光合微生物膜鈣化建造的底棲鮞粒
——以遼西葫蘆島三道溝剖面張夏組鮞粒灘為例
2021-07-30 09:10:00柴凱旗秦仁月
東北石油大學學報 2021年3期
柴凱旗, 秦仁月
(中國地質大學(北京) 地球科學與資源學院,北京 100083 )
0 引言
鮞粒(ooid)一詞最早由KALKOWSKY E[1]提出,是指產出于淺水動蕩環境、由核心和圍繞核心的同心狀圈層組成的包覆碳酸鹽顆粒。對鮞粒的形成機制有多種假說,目前最為人們接受的假說為BREHM U等[2]的微生物成因假說和DUGUID S M A等[3]的化學成因假說。形成鮞粒皮層的最重要非晶質碳酸鈣(Amorphous Calcium Carbonate, ACC) 微薄層是研究鮞粒的關鍵。同時,這種非晶質碳酸鈣微薄層可能與微生物形成的微生物膜之間存在復雜聯系,因此,研究微生物膜復雜鈣化作用為解決鮞粒形成機理提供一個重要方向。
遼寧省葫蘆島三道溝剖面保留強迫型海退體系域中數十米厚的高能鮞粒灘灰巖地層,其中寒武系張夏組具有極好的巨型鮞粒。基于張夏組鮞粒的野外描述和正交偏光顯微鏡下對鮞粒完整微觀組構的觀察,以及巨型鮞粒的核心和皮層中存在大量絲狀或管狀鈣化藍細菌鞘化石,認為鮞粒形成于光合作用生物膜復雜的鈣化作用和細胞外聚合物質(Extracellular Polymeric Substances, EPS)的異養細菌降解誘導的碳酸鹽礦物原地沉淀作用[4],為了解光合作用微生物膜鈣化作用在動蕩水體環境中形成鮞粒機制提供一個新方向。
1 區域地質背景
葫蘆島大地構造位于華北地臺東北緣(見圖1),處于燕山構造帶東南部。中生代之前,研究區屬于穩定的華北克拉通,由變質的結晶基底和上覆穩定的沉積蓋層組成。蓋層主要以典型的淺海相碳酸鹽巖沉積為主。在太古代花崗巖基底上,由下到上分別沉積長城系、薊縣系和青白口系。與上覆寒武系到奧陶統呈平行不整合接觸,缺失鐵嶺組、洪水莊組和下馬嶺組等。同時,沉積層序的底界超覆明顯,體現典型的古陸邊緣特征[5]。受羅迪尼亞超大陸大規模裂解運動影響,新元古代,研究區海平面上升,沉積長城系、薊縣系和青白口系。古生代早期,華北地臺邊緣為被動大陸邊緣,從寒武紀第二世開始至中奧陶世,由于海平面上升,全球海侵開始,在遼東半島陸表海沉積環境形成一套陸源碎屑巖和碳酸鹽巖的混合沉積。

圖1 華北地臺寒武系苗嶺統張夏組沉積相帶(據文獻[6]修改)Fig.1 Depositional zonation of Zhangxia Formation of Miaolingian in Cambrian, North China Platform(modified by reference [6])
研究區在華北地臺寒武紀第二世晚期開始接受紅色泥巖和碳酸鹽巖混合沉積,逐漸向上過渡到大面積發育的鮞粒灘碳酸鹽巖的苗嶺統沉積(見圖2),再變化到以灰泥巖為主的芙蓉統碳酸鹽巖臺地沉積[6-8]。寒武紀,海平面開始上升,華北地臺開始呈現緩慢海侵趨勢,整體沉積以潮坪相和緩坡相的紅色泥巖和碳酸鹽巖為主的饅頭組。毛莊組與下伏饅頭組呈整合接觸。苗嶺統由毛莊組、徐莊組、張夏組和崮山組組成。研究區由饅頭組、毛莊組和徐莊組下部自下至上形成一套海侵混合潮坪沉積序列,變化到由徐莊組、張夏組和固山組上部形成一套鮞粒大面積分布的緩坡型臺地沉積序列[7-10]。其中,張夏組在三級強迫型海退體系域中發育多種微生物碳酸鹽巖生物丘。在沉積序列變化過程中,張夏組與上覆崮山組、下伏徐莊組呈淹沒不整合接觸,苗嶺統的張夏組構成一個完整的淹沒不整合三級層序[8-11](見圖2)。張夏組頂部發育一套數十米厚的巨型鮞粒灘,類似于PRATT B R等[12]描述的代表一種較高的碳酸鹽沉積速率和較慢的侵蝕速率的球粒主導的碳酸鹽巖臺地新類型[6-7]。張夏組沉積時期,沿古陸分布的沉積環境是以白云巖為主的潮坪相,環繞潮坪相是以碳酸鹽巖為主的緩坡相沉積環境。
2 鮞粒宏觀特征
三道溝剖面寒武系張夏組位于葫蘆島市新臺門鎮三道溝村西南約1.5 km的山上,上部為一套巨厚(約50 m)的、由鮞粒灘灰巖組成的強迫型海退體系域沉積,代表由緩坡相低能環境過渡為潮下、潮間高能環境,記錄一個海平面下降的沉積過程。在張夏組頂底面可識別兩個淹沒不整合面(見圖2)。巨型鮞粒(giant ooid)一般指粒徑大于2 mm的鮞粒類型,也被稱為豆粒(Pisoid)(見圖3(a-b))。高密度的巨型鮞粒集中發育于張夏組頂部的海退體系域,張夏組頂部的鮞粒灰巖夾有核形石灰巖(見圖2),代表海平面下降階段晚期記錄。鮞粒剖面呈淺灰色,鮞粒粒徑大多超過2 mm,形態多樣,呈規則的圓形、橢圓形及不規則長條狀(見圖3(c))。巨型鮞粒間多不相互接觸,其間充填淺灰色灰巖基質和三葉蟲等生物化石碎片,個別不規則顆粒表現為破碎的鮞粒碎片,也可見復合型鮞粒類型(見圖3(d))。鮞粒皮層的方解石呈同心紋層或呈放射狀排列。核心大多為生物骨架碎片和沉積巖碎屑,或以其他小型鮞粒為核心,個別核心不明顯或沒有核心(見圖3(c))。有些鮞粒邊緣可見底棲生物侵蝕而形成的凹坑,凹坑以一定的間隔均勻分布于鮞粒外圍。鮞粒灘灰巖往上過渡為鮞粒和核形石混合沉積單元(見圖3(d))。

圖2 華北地臺東北緣葫蘆島三道溝剖面Fig.2 Sandaogou Section at Huludao City on the northeast margin of North China Platform
3 鮞粒微觀特征
3.1 鮞粒灰巖結構
野外采集的三道溝剖面張夏組頂部鮞粒灰巖樣品制成的薄片,在正交偏光顯微鏡下表現為明顯的顆粒支撐結構,鮞粒粒徑多為毫米級別,大小不等,最小的只有100 μm,大者可達3 mm,屬于巨型鮞粒范疇。鮞粒形態不一,多為比較規則的圓形和橢圓形,也有心形、腎形、長條狀、短棒狀和紡錘狀,以及其他不規則的形態(見圖4(a)),鮞粒的外部形態大體反映內部核心形態。鮞粒核心主要由鮞粒、生物骨架、化石碎屑及暗色泥晶顆粒構成(見圖4(a-b)),充填在亮晶方解石構成的基質中(見圖4(a-i))。放射鮞粒中明顯可見圍繞核心呈放射狀生長一圈亮晶方解石結殼(見圖4(b)),在這層放射狀方解石的外圍發育數個明暗交替的同心圈層(見圖4(c-d)),構成同心—放射鮞粒。大多數鮞粒的核心為生物骨架化石碎屑(如三葉蟲化石)和巖石碎屑(見圖4(h)),或以暗色泥晶團塊為核心(見圖4(g)),少數鮞粒不發育核心或核心不明顯,如亮晶方解石充填的單晶鮞粒和暗色泥晶充填的泥晶鮞粒,二者為假鮞(見圖4(d))。其中,單晶鮞粒與GERDES G等[13]描述的Lanzarote組微生物席中的碳酸鹽顆粒類似,把有機質核心被透明纖維狀膠結物包裹的、且缺乏同心紋層的球狀顆粒稱為Corcoids。由于這種顆粒的纖維狀膠結物雜質很少,又有相對于泥晶方解石較高的結晶程度,所以在后成巖作用過程中,顆粒有較大的可能性先重結晶而形成大的方解石晶體顆粒。鮞粒的形成過程常受生物活動影響,如生物對鮞粒外部進行侵蝕,在鮞粒的內部形成泥晶化的凹坑,甚至生物活動形成的凹坑破壞鮞粒的核心,形成泥晶填充的假鮞(見圖4(e))。暗色泥晶中有微生物活動的痕跡,在成巖過程中的微生物痕跡被破壞殆盡,只有一些小黑點(見圖4(f)),可能是黃鐵礦晶體的殘余物質保存下來,間接證明生物存在過。暗色泥晶顆粒在高倍顯微鏡下表現為圍繞暗色有機質生長的放射方解石(見圖4(g)),類似CASTANIER S等[14]觀察到的富營養條件下的放射狀鮞粒前體。鮞粒灘上部發育以三葉蟲等生物化石碎片和鈣質生物膜碎片為核心的放射鮞粒(見圖4(h)),鮞粒灘中見復合型的鮞粒,即兩個及以上的鮞粒存在不連續的包裹關系或較小的鮞粒被包括在一個大的鮞粒之中(見圖4(i))。

圖3 葫蘆島三道溝剖面寒武系張夏組頂部鮞粒灰巖宏觀特征Fig.3 Macroscopic features of ooids at the top part of Zhangxia Formation of Cambrian at the Sandaogou Section, Huludao City

圖4 葫蘆島張夏組鮞粒灰巖微觀特征Fig.4 Microscopic features of the ooids in Zhangxia Formation at Huludao City
3.2 絲狀藍細菌主導的微生物膜
三道溝剖面發育的鮞粒基本特征是皮層中明暗交替紋層堆疊(見圖5(a-f)),多數鮞粒被暗色泥晶微生物薄膜或“泥晶套”包裹(見圖6(a、d))。在高倍顯微鏡下,鮞粒皮層內部可見密集發育的絲狀或管狀鈣化藍細菌鞘化石(見圖5(b-c)),鮞粒中發育大量的鈣化絲狀藍細菌化石(見圖5(a)),鮞粒邊緣存在比較集中的黑色小點,可能為黃鐵礦晶體的殘余物(見圖5(b)),放大鮞粒邊緣可以觀察到小黑點附近混雜相互纏繞的絲狀鈣化藍細菌鞘化石(見圖5(c))。鮞粒可見紋層狀的結構,核心明顯,鮞粒明暗交替紋層中順層排列絲狀鈣化藍細菌鞘化石(見圖5(d))。這些絲狀藍細菌大致平行于紋層排列,可相互交叉(見圖5(d-e))。鮞粒皮層中的絲狀鈣化藍細菌具有均勻的直徑(15~30 μm)和暗色泥晶壁(厚度約為2 μm),往往相互纏繞(見圖5(d-f))。鮞粒外部微生物薄膜中的絲狀藍細菌直徑較小(10~15 μm),鞘泥晶壁厚度約為2 μm,不相互纏繞也不成束產出(見圖6(d))。這種幾乎將整個鮞粒包裹起來的微生物薄膜內部發育高密度的絲狀藍細菌(見圖6(a-b)),微生物膜包裹形成的復核鮞粒在鮞粒中常見(見圖6(c、e))。微生物膜局部位置較厚,與生存環境和生物特性相關。大部分鮞粒受到成巖作用破壞,表現為重結晶或白云石化形成的粗大晶體(見圖6(e-f)),破壞原始紋層結構,但包裹鮞粒的微生物膜完好保存下來。鮞粒內部和鮞粒之間存在一些微生物膜或微生物席碎片,絲狀藍細菌在鮞粒內泥晶團塊構成的核心處和鮞粒間的泥晶凝塊中具有較高的密度(見圖7(a-b))。

圖5 葫蘆島張夏組鮞粒皮層中的鈣化藍細菌Fig.5 Cyanobacteria in ooids cortex of the Zhangxia Formation at Huludao City
鮞粒發育絲狀鈣化藍細菌在富有機質薄膜中密集分布可能代表黃鐵礦晶體殘余物的黑色小點(見圖7(c-d)),黃鐵礦晶體在鮞粒的暗色泥晶紋層中具有更高的密度。以暗色泥晶團塊為核心的鮞粒,核心處為絲狀鈣化藍細菌構成,可見密集的菱形白云石晶體,白云石化多晶鮞粒內部呈現不明顯的同心狀紋層或失去同心狀紋層的均一結構(見圖7(a-b)),充填在白云石化鮞粒之間的亮晶基質并未表現白云石化的跡象(見圖7(b))。微生物膜或微生物席是一種微生物群落構成的生態系統,在巖石成巖過程中屬于不易保存的部分,導致鮞粒的形成很難保存直接的生物證據。保留下來的生物膜碎片和生物膜作用的產物可作為生物活動的證據[8],如微生物膜包裹生物骨架碎片,骨架上殘留密集的絲狀鈣化藍細菌(見圖8(a-b))。同心鮞粒暗色泥晶紋層中,雖不見古代微生物遺體,但保留微生物膜作用的發育密集的黃鐵礦殘余物[10](見圖8(c-d))。

圖6 鮞粒外部微生物膜中的絲狀鈣化藍細菌Fig.6 Filamentous calcified cyanobacteria in microbialfilms around ooids
3.3 放射纖維狀方解石結殼
在葫蘆島三道溝剖面張夏組的鮞粒切片上,清晰可見由圍繞鮞粒邊緣發育的放射纖維狀方解石(Radial Fibrous Calcite, RFC)晶體和束狀光性方解石(Fascicular Optic Calcite, FOC)晶體形成的結殼(見圖9)。這些放射纖維狀方解石晶體長度可達相應鮞粒的半徑長度,可見明顯的雙晶紋,大多不對稱地覆蓋在鮞粒最外部圈層上和鮞粒間,從發育放射纖維狀方解石向不發育端逐漸過渡為非放射狀的邊緣膠結物。葫蘆島鮞粒灘中,RFC或FOC更多地發育于外部泥晶質薄膜或富有機質的鮞粒圈層,以及核心及其附近,代表其有機成因的線索。KENDALL A C等[15]將RFC解釋為沉積期后交代成因的產物;黃思靜等[16]研究RFC的層狀膠結物特征和層狀交界殘留的內部沉積物,認為RFC是原生沉積。與張夏組鮞粒間放射方解石構成的微凝塊一樣,放射方解石結殼可解釋為特別的富有機質環境中的生物成因產物[13],也可能是厚層微生物席發生復雜的鈣化作用及其微生物誘發的沉淀產物[9]。
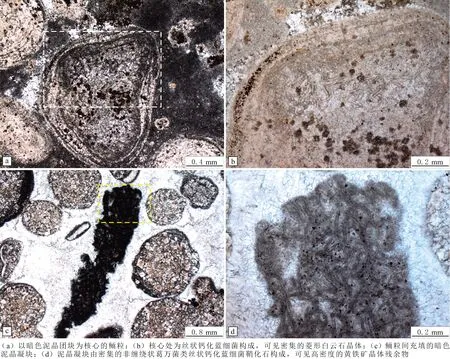
圖7 鮞粒內和外部的微生物膜或微生物席碎片Fig.7 Debris of microbialfilms or microbial mats in or around ooids
4 鮞粒形成機制
葫蘆島張夏組鮞粒的生長和底棲過程類似DAVIES P J等[17]提出的物理—化學模型和有機成因模型,主要包括三個重要階段——懸浮或擾動生長、短暫的棲息和沉睡。在整個生長周期中,前兩個階段只占不到5%的時間,大部分時間處于沉睡階段[18]。
第一階段,鮞粒開始最初的成核作用,圍繞核心發育代表無機沉淀作用[17]的放射狀方解石結殼,或類似放射鮞粒前體[14],放射狀方解石可能為有機質環境中生物成因的產物。由于微生物膜覆蓋,膜內藍細菌分泌的EPS上帶負電的官能團凝聚Mg2+和H+,造成“毒化作用”(poisoning effect)[17]而停止;隨后,微生物膜內的碳酸鹽礦物的原地沉淀產生泥晶和微亮晶。鮞粒皮層中發育的高密度相互纏繞可以歸為葛萬菌組合(GirvanellaGroup)[9-10,19],類似于齊永安等[20]描述的現代偽枝菌(Scytonema)。鮞粒外部微生物薄膜或泥晶套中的絲狀藍細菌可類比于現代微鞘菌(Microcoleus)[21]或鞘線菌(Coleofasciculus)[22]的絲狀或管狀鈣化藍細菌鞘化石(見圖5(e-f)、圖8(a-b))。這些絲狀藍細菌在鮞粒內部由泥晶團塊構成的核心處和鮞粒間的泥晶凝塊中具有較高的密度,泥晶凝塊和團塊可能是鈣化微生物膜的碎片(見圖7(c-d))。絲狀體的保存得益于藍細菌絲狀體分泌的黏液鞘的活體鈣化和微生物膜或微生物席的早期石化作用[10,23-24],藍細菌的活體鞘鈣化是一個直接與大氣圈二氧化碳濃縮作用機制(CCMs)相聯系的生理學機制[7,10,13]。
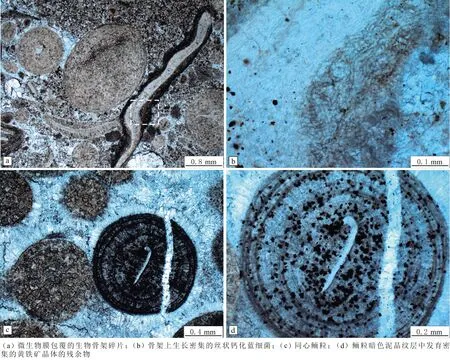
圖8 微生物膜內的黃鐵礦晶體Fig.8 Pyrite crystals in microbialmembrane
第二階段,張夏組鮞粒皮層,尤其是暗色泥晶紋層及鮞粒泥晶凝塊內常見黃鐵礦晶體的殘余物(見圖5(b-c)、圖7(d)、圖8(c-d)),表明異養細菌如硫酸鹽還原細菌(Sulfate-Reducing Bacteria, SRB)[14,25]造成的硫酸鹽還原反應產生HS-,并且升高SRB周圍微環境的堿度,從而誘發碳酸鹽礦物的原地沉淀和微生物席的早期石化而形成鮞粒紋層中的泥晶和微亮晶成分[10]。張文浩等[26]將超微的SRB和黃鐵礦晶體解釋為海水中缺氧條件下的產物。經過一段時間活動,微生物膜包覆的鮞粒核心在水體能量的作用下頻繁翻滾或躍移(如絲狀藍細菌平行于紋層分別表征的),形成具有一定體積的鮞粒和一定厚度的泥晶紋層而靜置下來。微生物席或微生物膜表現為趨光生長的特征,外在因素和內在因素共同驅動微生物席內“堿度發動機”[10,27]和微生物膜內鈣化藍細菌產生的有機物質(如細胞外聚合物質(EPS)),異養細菌降解(如硫酸鹽還原反應)并釋放Ca2+和Mg2+誘發碳酸鹽礦物的原地沉淀,從而重新活化鮞粒表面并進行無機沉淀[8-10,28]。
第三階段,鮞粒被埋藏在海底沉積物與海水界面處,并與富含可溶解有機質的海水頻繁接觸,鮞粒表面的有機質層成為新的下一個堆積圈層的沉淀基底。這層薄的高Mg/Ca的非晶質碳酸鈣(ACC)形成于鮞粒最外層的表面,作為一種含水有機大分子礦物相和生物礦化(或生物骨骼化)作用的過渡前體礦物相而存在[29]。非晶質碳酸鈣(ACC)很可能形成于藍細菌主導的生物膜外的細胞外聚合物。

圖9 葫蘆島張夏組鮞粒上的放射纖維狀和放射光性方解石結殼Fig.9 Fascicular fibrousand and fascicular optic calcite crystals on the cortex of ooids of Zhangxia Formation at Huludao City
5 討論
葫蘆島三道溝剖面張夏組同心—放射狀鮞粒,類似于澳大利亞鯊魚灣全新世至現代的鮞粒[30],以及巴哈馬[31-32]海灘的鮞粒特征;鮞粒灘上部發育以三葉蟲等生物化石碎片及鈣質生物膜碎片為核心的放射鮞粒(見圖4(h)),曾經被定義為努亞藻(Nuia Maslov)。HAN Z Z等[33]、LI Q等[34]、梅冥相等[6]曾描述過這樣的鮞粒。SPINCER B R[35]認為“努亞藻”是一種保留原始結構特征的假化石,應描述為“努亞藻狀的放射鮞粒”,“努亞藻狀鮞粒”的主導性體現寒武紀“方解石海”的基本特點[6]。鮞粒灘中可見復合型鮞粒,即兩個及兩個以上的鮞粒存在不連續的包裹關系,或者較小的鮞粒被包括在一個大的鮞粒之中(見圖4(i))。復合鮞粒類似于MARIOTTI G等[36]描述的由海底生物席包裹并通過膠結作用而形成的多核心鮞粒。
張夏組鮞粒中特別的白云石晶體,可能代表富有機質成巖微環境[10]條件下的白云石化作用,其有機基質,如EPS的降解釋放Mg2+,以及與之聯系的硫酸鹽還原反應促進碳酸鈣礦物的沉淀,使Mg2+/Ca2+相對升高而發生白云石化,形成這種特別的成巖白云石殘余物或燧石;也可能與鎂質黏土(如高嶺石和蒙脫石族礦物)有關,尤其是三八面體硅鎂石(Stevensite)前體白云石化的產物,代表生物礦化作用保留的初始結構[9,37]。
張夏組鮞粒還發育放射纖維狀方解石沉淀或結殼(見圖9)。方解石結殼不對稱地覆蓋在鮞粒最外部圈層上,不同鮞粒的放射纖維狀方解石結殼間有一個明顯的類似縫合線的邊界(見圖9(b)),表明這種結殼為同生或準同生形成的,類似于現代頭足類動物中生物控制的生物礦物(Biomineral)的形成。放射纖維狀方解石結殼可能形成于低Mg2+/Ca2+環境,Mg對放射狀方解石的形成有很大影響,至少對無機成因的放射狀方解石,低鎂利于其形成。自顯生宙以來,在“文石海”和“方解石海”中可見這類結殼物,而且葫蘆島張夏組鮞粒發育的放射纖維狀方解石結殼晶體內包含極薄的暗色泥晶薄膜(見圖9(c-d)),這種有機黏液薄膜的氧化使纖維狀方解石晶體形成并將其包裹在晶體內,代表富有機質條件下的放射纖維狀方解石的微生物沉淀作用[14,38];同樣,這樣的結殼也發生在由高密度的葛萬菌之類的絲狀鈣化藍細菌鞘構成的暗色泥晶微生物膜上(見圖5(e)),類似放射纖維狀方解石組構與微生物、某些宏觀藻類的關聯[39]。因此,鮞粒上的這種放射方解石結殼可認為是受微生物影響的碳酸鹽的原地沉淀作用。
發育在動蕩淺灘上的鮞粒比低能環境中的鮞粒更大些。SUMNER D Y等[40]運用數學模型闡釋控制鮞粒大小的因素,認為鮞粒的生長取決于質量損失和化學沉淀之間的平衡。該模型預測較大的鮞粒是由海水化學條件和水體擾動導致沉淀作用的加速而產生的,并假設鮞粒大小在懸浮過程中單調性增加,直到尺度超過其能在動蕩海水中懸浮的臨界值時停止。TROWER E J等[41]提出基于培養實驗的模型,將磨損率(Abrasion Rate)作為鮞粒大小和沉積物搬運機制(如拖曳和躍移)的函數,該模型否定SUMNER D Y等[40]的“鮞粒大小隨時間單調遞增關系”,并認為鮞粒大小在其大多數生長周期內處于動態平衡,且該平衡取決于沉積作用與磨蝕作用的相對速率。三道溝剖面寒武系張夏組發育含放射鮞粒的巨型鮞粒,代表相對海平面下降階段晚期的沉積記錄,表現寒武紀海洋“方解石海”的基本特點。葫蘆島張夏組中光合作用微生物膜主導的巨型鮞粒,是潮間到潮下間歇性高能環境產物,代表微生物和動蕩的水體環境之間復雜的相互作用[8,10]。
6 結論
(1)葫蘆島三道溝剖面寒武系苗嶺統張夏組頂部集中發育由巨型鮞粒構成的鮞粒灘。密集發育在鮞粒核心、皮層和外部泥晶套內的、可類比于葛萬菌和現代微鞘菌的絲狀或管狀鈣化藍細菌鞘化石,以及密集發育于鮞粒暗色泥晶紋層的黃鐵礦晶體,表明葫蘆島張夏組形成于典型的絲狀藍細菌主導光合微生物膜內復雜的鈣化作用和早期石化作用,以及微生物膜內EPS的異養細菌降解誘導的碳酸鹽礦物原地沉淀作用。
(2)發育于鮞粒內部的成巖白云石殘余物和密集的黃鐵礦晶體,以及圍繞鮞粒發育的放射纖維狀方解石結殼,代表EPS的降解或有機質的氧化及硫酸鹽的還原反應,與微生物影響的碳酸鹽礦物原地沉淀有密切關系。
(3)雖然成巖作用對巖石中的生物痕跡造成巨大破環,但三道溝剖面張夏組由絲狀藍細菌主導的鈣化微生物膜與高能水體相互作用而形成鮞粒的證據可成為鮞粒成因機制研究的參考。
