點】隨著福建省主栽品種的變化,稻瘟病菌的無毒基因出現頻率也隨之改變,因此有必要分析稻瘟病菌近年來無毒基因年份間的動態變化情況。【擬解決的關鍵問題】本研究通過采集、分離2018—2020年福建省各主要稻區的稻瘟病生理小種,篩選出具有抗性頻率高、廣譜抗性的抗瘟基因,為今后根據不同地區流行菌株的無毒基因型和變化趨勢選擇培育具有相應廣譜抗病基因的水稻品種并進行合理布局,為提高水稻品種的抗性水平、延長抗性周期、防治稻瘟病奠定基礎[22?27]。
1 材料與方法
1.1 試驗材料
24個近等基因水稻(NILs)和對照高感品種麗江新團黑谷(LTH)由福建省農業科學院生物技術研究所田大剛轉贈。供試的稻瘟病菌單孢分別于2018—2020年采集自福建省南平、三明、龍巖等26個水稻種植地,每年9月中下旬(中稻黃熟期)隨機采集稻瘟病發生區域的穗頸部樣本,標明采集日期和地點,共計樣本179份(表1)。每年分離的稻瘟病菌單孢培養于濾紙片上并放?20 ℃冰箱保存備用。

表1 2018—2020年福建省稻瘟病菌供試菌株Table 1 M. oryzae sample collection, 2018-2020
1.2 試驗方法
致病性測定方法參照陳福如等[20]的方法以及稻瘟病苗期鑒定標準,病情調查方法參照辛威等[5]的方法。單個稻瘟病菌株的致病力(Pathogenic frequency,PF)、致病力類型劃分等參考楊秀娟等[21]的方法。致病力PF(%)=(感病水稻資源材料數量/所有供試水稻資源材料總數)×100。當PF≥ 70.0%時為強致病力,70.0%>PF≥50.0%時為較強致病力,50.0%>PF≥20.0%時為中等致病力,PF<20.0%時為弱致病力。
單個抗病基因的抗性頻率(Resistance frequency,RF)RF(%)=(對測試抗病基因無致病力的菌株數/所有菌株數)×100。
稻瘟病菌培養基為淀粉培養基,配方:可溶性淀粉10 g,酵母膏2 g,CaCo33 g,瓊脂粉16 g,蒸餾水1 000 mL,pH6.5。
1.3 統計分析
根據供試菌株對NILs的致病情況建立0-1數據庫,抗病記為1,感病記為0。用IBM SPSS Statistics19數據處理系統進行系統聚類分析,對供試菌株進行分類劃分并綜合衡量其遺傳差異大小,構建其遺傳關系樹狀,參考劉翔[1]的方法。
2 結果與分析
2.1 福建省稻瘟病菌株對已知抗病基因水稻品種的致病性
供試的179個稻瘟病菌株對24個含單個抗病基因水稻品種的致病性顯示,強致病力菌株占比26.81%,較強致病力菌株占比36.31%,中等致病力菌株占比27.37%,弱致病力菌株占比9.51%,較強致病力菌株占優勢,說明福建省的大多數稻瘟病菌株對供試24個抗病基因表現較強的致病力。
2.2 單個抗稻瘟病基因對福建省稻瘟病菌的抗性頻率
表2顯示,24個含單個抗病基因的近等基因系對福建省的稻瘟病菌群體的抗性頻率為19.32%~91.49%,其中Pi-z5和Pi-9(t)兩個抗病基因的抗性頻率最高,分別為91.49%和91.25%,Pi-kh和Pi-1兩個抗病基因的抗性頻率也較高,在80%~90%,Pi-7(t)、Pi-5(t)、Pi-k、Pi-12(t)等4個抗病基因的抗性頻率在70%~80%,Pi-ta和Pi-b的抗性頻率較低,低于20%。
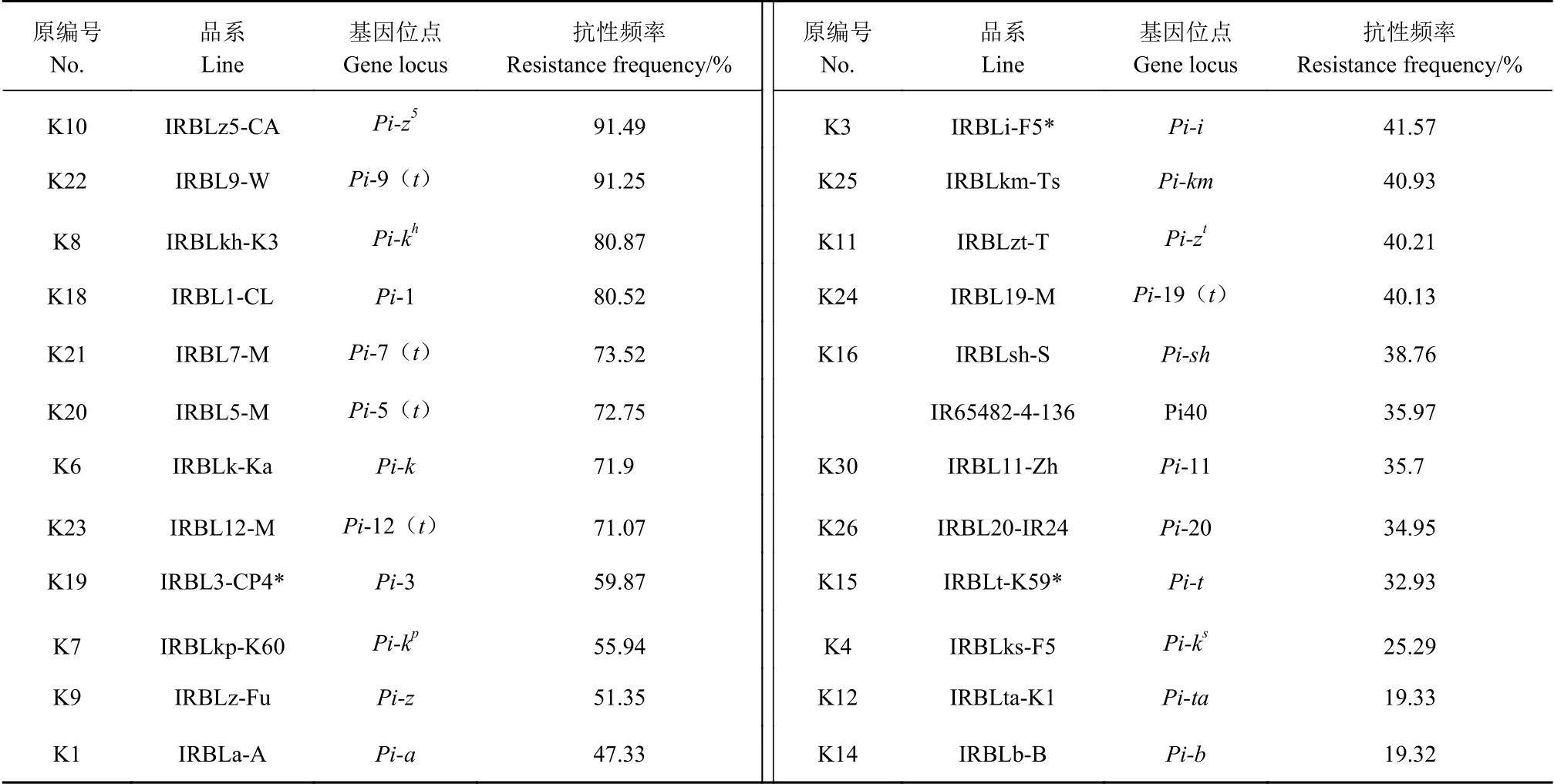
表2 抗稻瘟病基因對福建省稻瘟病菌的抗性頻率Table 2 Effect of blast resistant genes on frequency of resistance to M. oryzae isolated in Fujain
2.3 福建省稻瘟病菌無毒基因在2018—2020年間的分布
表3中24個福建稻瘟病菌的無毒基因在2018—2020年的分布變化情況可以看出,2018—2020年24個無毒基因出現的總的平均頻率逐年下降,由2018年的49.57%下降至2020年的38.01%,Avr-Z5,Avr-9(t)這2個無毒基因在3個年度中出現的頻率都在80%以上,平均出現頻率分別為88.95%和87.37%,年度間變化不大且穩定;Avr-1、Avr-kh、Avr-k、Avr-12(t)、Avr-7(t)、Avr-5(t)、Avr-3等7個無毒基因出現的平均頻率在50%~80%;無毒基因在2018—2020年出現頻率下降20%的有9個,分別為Avr-kh、Avr-7(t)、Avr-5(t)、Avr-3、Avr-kp、Avr-19(t)、Avr-km、Avr-z和Avr-ks,無毒基因在2018—2020年出現頻率上升20%有1個,Avr-20的頻率由16.67%上升至50.00%。平均出現頻率低于20%的3個無毒基 因為Avr-ks、Avr-b、Avr-ta,應用中應謹慎使用。
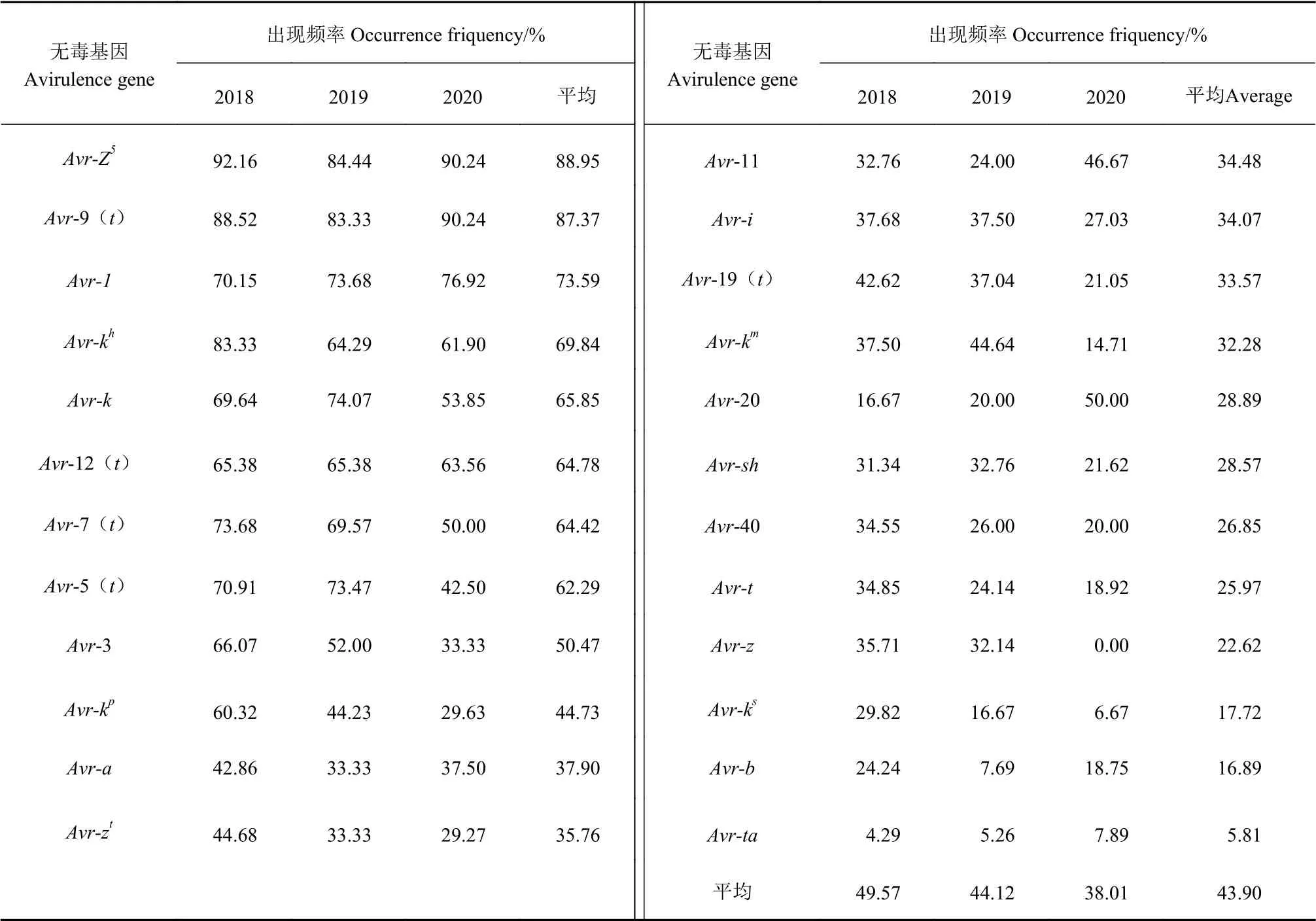
表3 福建省稻瘟病菌無毒基因在2018-2020年間的分布Table 3 Distribution of avirulent genes of M. oryzae in Fujian, 2018-2020
2.4 2018—2020年福建省稻瘟病菌的無毒基因的數量
表4的結果顯示,檢測的179個稻瘟病菌株分別含有0~22個數目不等的無毒基因,在數目上共出現了23種類型,含有6、7、8、9、11個無毒基因的病菌最多,在群體中的頻率分別為10.06%、8.94%、8.94%、7.82%和8.38%,其中含有6個無毒基因的頻率 最高。
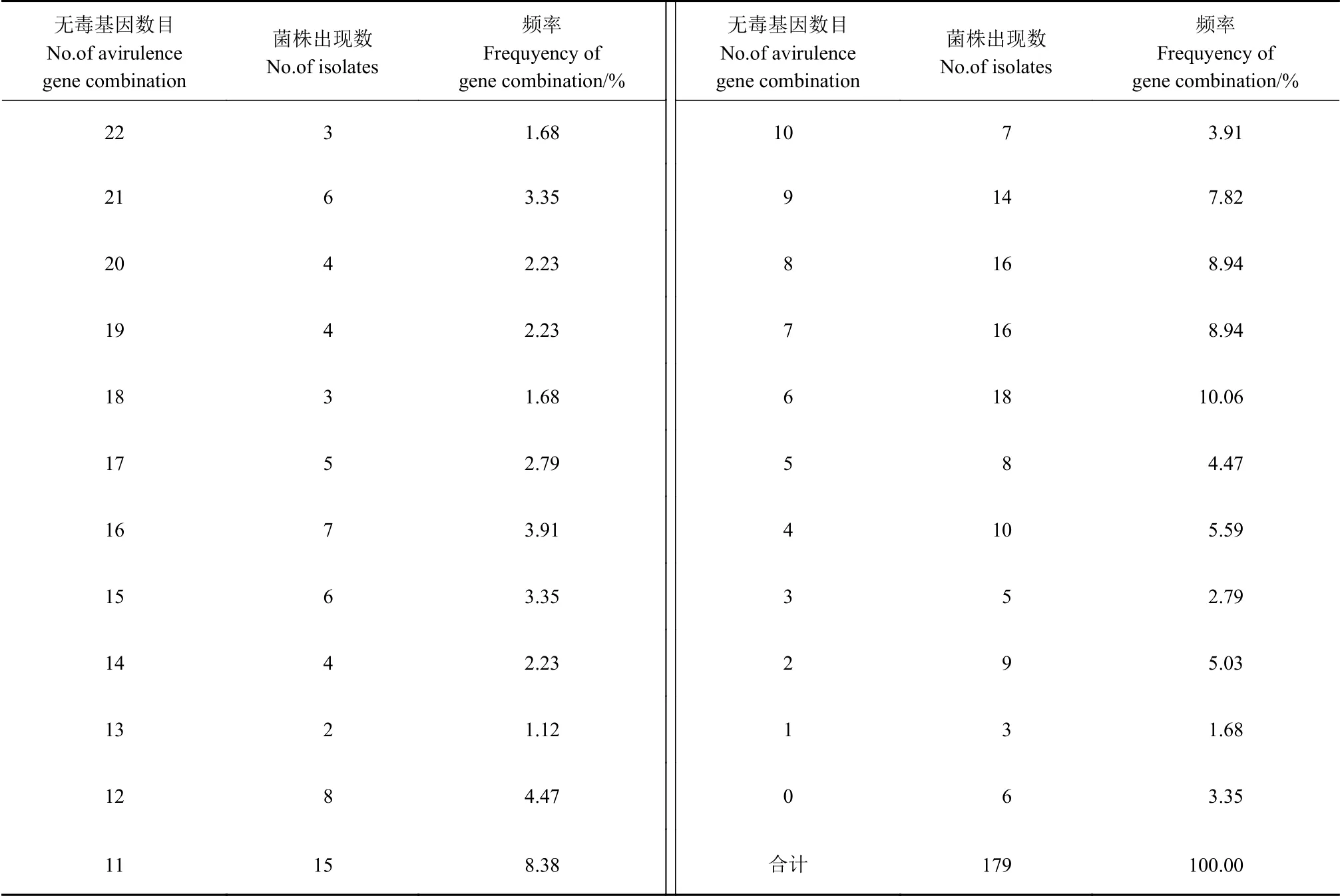
表4 2018—2020年福建省稻瘟病菌的不同數量無毒基因出現頻率Table 4 Occurrence frequency of avirulent gene combinations of M. oryzae in Fujian, 2018-2020
2.5 稻瘟病菌無毒基因在福建省9個地區的分布
表5的結果顯示,武夷山、松溪、邵武、浦城、光澤、建陽、寧化、將樂和上杭等9個地區都分布有24個無毒基因,無毒基因分布數量和出現頻率較高為上杭(49.19%),光澤次之(48.18 %),武夷山最低(41.23%)。在各地區中出現頻率達到90%以上的無毒基因有6個,分別為Avr-1、Avr-5、Avr-7、Avr-Z5和Avr-9、Avr-kh,其中Avr-1出現頻率最高的地區為寧化(94.74%),Avr-5出現頻率最高的地區為浦城(95.66%),Avr-Z5在武夷山、松溪、邵武、光澤、建陽和寧化等出現頻率都達到了90%以上,Avr-7出現頻率最高的地區為邵武(98.50%),Avr-9在武夷山、邵武和上杭的出現頻率都達到了90%以 上,Avr-kh出現頻率最高的地區為將樂(90.91%)。
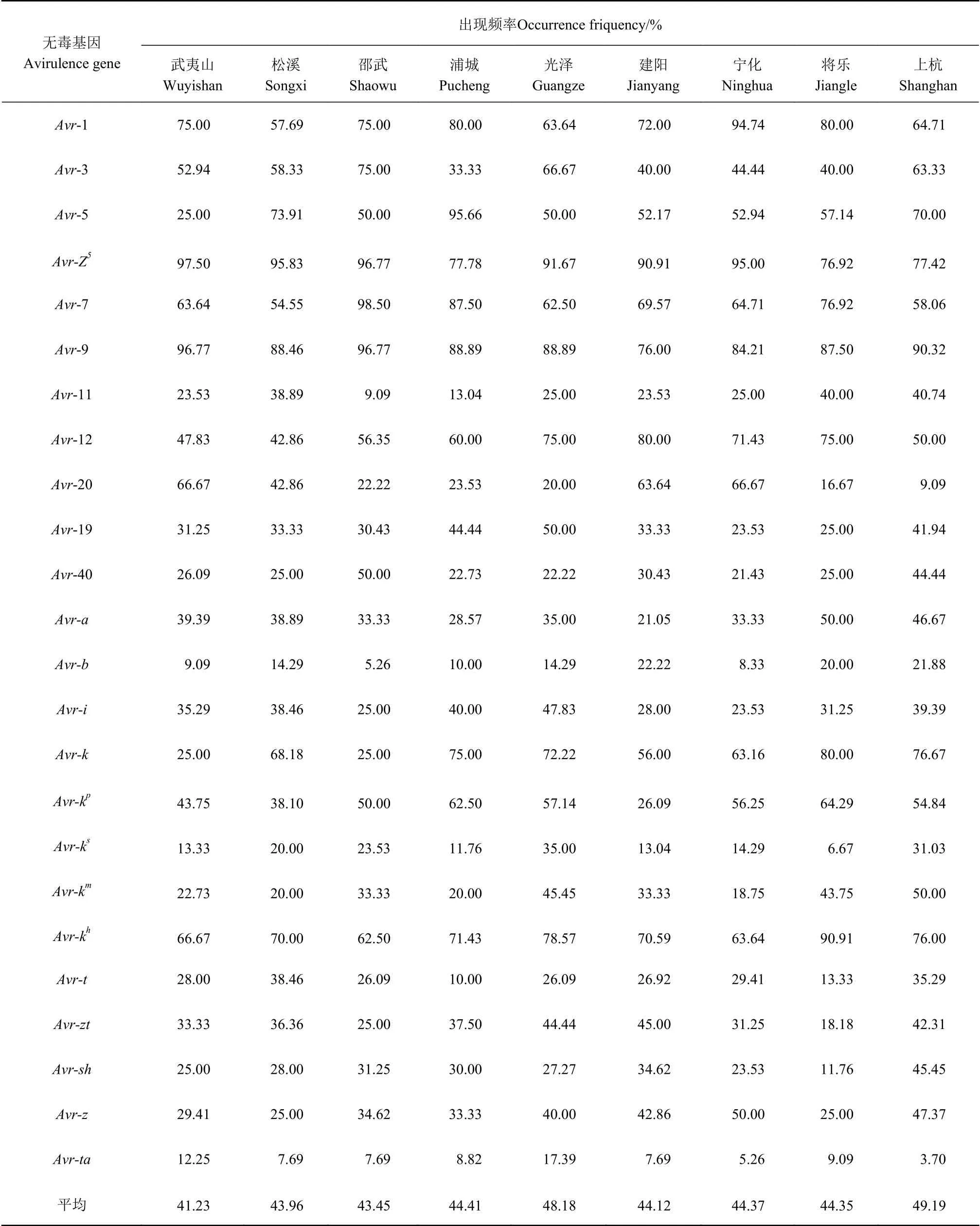
表5 稻瘟病菌無毒基因在福建省9個地區的分布Table 5 Distribution of M. oryzae avirulent genes in 9 regions in Fujian
2.6 179個菌株的抗性鑒定數據聚類分析
利用IBM SPSS Statistics19軟件聚類分析179個菌株對24個含單個抗病基因近等基因系的鑒定結果(圖1)表明,歐式距離(Euclidean distance)越小,菌株的遺傳距離越近,在歐式距離20遺傳相似水平處,不同抗病基因對不同菌株的抗感差異可以把179個菌株劃分為7個遺傳譜系(遺傳相似組)。在7個遺傳譜系中,第Ⅰ譜系為優勢譜系,含有124個菌株,占總數的69.27%;第Ⅱ譜系含有3個菌株;第Ⅲ譜系含有2個菌株,第Ⅳ譜系含有5個菌株,第Ⅴ譜系含有2個菌株,第Ⅵ譜系含有20個菌株,第Ⅶ譜系含有23個菌株。不同地區不同年份的菌株存在一定的遺傳相似性,但是大多數的遺傳相似程度并不與地區和年份的變化而呈現出很大的關聯性,例如歐式距離為1的相似菌株為4組,圖中由上至下依次為:第一組25號和75號菌株分別為2018年和2019年采集的光澤地區的稻瘟病菌單孢,第二組的57號和69號菌株分別為2018年采集的上杭地區的稻瘟病菌單孢和2018年松溪地區采集的稻瘟病菌單孢,第三組的2號和112號菌株分別為2018年采集的光澤地區的稻瘟病菌單孢和2019年建陽地區采集的稻瘟病菌單孢,第四組的84號和99號菌株都為2019年采集的將樂地區的稻瘟病菌單孢。從譜系的結果看稻瘟病菌的無毒基因在同一地區的種群豐富,在演變上隨機性較強且演變速度較快。
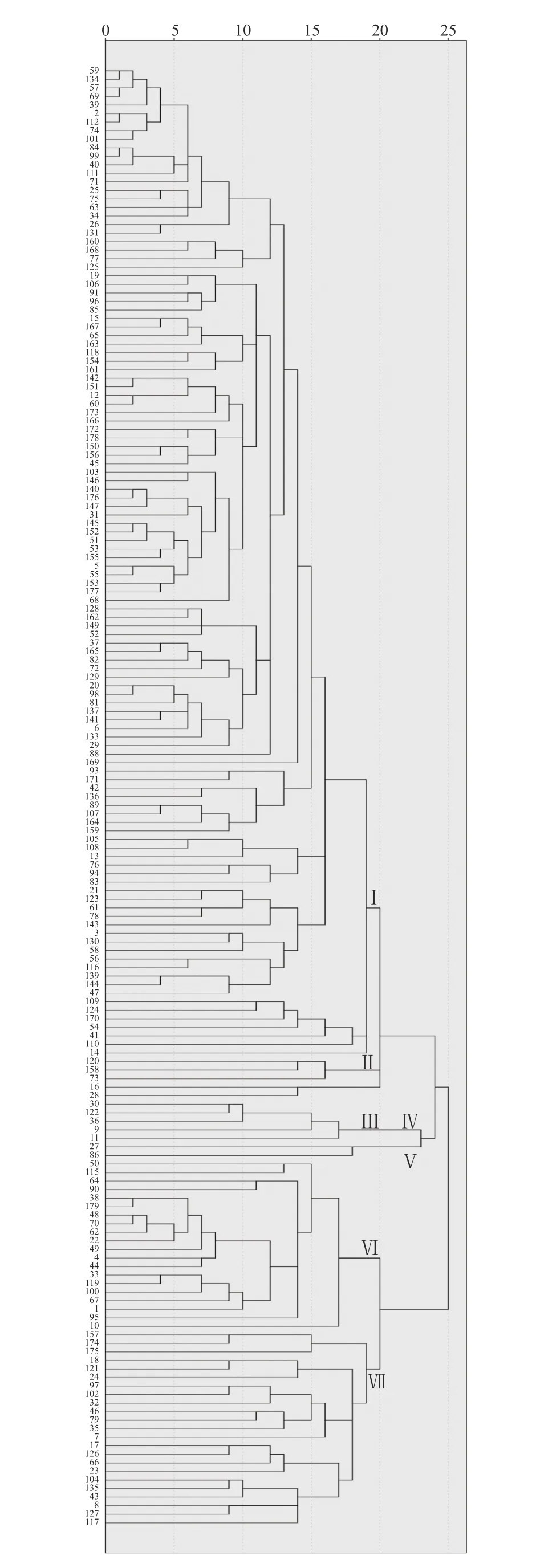
圖1 179個稻瘟病菌菌株對24個NILs親和反應型的聚類分析結果Fig. 1 Dendrogram on compatible response of 179 M. oryzae isolates to 24 NILs
3 討論與結論
目前國內外學者對于無毒基因與寄主之間的作用原理提出了以下幾點假說:Hammone-kosack K E等認為病原物侵染寄主時,無毒基因(Avr)編碼蛋白會被寄主細胞內對應的抗病基因產物識別,信號分子將局部性侵染反應迅速傳遞到整個植株,從而激活被侵入點細胞發生過敏性反應(hypersensitive response,HR)即防衛抗性機制,最終導致寄主細胞程序性死亡(PCD),病原物也因失去寄生繁殖生長受到抑制[27]從而獲得系統性抗性(systemic acquired resistance,SAR)。Mayank等[28]提 出 保 衛 假 說(guard hypothesis),該假說認為病原物的無毒基因的毒性因子Avr蛋白先與寄主體內相應的毒性靶向物接觸,并結合一起形成感受信號復合體,從而啟動防衛基因的表達激發植株產生抗性。Flor[9]提出“基因對基因”假說(gene-for-gene hypothesis),該假說認為病原的無毒基因編碼特異的激發子,引起細胞程序性死亡即過敏性反應;本試驗開展的理論基礎就是基于基因對基因假說以及稻瘟病菌與寄主水稻之間的互作效應。
本試驗于2018—2020年收集并分離福建省9個水稻主產區的中稻穗頸瘟單孢菌株179份,分別接菌鑒定24份含單個抗病基因的近等基因系。結果表明福建省較強致病力菌株占優勢。抗病基因的抗性頻率較高的有4個,其中Pi-z5和Pi-9(t)兩個抗病基因的抗性頻率最高,分別為91.49%和91.25%,Pi-kh和Pi-1兩個抗病基因的抗性頻率在80%-90%,Pi-7(t)、Pi-5(t)、Pi-k、Pi-12(t)等4個抗病基因的抗性頻率在70%-80%,這些抗病基因在育種中可以重點采用。出現頻率低于20%的3個無毒基因Avr-ks、Avr-b、Avr-ta應用中需注意觀察,謹慎使用。楊秀娟等[21]利用41個已知抗病基因水稻品種測定2003-2006年從福建省主要稻區采集分離的87個稻瘟病單胞菌株的致病性,結果表明:病菌群體對水稻抗病基因Pi-d2、Pi-k(1)、Pi-km、Pi-kh、Pi-1(1)、Pi-z5(1)、Pi-z5(2)、Pi-1(2)的毒力頻率均低于10%,提示這些抗病基因在福建省可作抗原使用;阮宏椿等[3]利用CO39近等基因系來鑒定2012-2015年間從福建省各稻區種植的普感品種麗江新團黑谷上采集的347株稻瘟病菌單孢菌株,結果表明:Pi-km、Pi-7(t)、Pi-9(t)、Pi-kp、Pi-k、Pi-kh、Pi-z5、Pi-ta等8個抗病基因的抗病性較強,抗譜均高于70%;本試驗的鑒定結果與前人成果大致相同,主要區別為本研究結果中的抗病基因Pi-ta和Pi-km的抗性頻率較前人研究結果下降幅度較大,應用時需謹慎,Pi-12(t)的抗性頻率有所上升,可以考慮采用,這也同時說明隨著福建省水稻主栽品種的布局變化,稻瘟病菌的無毒基因優勢基因型也隨之改變。因此,持續監測近年來的稻瘟病菌無毒基因型的組成和變化是抗病育種和抗病品種布局的關鍵依據。
Pi-z5和Pi-9(t)這兩個抗病基因是本研究結果中對福建省的稻瘟病菌抗性頻率最高的。Pi9基因定位于第6號染色體上,是一個廣譜的稻瘟病抗病基因。Qu等[29]研究發現Pi9的cDNA全長為4 009 bp,包括一段3 099 bp的編碼區和910 bp非編碼區,Pi9編碼一個羧基端含有富亮氨酸重復域(LRRs)的1 032個氨基酸組成的蛋白產物。在水稻中,Pi9基因是組成型表達,不受病原菌的誘導。Piz5基因也是定位在水稻第6號染色體上(又名Pi2),與Pi9基因復等位,也是一個廣譜的稻瘟病抗病基因。其最先被定位在標記RG64和RZ612之間[30?31]。而后被精細定位在分子標記RG64和AP22之間[32],Zhou等采用與克隆Pi9基因相同的方法克隆了Piz5基因,證明Piz5基因也屬于NBS-LRR類基因,與Pi9相同也含有2個內含子,其cDNA全長3332 bp,Piz5編碼由1個核苷酸結合位點(NBS)和3個富亮氨酸重復區(LRR)組成的包含1032個氨基酸的蛋白產物[33?37]。
本研究菌株的采樣方式為每年福建省中稻生產中稻瘟病發生地隨機采樣,不同于前人研究中的從單一的感病品種上采樣,隨機采樣存在如遇某地某年稻瘟病不發生則缺少樣本的情況,存在連續性不強的缺點,但是隨機采樣的樣本更能反映生產中主栽品種上正在流行的菌株,筆者認為隨機采樣更具有代表性。另外,稻瘟病菌群體的聚類分析結果顯示,同一地區的稻瘟病菌的菌群豐富且具有多樣性,今后應擴大采樣群體,加大分離稻瘟病菌單孢的數量,擴大群體進行研究以便得到更加完善的結論。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
