脅迫萌發對青稞籽粒中β-葡聚糖和γ-氨基丁酸含量的影響
2021-07-28 06:46:50李婷玉陳正行周文菊涂兆鑫
中國糧油學報 2021年6期
李婷玉 杜 艷 陳正行 周文菊 涂兆鑫 李 娟
(糧食發酵工藝與技術國家工程實驗室;江南大學1,無錫 214122)(青海華實科技投資管理有限公司2,西寧 810016)(江蘇省生物活性制品加工工程技術研究中心;江南大學3,無錫 214122)
青稞營養豐富,蛋白質含量適中,且脂肪含量較低,并含有豐富的礦質元素、維生素、膳食纖維以及β-葡聚糖、γ-氨基丁酸(GABA)和α-生育三烯醇等多種生理功效成分,其生態特性和營養保健作用都是其它糧食作物無法比擬的。其中,青稞中的β-葡聚糖平均含量為5.25%,是目前世界上β-葡聚糖含量最高的麥類作物。β-葡聚糖具有調節血糖、降低膽固醇、提高免疫力等功能[1]。GABA具有降血壓、神經營養功能、抗癲癇病、生殖生理功能等多種生理功能[2]。
發芽可用于改善谷物的外觀、質地、風味、口感及其營養價值。然而,發芽通常導致谷物中β-葡聚糖的分解,β-葡聚糖被內源性的β-葡聚糖酶水解。金屬離子對谷物萌發過程中β-葡聚糖的含量和β-葡聚糖酶的活性有很大影響。鐘正升[3]研究發現Mg2+、Zn2+、K+、Na+有助于β-葡聚糖酶的生成及β-葡聚糖的分解; Cu2+會抑制β-葡聚糖酶活性及β-葡聚糖的降解。畢靜[4]的研究結果表明Cu2+和Fe2+對酶活有明顯的抑制作用。彭維[5]的研究表明Na+、K+、Mg2+、Mn2+、Fe3+、Ca2+對β-葡聚糖酶具有激活作用,而Cu2+對其起抑制作用。富硒發芽可以抑制發芽青稞中β-葡聚糖的降解[6]。萌發可誘導生物活性成分的變化,尤其可以積累GABA等營養物質。研究表明,當植物暴露于許多不利條件時,如缺氧、干旱、低溫、高溫、高鹽等,GABA的積累迅速增加,表明GABA在非生物脅迫中可能起主要作用[7]。低溫脅迫[8-11]是促進GABA積累的有效途徑。Wallace等[12]研究發現,低溫脅迫通過改變植物細胞結構,降低pH,激活谷氨酸脫羧酶(GAD)活力從而促進GABA生成,同時抑制GABA轉氨作用,使GABA得到積累,但低溫脅迫萌發對β-葡聚糖含量的影響研究較少。
本實驗以青稞籽粒為原料,在萌發過程中采用金屬離子、亞硒酸鈉和低溫脅迫處理的方式,研究不同脅迫方式下青稞籽粒中主要功能性成分β-葡聚糖和GABA含量的變化情況和規律,以期為功能性青稞產品的開發提供參考。
1 材料與方法
1.1 原料與試劑
青稞(瓦藍,2019年9月收獲);七水合硫酸鎂、硫酸鋅、五水合硫酸銅、六水合氯化亞鐵、十二水合氯化鐵、亞硒酸鈉、四氫呋喃、三乙胺、鹽酸、結晶乙酸鈉、四硼酸鈉、2-巰基乙醇、三氯乙酸(TCA)、冰醋酸、氫氧化鈉、無水乙醇,均為分析純;次氯酸鈉(有效氯6%~14%)、鄰苯二甲醛(OPA)、γ-氨基丁酸標準品(色譜級)、Megazyme β-葡聚糖混聯檢測試劑盒、甲醇(色譜純)、乙腈(色譜純)。
1.2 儀器設備
BSC-250型恒溫恒濕箱, GZX-9246 MBE型數顯鼓風干燥箱, 150型高速多功能粉碎機, LXJ-IIB型低速大容量多管離心機, UV-3200型紫外分光光度計, KQ-50E型超聲波清洗器, NexIon 350D型電感耦合等離子體質譜儀, Agilent 1260 series 型高效液相色譜儀, Agilent Hypersil ODS柱(4.0 mm×250 mm,5 μm)。
1.3 方法
1.3.1 發芽青稞的制備
選取飽滿的青稞籽粒,用體積分數為0.2%~1%次氯酸鈉溶液浸泡消毒30 min后,去離子水清洗3~4次,再用去離子水在25 ℃下浸泡10 h。隨后,在直9 cm 的培養皿底部鋪四層紗布,然后均勻鋪放200粒青稞籽粒,在上面蓋2 層濕紗布。在溫度為15 ℃、相對濕度95%、避光的條件下發芽培養,每隔6 h灑100 mL去離子水。分別于發芽24、48、72 h時取樣,用去離子水洗滌3次后,用80 ℃熱水滅酶處理10 min,40 ℃干燥12 h,粉碎(過50目篩)。
1.3.2 金屬離子脅迫萌發
發芽48 h,在培養過程中噴灑不同濃度(0.04、0.08、0.16 g/L)金屬離子溶液(Fe2+、Fe3+、Cu2+、Mg2+、Zn2+)[5]。其余具體脅迫萌發方法同1.3.1。
1.3.3 亞硒酸鈉脅迫萌發
發芽48 h,在培養時噴灑不同濃度(4、8、16 mg/L)的亞硒酸鈉的溶液[6]。其余具體脅迫萌發方法同1.3.1。
1.3.4 低溫脅迫萌發
將洗凈浸泡后的青稞籽粒分別在5 ℃和-20 ℃脅迫24、48、72 h取出并在室溫下回溫30 min后萌發24 h。其余具體脅迫萌發方法同1.3.1。
1.3.5 β-葡聚糖含量測定
按照NY/T 2006—2011《谷物及其制品中β-葡聚糖含量的測定》。
1.3.6 GABA含量測定
采用OPA柱前衍生反相高效液相色譜-紫外檢測法。
供試品溶液的制備:精確稱取1.0 g青稞粉,加入5%三氯乙酸適量,搖勻,并定容至25 mL,常溫超聲40 min,離心(4 000 r/min,10 min)。取上清液,過0.22 μm濾膜。
色譜條件:色譜柱:Agilent Hypersil ODS柱(4.0 mm×250 mm,5 μm)。流動相A(pH=7.2)為27.6 mmol/L 乙酸鈉-三乙胺-四氫呋喃(體積比為500∶0.11∶2.5),流動相B(pH=7.2)為80.9 mmol/L 乙酸鈉-甲醇-乙腈(體積比為1∶2∶2),流速為1.0 mL/min,柱溫為40 ℃。檢測波長338 nm。采用梯度洗脫,洗脫程序為:0 min,8% B;17 min,50% B;20.1 min,100% B;24.0 min,0% B。
衍生條件:用高效液相色譜系統自動衍生裝置,將200 μL 供試品溶液和400 μL衍生劑[13]反應2 min。
1.3.7 金屬離子殘留量
按照GB 5009.268—2016《食品中多元素的測定》。
1.3.8 數據分析
所有實驗均重復2次,結果以平均值±標準差表示。應用SPSS軟件進行方差分析,顯著性分析采用Ducan’s多重檢驗,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結果與討論
2.1 發芽對青稞β-葡聚糖和GABA含量的影響
由圖1可知,隨著脅迫萌發時間延長,青稞中β-葡聚糖含量呈現下降的趨勢。這可能是因為青稞在發芽時,胚上皮層細胞分泌了赤霉酸,刺激胚乳糊粉層合成β-葡聚糖酶,水解胚乳細胞的β-葡聚糖[14],從而使得β-葡聚糖含量減少。其中,β-葡聚糖含量在發芽48~72 h時降解速率最快。這可能是因為發芽早期,β-葡聚糖酶活性較小,β-葡聚糖降解較慢。隨著發芽時間延長,β-葡聚糖酶活性增加,進而加快了β-葡聚糖降解。在整個發芽過程中,青稞中GABA含量總體呈先增加后減少的趨勢。GABA在發芽前期(0~24 h)積累較慢,到了發芽中期(24~48 h)GABA積累最快,而到了發芽后期(48~72 h) GABA含量減少。這是因為青稞籽粒在發芽初期,蛋白酶未被充分激活,隨著發芽時間延長,蛋白質水解形成了谷氨酸,在谷氨酸脫羧酶的作用下使得谷氨酸轉化為GABA[15]。而GABA的生成與消耗是一個動態平衡的過程[16],隨著GABA的積累,反遏抑制了GAD酶的活性而促進了GABA轉氨酶的活性,使GABA轉化為琥珀酸半醛,從而導致GABA含量的下降[17]。
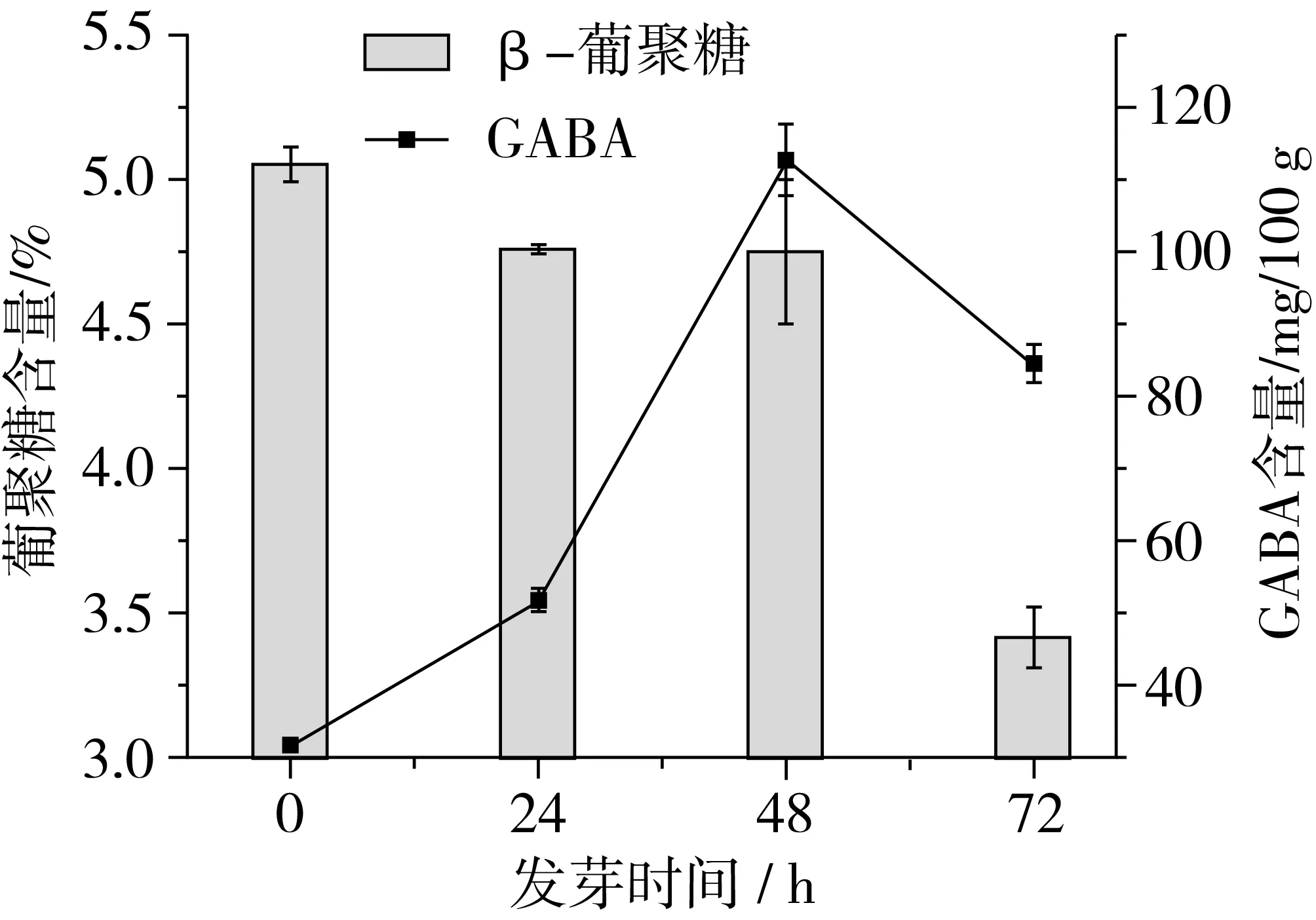
圖1 發芽對青稞β-葡聚糖和GABA含量的影響
2.2 金屬離子脅迫萌發對青稞β-葡聚糖和GABA含量的影響
由表1可知,采用5種金屬離子脅迫萌發48 h β-葡聚糖含量低于對照組,明顯可以促進β-葡聚糖降解,可能是因為Fe2+、Fe3+、Cu2+、Mg2+、Zn2+可以促進β-葡聚糖酶的活性[5-7]。而且采用5種金屬離子脅迫青稞萌發48 h 后GABA含量均低于對照組,表明Fe2+、Fe3+、Cu2+、Mg2+、Zn2+脅迫萌發抑制了青稞中GABA的積累。在5種金屬離子中,用Zn2+脅迫萌發48 h GABA含量增加量最少,最不利于GABA富集。關于Fe2+、Fe3+、Cu2+、Mg2+、Zn2+脅迫萌發對青稞籽粒 GABA富集效果的研究還鮮見報道,可能是因為金屬毒害導致青稞萌發能力喪失[17]。
用不同濃度的Fe2+、Fe3+、Cu2+、Mg2+、Zn2+溶液脅迫萌發青稞種子會造成金屬離子進入種子,形成富集,而這些離子攝入過多對人體造成一定的傷害。因此本實驗對經不同濃度(0.04、0.08、0.16 g/L)金屬離子溶液(Fe2+、Fe3+、Cu2+、Mg2+、Zn2+)脅迫48 h的發芽青稞中的Fe、Cu、Mg、Zn殘留量進行了檢測。經測定,用Fe2+、Fe3+、Cu2+、Mg2+、Zn2+脅迫發芽48 h的發芽青稞中各元素的殘留量如表2所示,Cu、Zn殘留量低于NY 861—2004 糧食(含谷物、豆類、薯類)及制品中鉛、鎘、鉻、汞、硒、砷、銅、鋅等8種元素限量中規定的上限(10.50 mg/kg)。
2.3 亞硒酸鈉脅迫萌發對青稞β-葡聚糖和GABA含量的影響
Marco等[19]研究發現,亞硒酸鈉會影響小麥籽粒的發育,減緩胚根的生長,一定濃度的亞硒酸鈉溶液會抑制植物籽粒的生長,進而減緩了β-葡聚糖的降解速率。由圖2可知,4 mg/L的亞硒酸鈉溶液脅迫下的發芽青稞中β-葡聚糖含量高于對照組,8、16 mg/L亞硒酸鈉溶液脅迫下的發芽青稞中β-葡聚糖含量明顯低于對照組,表明亞硒酸鈉溶液濃度較低時可以抑制發芽青稞中β-葡聚糖的降解,亞硒酸鈉溶液濃度較高時可以促進發芽青稞中β-葡聚糖的降解。4、8、16 mg/L亞硒酸鈉溶液作用下的發芽青稞GABA含量低于對照組,表明亞硒酸鈉不利于GABA積累。可能是由于一定濃度的亞硒酸鈉溶液會抑制植物籽粒的生長。
用不同濃度的亞硒酸鈉溶液脅迫萌發青稞種子,會造成硒進入種子,形成富集,而這些元素攝入過多對人體造成一定的傷害。因此本實驗對經不同濃度(4、8、16 mg/L)亞硒酸鈉溶液脅迫48 h的發芽青稞中的硒殘留量進行了檢測。經測定,用亞硒酸鈉脅迫發芽48 h的發芽青稞中硒的殘留量如表3所示,硒殘留量低于NY 861—2004 糧食(含谷物、豆類、薯類)及制品中鉛、鎘、鉻、汞、硒、砷、銅、鋅等8種元素限量中規定的上限(300 μg/kg)。
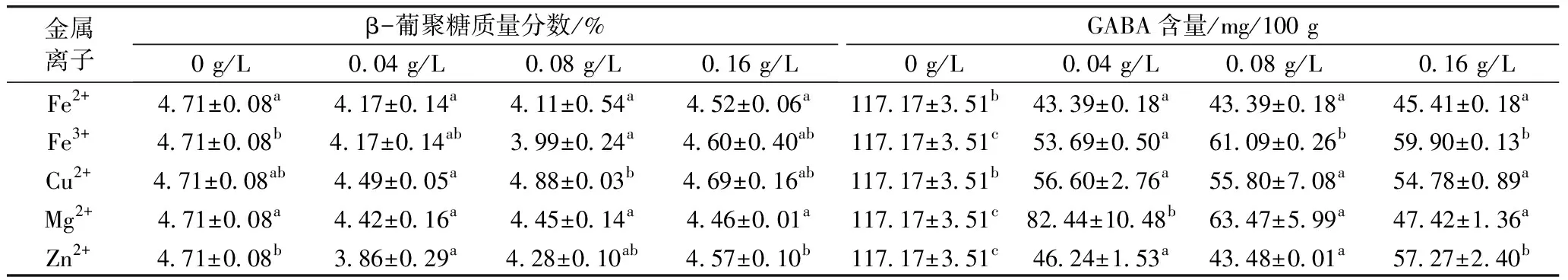
表1 金屬離子脅迫萌發對青稞中β-葡聚糖和GABA含量的影響

表2 發芽青稞中金屬的殘留量/mg/kg

表4 低溫脅迫萌發對青稞中β-葡聚糖和GABA含量的影響

圖2 亞硒酸鈉脅迫萌發對青稞β-葡聚糖和GABA含量的影響
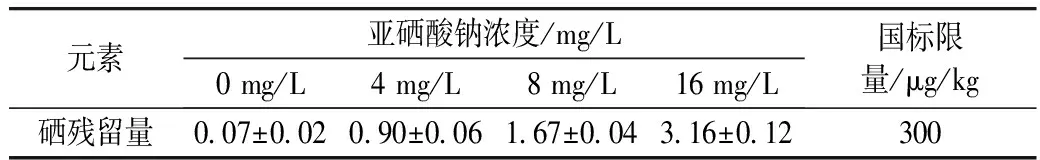
表3 發芽青稞中硒的殘留量/μg/kg
2.4 低溫脅迫對青稞β-葡聚糖和GABA含量的影響
由表4可知,-20 ℃脅迫條件下β-葡聚糖含量比5、15 ℃高,這可能是由于過低溫度破壞了青稞細胞結構導致的[8],青稞發芽率嚴重下降,抑制了青稞的發芽活力,鈍化了大部分β-葡聚糖酶,β-葡聚糖降解很少。5 ℃脅迫條件下β-葡聚糖含量比15 ℃高,這可能是較低的溫度抑制了β-葡聚糖酶的活性,β-葡聚糖降解越少。
-20 ℃脅迫條件下發芽青稞中GABA含量顯著高于15 ℃的原因是由于脅迫溫度過低形成了冰晶體,破壞了植物細胞內部結構,Ca2+、H+等滲入細胞質激活細胞質中鈣調素結合區的GAD活性,催化谷氨酸脫羧生成GABA[20],同時催化其轉氨降解的酶活性受到抑制,GABA轉氨作用減弱,使GABA得以大量積累。15 ℃發芽青稞中GABA含量顯著高于5 ℃的原因可能是由于脅迫溫度較高時,青稞對外界逆境環境反應不強,GAD激活不徹底,同時GABA轉氨作用增強,將GABA氧化降解形成琥珀酸,為TCA循環提供所需碳源,造成了GABA及其合成原料的消耗[21],使得發芽青稞中GABA含量有所下降。
3 結論
青稞籽粒在萌發過程中可以產生GABA,提高了GABA含量,但青稞中β-葡聚糖則逐漸被降解。低溫-20 ℃脅迫萌發,可以促進青稞中GABA的大量富集,且部分脅迫萌發方式(低溫-20 ℃)可以抑制β-葡聚糖的降解。此外,部分脅迫萌發方式(5種金屬離子、亞硒酸鈉、低溫5 ℃)卻促進了β-葡聚糖的降解。
