桂西北峰叢洼地農作區土壤真菌多樣性對不同耕作模式的響應
2021-08-02 10:39:30蔡蕓霜張建兵鐘麗雯陸雙龍黃麗排閆妍羅為群朱同彬胡寶清
生態學報 2021年12期
蔡蕓霜,張建兵,*,鐘麗雯,陸雙龍,黃麗排,閆妍,羅為群,朱同彬,胡寶清
1 北部灣環境演變與資源利用教育部重點實驗室, 南寧 530001 2 廣西地表過程與智能模擬重點實驗室, 南寧 530001 3 南寧師范大學 地理科學與規劃學院, 南寧 530001 4 國土資源部巖溶生態系統與石漠化治理重點實驗室, 桂林 541004
全球碳酸鹽巖分布面積達陸地面積的12%,其中我國分布有344萬km2,約占國土面積的1/3,我國西南更是全球三大碳酸鹽巖連續分布區之一[1]。峰叢洼地是該區典型的喀斯特地貌類型之一,面積約9.7萬km2[2]。在早期不合理人類活動的影響下,桂西北峰叢洼地區石漠化現象嚴重,導致當地土地生產力低、居民生活貧困[3]。從2000年開始,我國開始了大規模的石漠化治理工程,如封山育林、退耕還林還草、生態移民等,極大改善了該區的生態環境狀況[4]。但在該區還分布著較大面積的農作區,是當地居民生活、生產(主要為耕作活動)的集中區,對土壤的干擾強度大,土壤流失與石漠化風險高,是峰叢洼地區生態演變熱點區與生態保育的焦點區。因此,評價該區不同耕作模式的土壤安全效應,選擇適宜的耕作模式,對降低該區石漠化風險,保持土地可持續利用及持久脫貧具有重要意義。
土壤安全是指土壤持續地為人類提供食物、纖維和淡水資源等生態系統服務,同時維持生物多樣性和相對穩定性的一種狀態[5-7],是土壤物理、化學、生物性質的綜合反映。土壤微生物是地球化學循環中的主要驅動力,是土壤生態系統中最為活躍的部分,對生態系統的功能及其可持續性發揮著重要作用[8]。真菌作為土壤動植物殘體的分解者、互助者或病原體,其通過調節土壤能量流動和養分轉化、有機質積累、土壤結構形成、動植物殘體分解和抑制病蟲害等重要的土壤生態過程,在維持生態系統功能的基礎和生態系統健康方面發揮重要功能[9-10]。同時,土壤真菌對環境變化敏感,地理空間變化[11]、生境改變[12]、耕作模式變更[13]、植被演替[14]等都將引起真菌群落結構與多樣性的變化。因此,真菌群落與功能特征及其對外界環境變化的響應已成為評價土壤安全的重要指標之一[15-16]。
目前,耕作模式對土壤真菌影響的報道較多,大部分研究發現保護性耕作管理可增加微生物量的積累,改善土壤真菌特征[17-19]。關于耕作模式對喀斯特峰叢洼地土壤真菌影響的研究較少,該區已有研究主要集中在植被恢復的土壤,如劉雯雯等[20]對喀斯特區不同植被恢復階段土壤真菌進行分析,發現喬木林、灌木林階段土壤真菌群落結構相近,灌草過渡階段與草本群落階段真菌群落結構各成一類;Liang等[21]研究發現植被恢復過程中叢枝菌根真菌的多樣性依次為灌木≈原生林≈次生林<草地,何敏紅等[22]發現石漠化程度對真菌組成及多樣性的影響大于植被演替的影響,隨石漠化程度加深,土壤真菌多樣性總體呈現下降趨勢。因此,本文以典型峰叢洼地農作區—廣西果化巖溶生態研究基地為研究區,采用高通量測序技術,對不同耕作模式土壤真菌群落多樣性進行比較,并結合FUNGuild工具預測真菌功能,系統分析該區不同耕作模式對土壤真菌功能的影響,以期為該區土壤安全及農業生產的可持續性評價與管理提供科學依據。
1 材料與方法
1.1 研究區概況
廣西果化巖溶生態研究基地建于2001年,現為國家自然資源部野外科學觀測研究基地,位于廣西平果縣果化鎮西南方向約5 km處(107°22′30″—107°24′47″E,23°22′30"—23°24′32″N)。該區地處南亞熱帶季風氣候區,日照充足,年日照時數平均1682 h,日照百分率38%;氣候溫和濕潤,年均氣溫21.5℃,無霜期345 d以上;雨量充沛,平均年降雨量1400—1550 mm。該區屬典型的喀斯特峰叢洼地,地層主要為石炭系上統,巖石主要為純石灰巖。土壤類型主要為棕色石灰土。
該基地屬于典型的峰叢洼地農作區,人口密度高,耕作強度大。建設初期,該區植被覆蓋率低(不足10%),石漠化程度嚴重、類型齊全,生態極度脆弱。在封山育林、坡地退耕還林還草、土地整理、水土保持等系列石漠化治理措施下,該區石漠化得到了較好的控制,現植被覆蓋率已提高至75%左右,同時,洼地區和部分坡耕地得以繼續耕作,種植作物除傳統的玉米、甘蔗、水稻以外,還有引進作物火龍果。其多種模式的種植樣地為本研究的開展提供了良好的野外試驗平臺。
1.2 研究方法
1.2.1樣地選取
為評價研究區不同耕作措施對土壤真菌的影響,選擇該區主要的農田類型(耕作模式)——火龍果地(相當于保護性耕作模式,HF)、玉米地(相當于翻耕模式,MF)、甘蔗地(相當于少耕+秸稈還田耕作模式,SF)和水稻田(PF)為研究對象,并以退耕還林地(人工林,FL)、退耕還草(自然演替,AF)為對照樣地,各樣地情況介紹于表1。

表1 樣地描述
1.2.2樣品采集與處理
在2018年4月,利用土鉆(鉆頭直徑5 cm)采集不同耕作模式樣地土壤樣品,各模式選擇典型樣地至少3塊,每塊樣地采用S形布點法采集5點樣品,然后混合為該地塊樣品。采集土層為0—10 cm、10—20 cm、20—40 cm(退耕還草地僅有0—10 cm、10—20 cm)。共采集混合樣品51個,每個樣品一分為二,分別用于測試土壤微生物性質(裝入樣品袋后,速置于帶冰塊的保溫箱中,然后帶回保存于-80℃冰箱中)、土壤理化性質(裝入樣品袋帶回自然風干待處理)。
1.2.3土壤理化性質的測定
將自然風干后的土壤樣品,去除根系、礫石等雜質,按需磨碎過篩,然后分別采用酸度計法測定土壤pH 值(土水比1∶5,pH計S400-K,梅特勒-托利多國際貿易有限公司),K2Cr2O7氧化-外加熱法測定土壤有機質含量,凱氏定氮法測定土壤全氮含量(全自動凱氏定氮儀K1100F,濟南海能儀器股份有限公司),鉬銻抗比色法測定土壤全磷含量(紫外可見分光光度計L9,上海儀電分析儀有限公司),馬爾文激光粒度分析儀(Mastersize 2000,英國馬爾文儀器有限公司)測定土壤粒度[23]。各耕作模式下的土壤理化性質如表2所示。按照國際制土壤質地分級標準,火龍果地的土壤質地屬于壤質粘土,其他耕作模式土壤質地為粉砂質粘壤土。
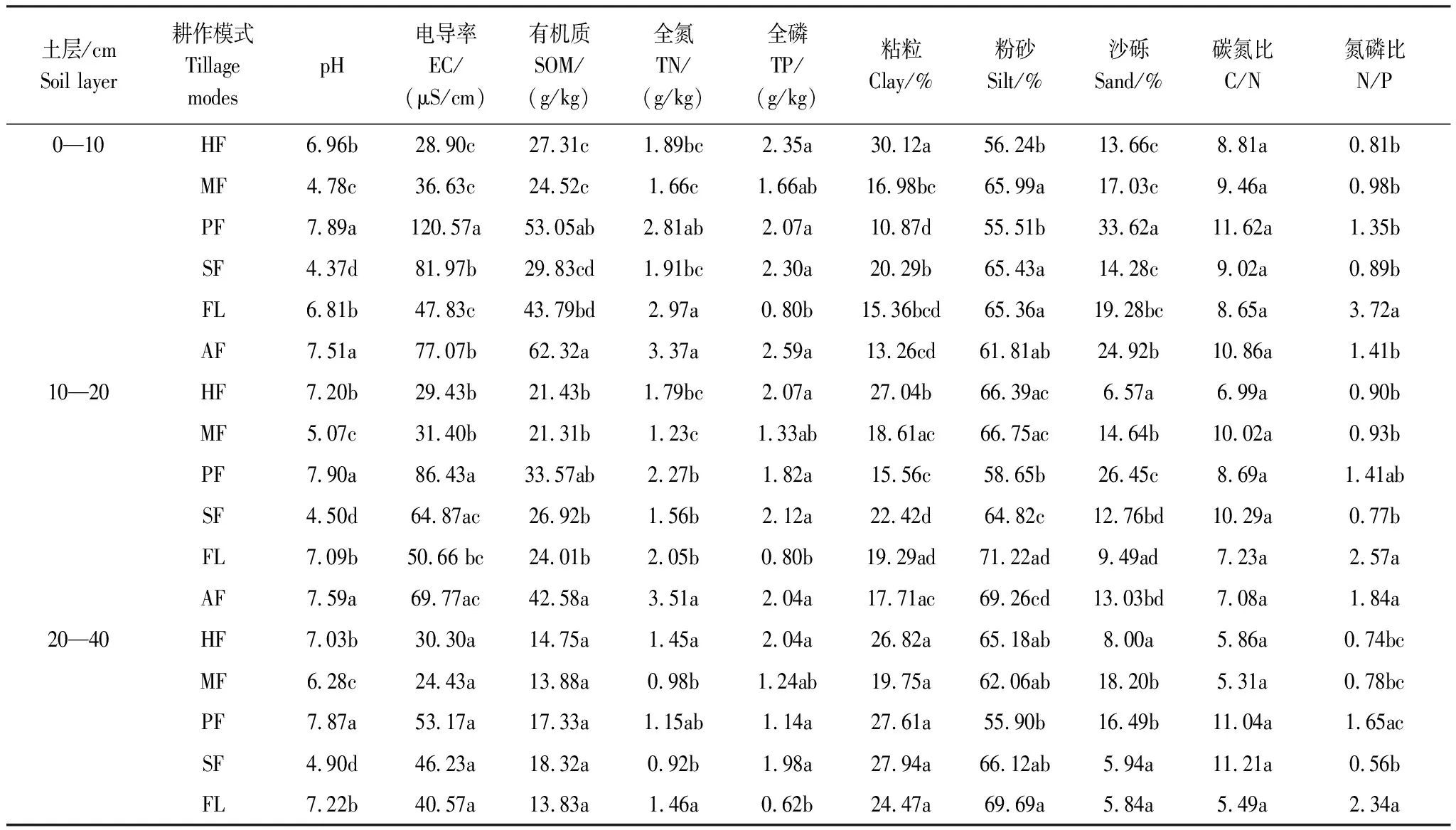
表2 不同耕作模式下的土壤理化性質特征
1.2.4樣品DNA 提取、擴增和高通量測序
土壤樣本DNA的提取采用CTAB標準法,DNA提取完成后利用濃度1%瓊脂糖凝膠進行電泳檢測其純度和濃度。選擇引物ITS5-1737F(5′-GGAAGTAAAAGTCGTAACAAGG-3′)和ITS2-2043R(5′-GCTGCGTTCTTCATCGATGC -3′)對土壤真菌ITS基因ITS1區進行PCR擴增。反應使用New England Biolabs公司的Phusion? High-Fidelity PCR和高效高保真酶進行PCR擴增,確保擴增效率和準確性。得到的產物使用2%濃度的瓊脂糖凝膠進行電泳檢測,對目的條帶使用Qiagen公司提供的膠回收試劑盒回收產物。根據所擴增的ITS1區域特點,基于Illumina HiSeq測序平臺,使用TruSeq? DNA PCR-Free Sample Preparation Kit建庫試劑盒進行文庫構建,構建好的文庫經過Qubit和Q-PCR定量,文庫合格后,使用HiSeq2500 PE250進行上機測序(由諾禾致源生物信息科技有限公司完成)。
1.2.5數據處理與統計分析
原始測序數據經過嚴格的過濾處理,去除標簽序列(barcode)、引物序列,過濾低質量和短長度的序列,得到高質量的Tags數據。運用Uparse軟件(Uparse,Version 7.0.1001)對序列進行聚類,默認以97%的一致性(Identity)將序列聚類成為OTUs(Operational Taxonomic Units),用Mothur方法與SILVA的SSUrRNA數據庫進行物種分類,在門類水平上統計各樣本的真菌群落組成。使用Qiime軟件(Quantitative Insights Into Microbial Ecology,Version 1.9.1)計算ACE指數、Shannon指數、Weighted Unifrac距離、構建UPGMA(Unweighted pair-group method with arithmetic means)樣本聚類樹等,ACE、Shannon指數箱型圖使用Oringin(Version 2017)軟件繪制,稀釋曲線(Rarefaction Curve)、維恩圖(Venn plot)、花瓣圖(Flower plot)、主坐標分析(Principal Co-ordinates Analysis,PCoA)、典型關聯分析(CanoicalCorrelation Analysis,CCA)、UPGMA聚類分析通過R軟件(Version 2.15.3)完成。基于線性判別分析流程[Linear discriminant analysis (LDA) effect size pipeline,LEfSe],使用LEfSe軟件分析,默認設置LDA Score的篩選值為3.5,分析分組樣本間具有顯著差異的Biomarker。真菌群落功能預測使用FUNGuild(Fungi+Functional+Guild)工具,該方法基于已發表文獻,對真菌的生態功能進行了歸類,構建FunGuild數據庫。土壤理化數據的統計分析和顯著性檢驗選用SPSS 22.0軟件。
2 結果與分析
2.1 土壤樣品測序深度及合理性分析
真菌ITS基因測序結果顯示,51個土壤樣本共檢測出4369767條序列,經過質量控制,總共得到4216941條高質量序列(每個土壤樣品序列數在47866—97427之間,平均值=82685)。以97%的相似度將序列聚類成為OTUs,得到11702個OTU。為檢測樣本的數據量是否合理,從樣品中隨機抽取一定測序量的數據,統計它們所代表物種數目(即OTUs數目),以抽取的測序數據量與對應的物種數來構建稀釋曲線(圖1),稀釋曲線表明,隨著測序量的增加,稀釋曲線逐漸趨于平坦,雖尚未達到飽和,但更多的測序量只會產生少量新的OTU,表明本次試驗的測序數據量合理,測序結果包含了絕大多數真菌類群,能夠較真實地反映研究區不同耕作模式土壤樣品的真菌群落,可用于分析不同耕作模式對土壤真菌群落結構與組成的影響。
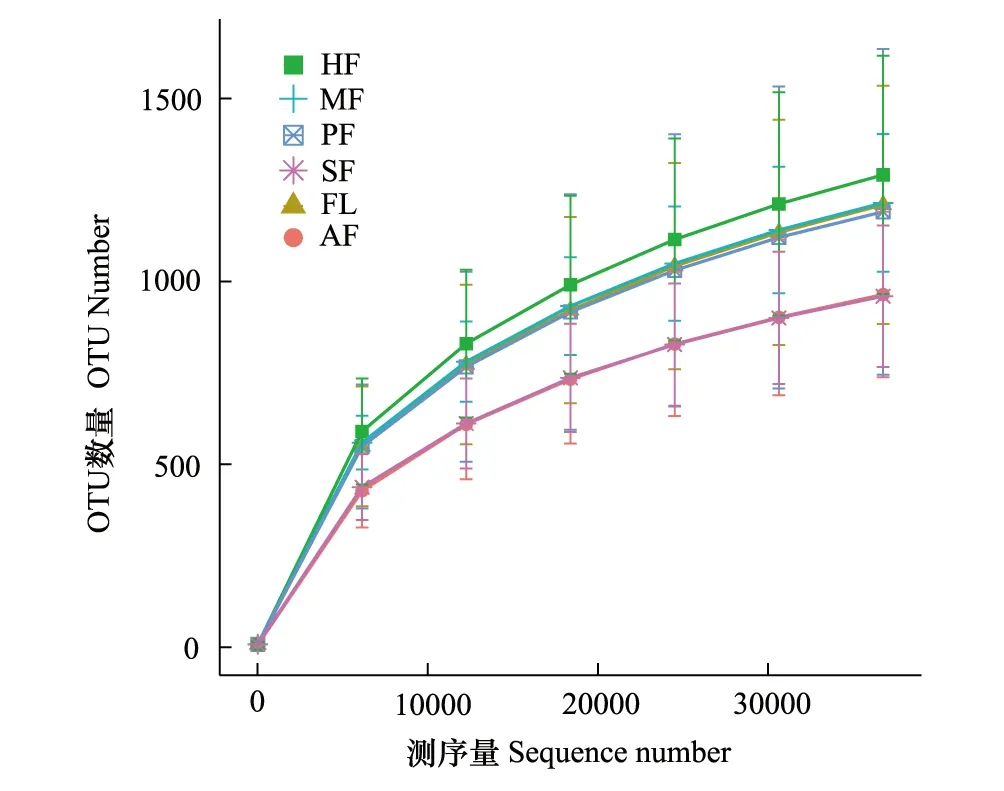
圖1 不同耕作模式土壤真菌稀釋曲線 Fig.1 Dilution curve of soil fungi under different tillage modesHF: 火龍果地 Pitaya field; MF:玉米地 Maize field; PF: 水稻田Paddy field; SF: 甘蔗地Sugarcane field; FL: 林地 Forestland; AF: 草地Grassland
2.2 土壤真菌α-多樣性分析
2.2.1土壤真菌群落豐富度及多樣性
微生物物種α-多樣性用于分析樣本內的微生物群落多樣性,本文包括代表土壤真菌群落豐富度的ACE指數和體現菌群的多樣性的Shannon指數。研究結果表明,各土層不同耕作模式土壤真菌豐富度和多樣性無顯著差異(P>0.05),20—40 cm土層土壤真菌多樣性(P<0.05)除外。土壤真菌ACE指數與Shannon指數在0—10 cm、10—20 cm土層中的變化趨勢較為一致,整體表現為HF、PF、MF、FL維持較高的土壤真菌豐富度及多樣性,SF、AF土壤真菌豐富度及多樣性較低;在20—40 cm土層,PF土壤真菌豐富度最低,而FL土壤真菌多樣性指數最低。隨著土壤深度增加,各耕作模式的土壤真菌ACE指數與Shannon指數均呈下降趨勢(圖2)。
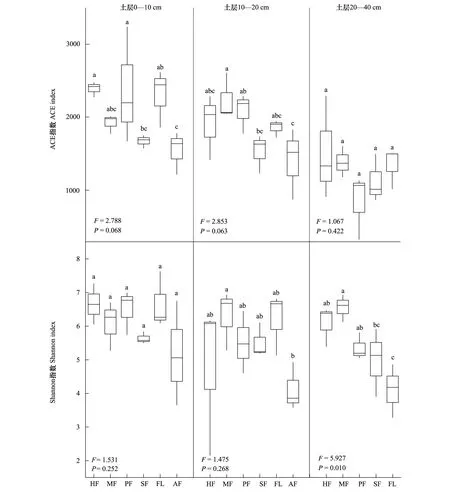
圖2 土壤真菌ACE指數、Shannon指數箱型圖Fig.2 Box plots of ACE index and Shannon index of the soil fungi不同小寫字母表示差異性顯著(P<0.05)
2.2.2土壤真菌群落組成分析
研究區土壤真菌ITS高通量測序所得序列經物種注釋后分屬18門、63綱、163目、370科、860屬、1049種。在門類水平上,相對豐度排名前10的有Ascomycota(子囊菌門)、Basidiomycota(擔子菌門)、Mortierellomycota(被孢霉門)、Rozellomycota(羅茲菌門)、Glomeromycota(球囊菌門)、Olpidiomycota(油壺菌門)、Mucoromycota(毛霉門)、Chytridiomycota(壺菌門)、Zoopagomycota(捕蟲霉門)、Blastocladiomycota(芽枝霉門),其中前3者為優勢菌門(相對豐度均>1%),3者相對豐度總和為25.93%—83.78%。SF的Ascomycota相對豐度(69.877%—74.242%)明顯高于其他耕作模式(9.312%—60.194%),同時PF的Ascomycota相對豐度(9.312%—20.598%)最低;FL的Basidiomycota相對豐度(11.864%—12.550%)比其他耕作模式(2.704—10.212%)高,除0—10 cm土層的HF(19.099%);SF、AF的Mortierellomycota相對豐度明顯低于HF、MF、PF和FL;Rozellomycota在MF的相對豐度(0.646%—2.875%)最高;Glomeromycota在FL具有最高的相對豐度(0.848%—2.486%);PF的Olpidiomycota相對豐度(0.171%—1.804%)最高;Mucoromycota在HF具有最高的相對豐度(0.457%—1.379%);HF、PF、MF的Chytridiomycota相對豐度高于SF、FL、AF;Zoopagomycota、Blastocladiomycota在各耕作模式土壤的相對豐度無明顯差異。各耕作模式土壤真菌群落組成中均存在相對豐度較高的目前數據庫無法鑒定的真菌門。
隨土層深度的增加,HF、SF的Ascomycota相對豐度表現為先增加后減少的趨勢,MF、PF、FL則表現為先減少后增加的趨勢,AF表現為增加的趨勢;HF、MF、FL、AF的Basidiomycota表現為遞減趨勢,而PF表現為先減少后增加,SF則是先增加后減少的趨勢;HF、PF、SF、AF的Mortierellomycota相對豐度隨土壤深度增加逐漸減少,MF、FL表現為增加趨勢(圖3)。
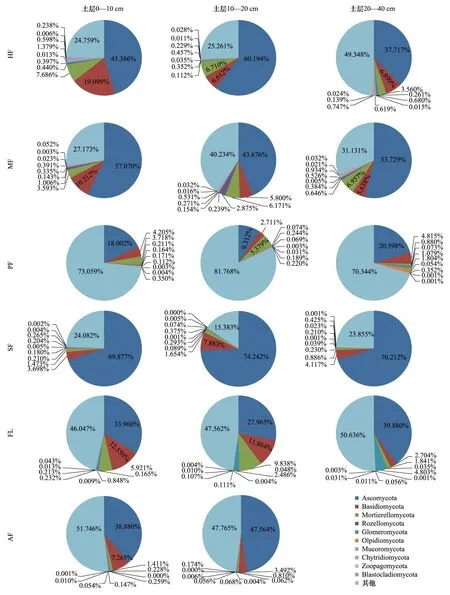
圖3 土壤真菌群落在門類水平上的組成及相對豐度Fig.3 Composition and relative abundance of the taxonomic classes of soil fungi
2.2.3土壤真菌群落共有OTUs與特有OTUs
花瓣圖和Venn圖可直觀展現并反映組間或樣品之間OTU數量組成相似性、重疊情況以及特異性。在0—10 cm、10—20 cm、20—40 cm土層,各樣地間土壤真菌群落共有OTU數量分別為300、275、194,占各層真菌總數比例分別為3.47%、3.35%、3.00%。
土壤真菌特有OTUs方面, PF在0—10 cm土層的土壤真菌特有OTUs(1050)最多,占真菌總數的12.15%,MF的土壤真菌特有OTUs比例最低(3.09%),其他耕作模式的土壤真菌特有OTUs占比在5.76%—7.53%之間;10—20 cm土層中,PF土壤真菌特有OTUs(958)最多,占真菌總數的11.68%,HF的土壤真菌特有OTUs比例最低(3.78%),其他耕作模式的土壤真菌特有OTUs比例在5.83%—7.81%之間;在20—40 cm土層,HF特有OTUs(914)的比例最高,為14.16%,其他耕作模式的土壤真菌特有OTUs的比例在9.79%—11.21%之間(圖4)。

圖4 不同土層土壤真菌群落共有OTUs及特有OTUsFig.4 Shared OTUs and specific OTUs of fungal communities in different soil layers
同一耕作模式不同土層間土壤真菌群落大體相似,共有OTUs數量大于各層特有OTUs數量,0—10 cm與10—20 cm土層間的共有OTUs數量大于0—10 cm與20—40 cm共有OTUs數量,并整體表現出表層擁有最大數量的OTUs總量與特有OTUs數量。HF、MF、PF、SF、FL、AF不同土層共有OTU數目分別為936、960、516、2829、964、1133,占各耕作模式全部OTUs數量的19.01%、22.19%、11.22%、22.30%、23.75%、41.19%(圖5)。
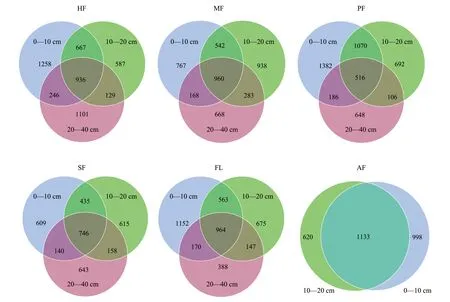
圖5 同種耕作模式的不同土層間土壤真菌共有OTUs及特有OTUsFig.5 Total and specific OTUs of soil fungi between different soil layers under the same farming mode
2.3 土壤真菌β-多樣性分析
2.3.1基于Weighted Unifrac距離的聚類分析
該區土壤樣本真菌的聚類親緣關系分析結果顯示(圖6),不同耕作模式、土層之間的土壤真菌群落結構具有一定的差異性。耕作模式方面,AF、FL為一簇,MF、SF、HF為一簇,PF單獨成簇。土層之間,0—10 cm、10—20 cm土層土壤真菌群落相似性較高,與20—40 cm土層真菌群落相似度不高。
2.3.2土壤真菌LEfSe分析
各耕作模式具有不同數量和類型的顯著差異物種(Biomarker),數量方面,FL(33)、SF(29)和 HF(28)具有較多的顯著性差異物種,MF(16)、PF(12)、AF(4)最少;在門類水平上,不同耕作模式中,SF的顯著差異真菌門為Ascomycota,PF為Olpidomycota,MF為Rozellomycota,HF為Mortierellomycota,FL為Glomeromycota,AF在門類水平無顯著差異物種(圖7)。
2.4 土壤真菌群落結構與土壤環境因子的CCA分析
分析結果可以看出,同種耕作模式的土壤樣本能夠清晰地聚集在一起,說明同一耕作模式生境相似,哺育了相似的真菌群落;其中FL、AF的樣本距離比較接近,HF、MF、SF樣本距離接近,與PF相距較遠,說明FL、AF土壤真菌群落結構相近,HF、MF、SF土壤真菌群落結構相似;PF土壤真菌群落結構聚集為單獨的一簇,與旱地系統差異明顯,與β-多樣性分析結果一致。軸1和軸2的特征值分別為15.78%和14.62%,共解釋了30.40%的土壤真菌物種-環境關系,說明理化因子對土壤真菌群落結構有一定的影響。土壤pH、TN、N/P的射線較長,粒度(砂礫、粉砂、粘粒)含量、SOM、EC、TP、C/N的射線較短,表明pH、TN、N/P是影響土壤真菌群落的重要環境因素,其中pH是最主要的影響因子,土壤pH值與PF、FL、AF的真菌群落結構呈正相關,與HF、MF、SF土壤真菌群落結構呈負相關(圖8)。

圖8 OTU 水平上基于 Weighted Unifrac差異的CCA分析Fig.8 Weighted UniFrac distance-based CCA analysis at the OTU levelTN: 全氮 Totalnitrogen; TP:全磷 Total phosphorus;EC:電導率Electrical conductivity;SOM: 有機質Soil organicmatter
2.5 土壤真菌群落功能預測
研究區土壤真菌主要分為病理營養型、腐生營養型和共生營養型以及其他無法鑒定營養型的種群,將注釋獲得功能的 OTUs進行統計,無法鑒定營養型的種群占據土壤真菌成分的絕大部分,說明目前還缺少對這些土壤真菌功能的認識。在土壤真菌功能類群中,腐生營養型真菌比例最大,其次為病理營養型真菌,共生營養型真菌的比例最小。耕作模式對土壤功能類真菌影響明顯,HF、MF、SF、PF土壤腐生營養型真菌、病理營養型真菌的含量顯著高于FL、AF,說明耕作活動提高腐生營養型真菌含量的同時也刺激病理營養型真菌的生長;相對其他耕作模式,SF明顯促進土壤腐生營養型真菌的生長。值得說明的是,各耕作模式中,HF明顯提高共生營養型真菌數量,表明保護性耕作有利于共生營養型真菌的富集(圖9)。
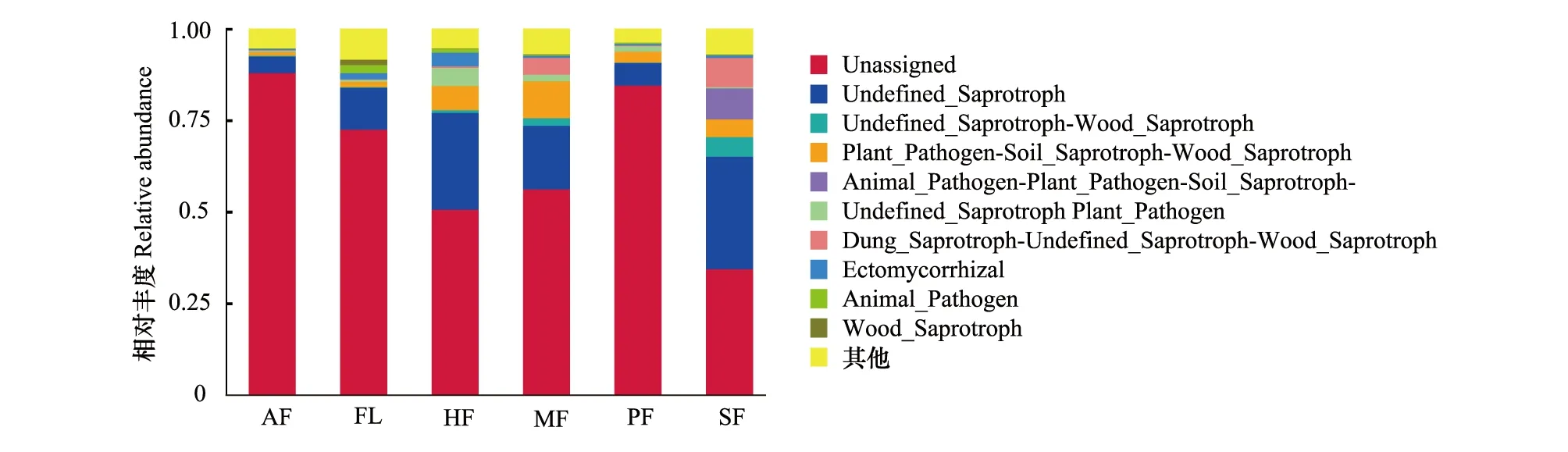
圖9 不同耕作模式土壤真菌群落功能預測Fig.9 Functional predictions of the fungal communities in soils under different tillage modes
3 討論
3.1 耕作模式對土壤真菌α-多樣性的影響
本文對桂西北峰叢洼地土壤真菌多樣性的研究結果表明,不同耕作模式下的土壤真菌ACE指數和Shannon指數整體表現為:HF、MF、PF、FL的土壤真菌豐富度和多樣性明顯提高,而SF、AF豐富度和多樣性降低。其原因可能與石灰土質地粘重有關,因為石灰土的成土母質主要為碳酸鹽巖風化溶蝕存留的酸不溶物,且多為粘土成分。而耕作活動可增加土壤通透性,利于土壤水、熱和養分的遷移轉化,從而利于土壤真菌的生存繁衍。隨著土壤深度增加,各耕作模式下的土壤真菌的豐富度和多樣性均有下降趨勢。這主要是由于深層土壤缺乏通透性,其土壤溫度、氧氣、活性碳庫等狀況均變差,土壤真菌多樣性也隨之降低[24]。
各耕作模式所共有的OTUs數量大于各自特有的OTUs數量,表明耕作模式對土壤真菌群落產生影響,但整體上沒有改變某一區域土壤真菌群落的主要構成。真菌群落組成分析顯示Ascomycota、Basidiomycota、Mortierellomycota是研究區土壤的優勢菌門,與喀斯特區的相關研究結果相似[25];這也表明耕作活動一般不會顯著改變土壤優勢真菌的種類,但會影響其相對豐度。
3.2 耕作模式對土壤真菌β-多樣性的影響
通過對不同耕作模式下土壤真菌群落結構之間的相似性和差異性的分析,發現HF、MF、SF(旱作)土壤真菌結構相似,FL、AF(退耕)土壤真菌群落結構相似,均與PF(水田)土壤真菌群落結構差異明顯,同時,隨著土壤剖面深度增加,土壤真菌群落結構也存在差異。這是由于土壤水分、通氣性、溫度、養分等土壤真菌生存狀況不同,導致土壤真菌豐度、組成存在差異,進而引起真菌群落結構存在差異。值得指出的是,盡管FL與AF土壤真菌α-多樣性不相似,但β-多樣性卻接近,可見,α-多樣性并不總是與β-多樣性存在正相關[26]。
3.3 土壤理化性質對真菌群落結構的影響
土壤理化性質的變化會直接或間接地影響土壤微生物群落特征[27]。多數研究發現,土壤 pH是決定細菌群落變化的主要因素,而真菌群落結構通常與土壤養分含量密切相關[28-29]。然而,該區土壤真菌群落結構受土壤pH值影響最為明顯。同時,TN、N/P對土壤真菌群落結構顯著相關[30],有研究表明,當微生物氮的利用率過高時,會抑制N 吸收相關酶的活性,刺激P 吸收相關酶的活性[31-33],加劇土壤對磷的需求[34],說明土壤氮、磷可促進真菌物種之間的相互作用[35]。
3.4 耕作模式對土壤真菌群落功能的影響
從土壤微生物群落中的關鍵真菌功能類群來看,如果選擇適宜的耕作措施,可以預防或減少植物根系病害的發生和危害,在保持土壤質量和健康方面可發揮至關重要的作用[36]。本研究結果顯示,該區土壤中未鑒定營養型真菌含量較高,可見,這類真菌類群還無法依據現有研究結果進行區分,還缺少深入研究。該區土壤真菌功能類群中,腐生營養型真菌比例最大,其次為病理營養型真菌,共生營養型真菌的比例最小。耕作模式對土壤功能類真菌影響明顯,HF、MF、SF、PF土壤腐生營養型真菌、病理營養型真菌的含量顯著高于退耕模式(FL、AF),耕作活動在提高腐生營養型真菌含量的同時,也刺激病理營養型真菌的生長,說明耕作活動較為頻繁的土壤容易造成腐生營養型真菌的富集,同時增加了從宿主細胞獲取營養來源、對植物生長具有一定危害影響的病理型真菌[37]。SF 明顯促進腐生營養型真菌的生長,這可能是田間填加秸稈作為有機肥的措施為腐生真菌的生長提供了營養。已有研究結果表明,共生營養型真菌可能在作物健康、營養和品質方面發揮重要作用[38]。值得說明的是,保護性耕作(HF)共生營養型的真菌類群最為豐富,共生營養真菌在作物健康、營養和品質方面發揮著重要作用,可降低農田病蟲害風險,促進作物生長,提高產量與品質。因此,相對其他耕作活動來說,HF是對土壤健康和作物生長的較優處理方式。FUNGuild雖然在一定程度上解析了真菌的功能,但由于這種方法是基于已有文獻和數據,具有一定的局限性,因此,依該方法確定的土壤真菌類群結構和功能群數據庫仍有待進一步完善。
4 結論
農作區是喀斯特峰叢洼地區土地利用與生態環境變化的熱點區域,生態風險高,耕作活動是引發該風險的導火線之一。本研究認為,不同耕作模式通過改變土壤性質與環境狀況,改變了土壤真菌群落構成、豐度和功能。保護性耕作模式(HF)的土壤真菌多樣性與群落結構具有與退耕模式相似的特征,且HF土壤共生營養型真菌最為豐富,能較好地維持土壤真菌群落結構特征和提高作物健康和品質,具有良好的生態環境效應。此外,本研究中保護性耕作模式的火龍果還具有較高的經濟效益,因此,火龍果種植模式可作為峰叢洼地農作區生態、經濟雙提升的一種耕種模式。
