超親優勢數量遺傳學理論推導及其在玉米育種中的應用
2021-08-05 13:51:26陳澤輝王安貴祝云芳郭向陽劉鵬飛
西南農業學報 2021年6期
關鍵詞:效應
陳澤輝,吳 迅,王安貴,祝云芳,郭向陽,劉鵬飛
(貴州省農業科學院旱糧研究所,貴州 貴陽 550006)
【研究意義】雜種優勢是遺傳基礎不同的2個親本的雜交一代在生長勢、存活力、生殖力和抗性、品質等方面均優于雙親的現象[1]。雜種優勢是非常重要的生物學現象,超親優勢則作為一種雜種優勢表現形式,被廣泛用于生產實踐,但對其認識仍處于不斷加深中,對超親優勢數量遺傳學理論進行推導并應用于實踐,對豐富玉米育種理論具有重要的意義。【前人研究進展】我國北魏《齊民要術》一書對馬和驢雜交產生騾的現象進行了詳細記載,騾表現出較馬和驢強壯和高大,且騾的壽命比馬和驢都長。這種子代表現超過親本的現象很好地體現了超親優勢,為人類認識和利用超親優勢開辟了成功案例。在農業上,雜種優勢已被廣泛應用于水稻、玉米、高粱、油菜等植物及家畜和家禽等動物育種上,并已取得了明顯效果。【本研究切入點】關于雜種優勢形成的遺傳基礎較為復雜,其理論研究較多,對雜種優勢理論的探索卻遠遠落后于應用實踐,雜種優勢形成的遺傳基礎仍停留在假說階段。關于雜種優勢理論尤其是超親優勢的認識仍然處于不斷加深中。【擬解決的關鍵問題】在綜述前者研究成果的基礎上,結合多年的玉米自交系和雜交種選育實踐,基于加性-顯性模型對超親優勢數學表達式進行推導,并對超親優勢可能的遺傳基礎及其在指導玉米自交系和雜交種選育方面的應用進行探討,以期提高玉米育種效率,為玉米育種的理論指導提供參考。
1 超親優勢數量遺傳學理論推導
1.1 中親優勢的數量遺傳學理論
數量性狀是由多個位點基因控制,往往表現出很強的雜種優勢現象。為推導出雜種優勢的數學表達式,以具有2個等位基因G和g的單個位點為例,A和B兩個隨機交配群體,雜交產生的F1往往表現出雜種優勢,其中一個群體等位基因G和g的頻率分別是p和q;另一個群體同一位點上的等位基因頻率分別為p′和q′;兩群體間同一位點同一等位基因頻率之差為y,則y=p-p′=q′-q。把p′和q′寫作p-y和p+y。按基因型尺度(圖1)理論,GG、Gg和gg的基因型值分別為a、d和-a[2-3]。
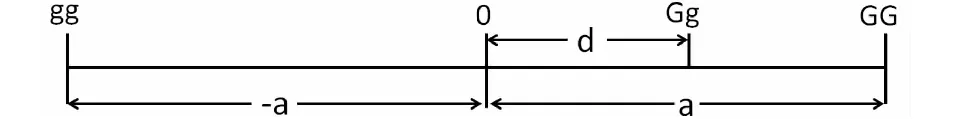
圖1 基因型尺度
按基因型尺度,A群體的平均值:
MPA=a(p-q)+2dpq
B群體的平均值:
MPB=a(p-y-q-y)+2d(p-y)(q+y)=a(p-q-2y)+2d[pq+y(p-q)-y2]
即2個群體雜交產生F1時,從一個群體隨機得到的個體與從另一個群體隨機得到的個體交配獲得F1的基因型頻率列于表1。

表1 2個群體雜交后代的基因頻率和基因型頻率

表2 在2個Hardy-Weinberg平衡群體雜交組合1個位點2個等位基因下的基因型值和基因型頻率
F1的平均基因型值:
MF1=a(p2-py-q2-qy)+d[2pq+y(p-q)]=a(p-q-y)+d[2pq+y(p-q)]
中親優勢值的度量表示在F1與中親值間的差值,則:
式中,d為顯性效應,若該位點無顯性,則不會產生雜種優勢。Y為兩群體該位點基因頻率之差,若兩群體間基因頻率無差異,則不會產生雜種優勢。考慮兩群體與研究性狀有關的所有位點的聯合效應,不考慮位點間的上位效應,即,在加性-顯性模型下,F1總的中親優勢如下。
1.2 超親優勢的數量遺傳學理論
超親優勢指雜交組合F1的表現高于高值親本或低于低值親本的現象。超顯性假說由Shull[4]提出,經East[5]利用基因理論將此觀點具體化。按基因型尺度模型導出超親優勢的數學表達式。
1.2.1 超高值親本的超親優勢 假定A群體為高值親本,B群體為低值親本。在單個位點情況下,超高值親本的超親優勢數學推導如下。
mPA=a(p-q)+2dpq
MPB=a(p-q-2y)+2d[pq+y(p-q)-y2]
MF1=a(p-q-y)+d[2pq+y(p-q)]
HPHF1=MF1-MPA=a(p-q-y)+d[2pg+y(p-q)]-a(p-q)-2dpq=a(p-q-y-p+q)+d[2pg+y(p-q)-2dpq]=-ay+yd(p-q)=y[d(p-q)-a]
若雙親均為純系(純合自交系),p為1,q為0,則超高值親本的超親優勢如下:
HPHF1=a(1-y)+dy-a=a-ay+dy=y(d-a)
從上式可看出,在單個位點時,超親優勢等于雙親同一個基因的頻率之差y與該位點顯性效應d和加性效應a之差的乘積。若y為0,若雙親是均含該基因的純合系,該位點無超親優勢。y為1時,即親本1含該基因,親本2不含該基因,超高值親本的超親優勢如下:
HPHF1=d-a
從上式可知,在單個位點時,只有顯性效應d大于加性效應a才有超親優勢。
1.2.2 超低值親本的超親優勢 在單個位點情況下,超低值親本的超親優勢數學推導如下:
LPHF1=MF1-MP2=a(p-q-y)+d[2pq+y(p-q)]-[a(p-q-2y)+2d[pq+y(p-q)-y2]=a(p-q-y-p+q+2y)+d[2pq+y(p-q)-2pq-2y(p-q)+2y2]=ay-dy(p-q)+2dy2
LPHF1=MF1-MPB=a(p-q-y)+d[2pq+y(p-q)]-[a(p-q-2y)+2d[pq+y(p-q)-y2]=a(p-q-y-p+q+2y)+d[2pq+y(p-q)-2pq-2y(p-q)+2y2]=ay-dy(p-q)+2dy2
若雙親均為純系(純合自交系),p為1,q為0,則:
LPHF1=ay-dy+2dy2
y為1時,即親本A群體含該基因,親本B群體不含該基因,那么,超低值親本的超親優勢如下:
LPHF1=a+b
從上式得出,在單個位點時,只有顯性效應d的絕對值大于加性效應a才有超低值親本的超親優勢。
雙親的基因頻率差異y為0。
LPHF1=ay-dy+2dy2=0
1.2.3 多位點下的超親優勢 考慮兩群體與研究性狀有關的所有位點的聯合效應,不考慮位點間的上位效應,即在加性-顯性模型下,F1總的中親優勢為多個位點超親優勢的累加。
數量性狀由多個位點基因控制,如圖2中A和G位點。A位點內A基因和a基因與G位點內G基因和g基因間的相互作用為上位性互作關系,多數情況下位點間的上位性不顯著,或占比較小。當上位性互作為正時,優勢會增大,當上位性互作為負時,優勢會降低。超高值親本的超親優勢數學表達式HPHF1=Σy(d-a),及超低值親本的超親優勢數學表達式LPHF1=Σ(d+a),在加性-顯性模型條件下這2個數學表達式才成立,但上位性可能對雜種優勢表現有所貢獻。在無上位性互作情況下,只有在多個位點的(d-a)的代數和為正時,才有超親優勢。則需要多個位點的顯性效應是超顯性效應時,雜種F1才會表現出超親優勢。說明,超親優勢主要是超顯性在起作用。
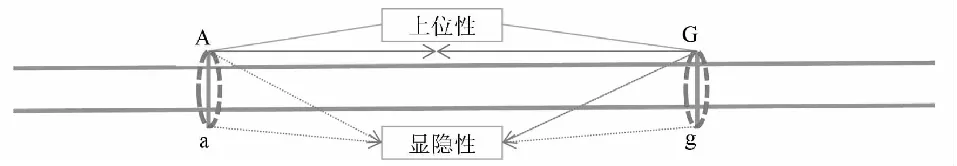
A位點內A基因與a基因間及G位點內G基因與g基因間的相互作用均為顯隱性關系,A位點內A基因和a基因與G位點內G基因和g基因間的相互作用為上位性互作關系
上位性效應在雜種優勢表現中存在潛在作用[6-7]。但是上位性互作效應只能解釋玉米群體中微不足道的遺傳變異,而加性和顯性方差能夠解釋除此之外的大部分遺傳變異。上位性似乎并不是遺傳變異的主要成分,上位性應該表現在由優良自交系組成的特定雜交種中,因此,上位性的相互作用似乎在特定的,而不是在雜交組合間普遍存在[8]。
Hallauer等[8]為獲得加性a、顯性d組分和3種上位性組分(aa、ad和dd)的方差,利用從玉米群體BSSS中獲得的一系列隨機自交系進行的研究結果表明,加性組分占總產量遺傳方差的93%;加性與顯性共占99%。說明,上位性對產量的遺傳變異為次要貢獻因素[9]。
考慮到上位性對雜種優勢可能存在的影響,超親優勢的數學表達式為HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a)。除基因頻率之差外,還考慮雙親互補性,在此基礎上形成雜種優勢的基因頻率差異互補的理論觀點,可用于指導玉米育種實踐。
如具有4個位點開展的數量性狀,不同基因型AABBccdd和aabbCCDD的2個自交系雜交,F1的基因型為AaBbCcDd。假設顯性基因A對某一數量性狀的貢獻為12,B的貢獻為10,C的貢獻為8,D的貢獻為6;相應的隱性等位基因的貢獻分別為6、5、4、3。在不存在上位性時,親本AABBccdd的表現型值為12+10+4+3=29,另一親本aabbCCDD的表現型值為6+5+8+6=25。根據顯性基因的效應可知F1的表現型值。若無顯性效應,雜合的等位基因Aa、Bb、Cc、Dd的貢獻值均等于相應的等位顯性基因和隱性基因的平均值,(12+6+10+5+8+4+6+3)/2=27,與雙親的平均值相同,無雜種優勢。若表現部分顯性,則P1表現型值大于中親值偏向高值親本,表現出部分雜種優勢,即AaBbCcDd>27。若表現為完全顯性,則F1大于高值親本,雜種F1為AaBbCcDd=12+10+8+6=36,表現出超親雜種優勢。利用超親優勢的數學表達式HPHF1=y[d(p-q)-a]分別計算各位點的超親優勢。
對A位點,雙親平均值m=9,a=3,d=3,y=1,p=1,q=0,超親優勢:HPHF1=1×[3(1-0)-3]=0,F1=P1+HPHF1=12+0=12,對B位點,雙親平均值m=7.5,a=2.5,d=2.5,y=1,p=1,q=0,超親優勢:HPHF1=1×[2.5(1-0)-2.5]=0,F1=P1+HPHF1=10+0=10,對C位點,雙親平均值m=6,a=2,d=2,y=-1,p=0,q=1,超親優勢:HPHF1=-1×[2(0-1)-2]=4,F1=P1+HPHF1=4+4=8,對D位點,雙親平均值m=4.5,a=1.5,d=1.5,y=-1,p=0,q=1,超親優勢:HPHF1=-1×[1.5(0-1)-1.5]=3,F1=P1+HPHF1=3+3=6。
4個位點總的超親優勢:
HPHF1=∑y[d(p-q)-a]=(0+0+4+3)=7
基因型AaBbCcDd的表現:
總的F1=12+10+8+6=36,從單個位點看,超親優勢的遺傳基礎是超顯性效應。多個位點控制的數量性狀,顯性效應對超親優勢的表現起主導作用,其中包括超顯性和完全顯性,或者部分顯性;不同位點的顯性程度可能不一,一個位點可能是顯性效應,另一位點可能是超顯性或部分顯性。另外,顯性效應的方向也可能不一,總的效應是多個位點顯性方向的代數和。
與中親優勢一樣,超親優勢大小與雙親基因頻率有關,在顯性效應和加性效應相對固定時,雙親基因頻率差異越大,超親優勢越強,這對指導玉米自交系選育和雜交組合組配具有重要意義。
2 超親優勢理論在作物雜種優勢上的應用
2.1 幾種基因作用方式對雜交種表現的影響
在單個位點,雜種F1的表現等于高值親本群體的平均值,加上中親優勢。若在多個位點情況下,可能存在上位性。因此幾種基因作用方式,即加性效應、顯性效應和上位性對雜交種的表現有重要影響。
基因的加性對雙親群體的平均值起作決定作用。超親優勢HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a),含有加性效應a。一個群體或自交系的一般配合力主要由基因的加性效應決定。因此,在作物育種中通過測定一般配合力來確定材料(群體、自交系等)具有加性基因及其效應的多少。反之,要提高一個純系或自交系的一般配合力,則需用優勢強的雜交組合進行二環選系、復合雜交選系和輪回選擇后選系等方法來提高優良基因數量,提高總的遺傳效應。即在一個自交系中集中更多的加性基因來提高親本自交系的平均值[2]。
經驗證據已表明,自交系的相對活力、抗性、產量潛力等在連續多個循環的選擇中不斷增加,此為美國玉米帶單交種成功的原因[10],玉米二環系的產量較一環系高30.8%。另外,Collins[11]研究表明,若控制一個性狀的因子對數太多,則很難獲得所有基因都純合的植株。控制一些性狀,如活力、抗性、產量等的基因數量未知,但可以肯定其數目很大。在玉米育種實踐中,從整個基因組的角度看,雙親間的基因差異不大。所以育種上盡可能用雙親基因差異大的自交系組配雜交組合,達到雜種優勢最大化,則需建立雜種優勢群。玉米育種的不同雜種優勢群體是人為創造和保持的,不同國家以及同一國家的不同育種單位所采用的雜交優勢群不一。在育種實踐中,育種者有固定的雜種優勢群是首要,并在相應優勢群下對雙親不斷互為改良,才能不斷地培育出好品種,這一點尤其在正確利用二環系選擇中具有指導意義[12]。
在孟德爾遺傳中,顯性是非常重要的遺傳現象。顯性遺傳有完全顯性、部分顯性和超顯性,呈正向顯性和負向顯性。顯性效應的大小和方向,既因作物不同,同一作物不同性狀而不同,也因控制數量性狀的不同位點而不同。但同一位點,顯性效應大小和方向是相對固定的。同一數量性狀有關位點的定向顯性,若不同位點顯性方向不同,則會相互抵消,削弱總的顯性表現。
顯性效應的重要性因組合和性狀而有很大變化。在中親優勢的數學表達式MPHF1≈Σdy2中,顯性效應對中親優勢的出現非常重要,無顯性效應,就無雜種優勢,顯性效應的正負決定雜種優勢的方向。在雙親均為純系時,各位點的中親優勢等于顯性效應。超親優勢的數學表達式:HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a),超顯性對超親優勢起決定作用。
上位性是指不同位點基因間的相互作用,可能是2個位點,也可能是3個或多個位點基因間的相互作用。上位性效應極其復雜,具體效應值很難估算。因此,在數量遺傳的研究模型中,多假定無上位性或上位性不顯著;若上位性顯著,也簡單采用加性×加性、顯性×顯性和加性×顯性進行分類,很難確定是哪些基因間的相互作用及效應值大小。Hallauer[8]解釋上位性效應在育種中的影響認為,若上位性方差的比例較小,則可忽略上位性偏差對選擇進度的影響。
2.2 超親優勢理論在作物雜種優勢上的應用
超親優勢的數學表達式HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a)。由于多數情況下,上位性對性狀表現的作用相對較小,且上位性作用相當復雜,在此重點考慮加性和顯性效應,及雙親基因頻率差異對作物雜交種表現的影響,從而指導育種實踐。
2.2.1 基因頻率差異大或顯性程度高能夠提高雜種優勢 要提高雜種優勢,須增加雙親間基因頻率差異和尋找顯性程度高的基因位點。除利用商業栽培品種外,從基礎群體進行選系是一個重要途徑。很多育種材料可被育種者作為半成品引進,大多源自農家品種或傳統的原始栽培品種,這些原始栽培品種經過數百年甚至上千年的演化被自然和人為選擇所馴化,其內部多樣性豐富。玉米起源于熱帶亞熱帶地區,當地豐富種質資源是實現現代玉米產業發展目標的重要育種材料來源,特別是經CIMMYT等機構改良過可應用于生產的各類品種。此外,充分利用與栽培品種關系較近的野生種為另一個重要途徑。這些近緣野生種或是栽培作物的祖先,或與作物親緣關系較近(大芻草為典型例子),這些種質資源正變得日益重要。
2.2.2 兩群雙向選擇育種模式 利用中親優勢MPHF1≈Σdy2和超親優勢HPHF1≈Σy(d-a),以有利顯性基因分散在雙親群體中,這一理論能較好地指導玉米自交系和雜交種的選育[13]。
(1)玉米雜種優勢類群劃分與利用。美國玉米育種者根據自交系的來源把自交系劃分為Reid和Lancaster 2個優勢群,發現兩者間有很強的雜種優勢,形成了玉米研究中最典型的雜種優勢利用模式。迄今為止,美國利用時間最長、使用范圍最廣的仍然是這2個雜種優勢群及由其組成的雜種優勢模式。歐洲最常用的模式為美國馬齒型×歐洲硬粒型;在熱帶地區則以ETO復合品種×Tuxpeňo為主要模式;巴西以馬齒型×Tuxpeňo或Suwan1×Tuxpeňo 為常用模式。
國內主要雜交種的親本自交系大致可分為7個雜種優勢類群:唐四平頭群、旅大紅骨群、改良Reid群、Lancaster群、PA群、PB群、Suwan群和Tuxpeňo群等[14]。分別從這些類群中選育自交系,然后類群間自交系雜交,獲得強優勢雜種組合的頻率較高。西南地區主要的玉米雜種優勢模式:改良Reid群×Suwan群、改良Reid群×PB群、Tuxpeňo群×Lancaster群和Suwan1×Tuxpeňo等。
(2)兩群相互輪回選擇模式及其利用。兩群相互輪回選擇,即2個群體互為測驗種,不僅改良2個群體本身,更重要的是改良了群體間雜交組合的表現。該改良方法是基于雜種優勢的數學表達式,重點是增加2個群體間基因頻率差異,既增加了2個群體的加性效應,也提高了群體間非加性基因效應。群體的一般配合力和2個群體間的特殊配合力均得到提高。
Chen等[15]對溫熱群體墨瑞(TR)和蘇蘭(SL)進行3輪相互輪回選擇,群體間雜交組合的籽粒產量提高了13.89%,達當地推廣雜交種(貴單8號)的產量水平(圖3)。同時,群體間雜交組合的中親優勢從15%增至25.96%,超親優勢從10.91%增至16.49%(表3),而吐絲期表現為負向超親優勢。目前,已分別從墨瑞和蘇蘭改良群體中選育出自交系,并組配出優良組合。
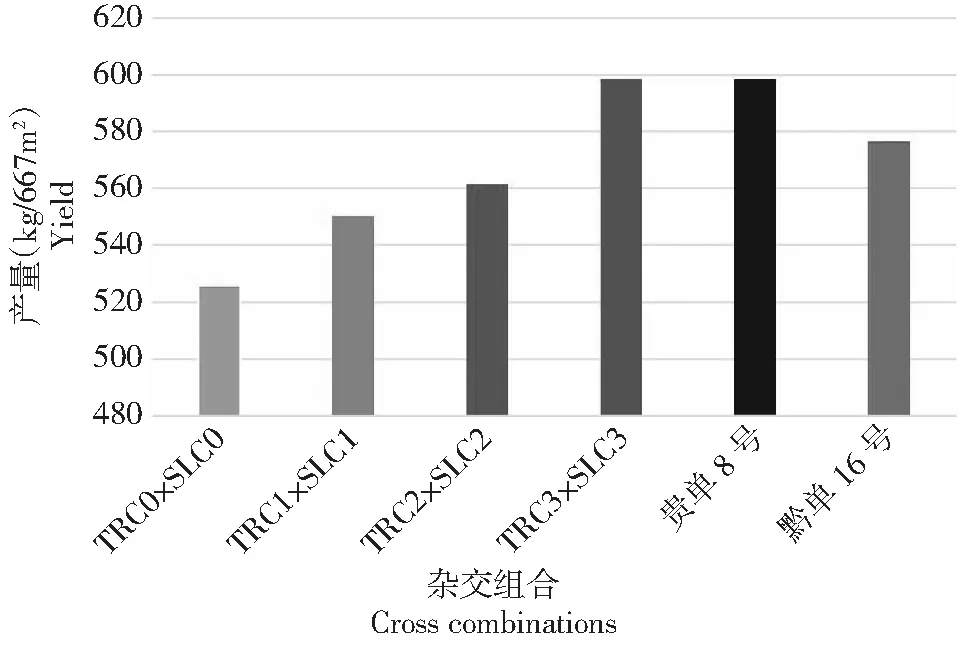
圖3 玉米TR1群體×SL1群體相互輪回選擇的籽粒產量
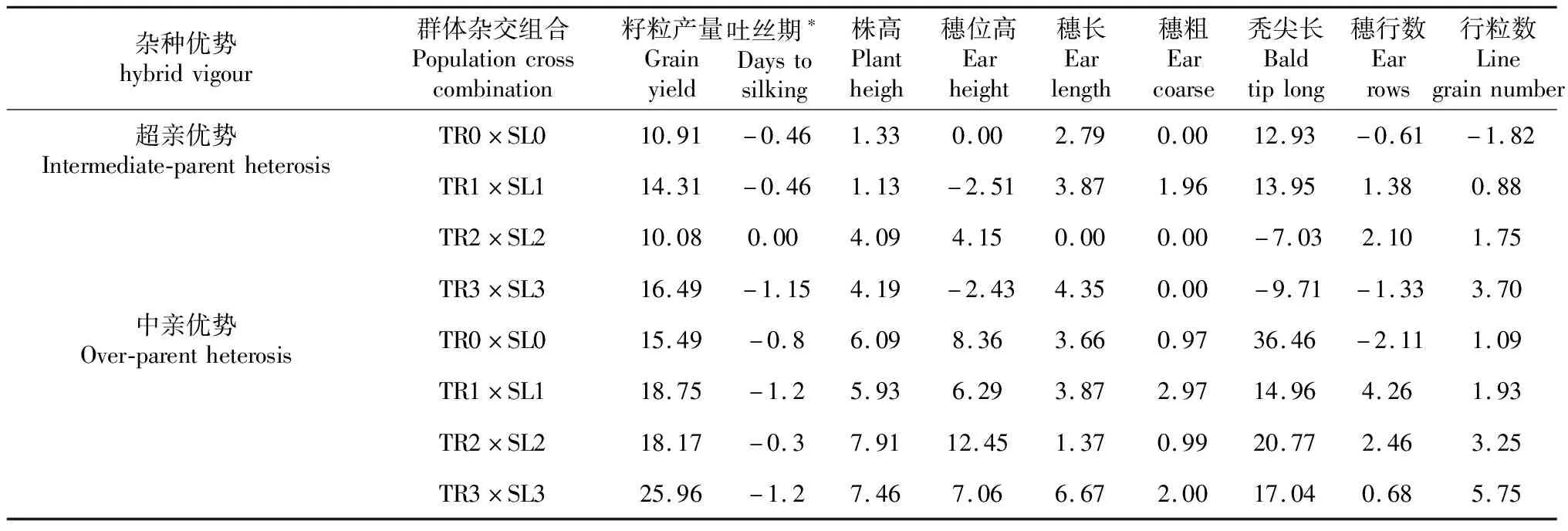
表3 玉米群體間雜交組合的超親優勢和中親優勢
(3)雜種優勢的全基因組預測。基因組預測是利用訓練群體的基因型結果和表型數據,建立最佳線性無偏預測模型,得到訓練群體的基因組估計育種值(GEBV)。在此基礎上,根據育種群體的基因型數據,基于BLUP模型和與訓練群體中的等位基因同一性預測育種群體各性狀表現值,從而得到各個體具有優良表型的估計值,即高育種值。研究表明,影響全基因組預測準確性的因素有用于估計基因組估計育種值的標記數、性狀遺傳力、參考群體大小、統計模型、分子標記的數量和類型、連鎖不平衡、有效群體大小、參考和預測群體的關系和群體結構[16],其中訓練群體的結構是影響多親本群體基因型預測準確性的重要因素,且雙向抽樣能夠有效提高基因組預測效率。基于兩群體雙向選擇獲得材料所具有的遺傳獨立性,它們間雜交會產生很強的雜種優勢,所獲得的后代雜交群體具有系譜清晰、遺傳結構簡單且遺傳重組較雙親分離群體多的優點,利用該群體做訓練群體,可降低標記密度。加上2個類群間雜交后代與類群親本間所產生的同胞關系,可進一步減少對訓練群體大小的要求。
2.2.3 群間選系組配模式及其利用 按雜種優勢的數學表達式,2個不同類群間雜交F1的表現最強,說明集中了較多的有利顯性基因,雙親差異大。若用F1雜交組合選育自交系,在多次混合授粉和加大后代群體種植數量,則有可能選育出集中更多優良基因的自交系,然后與其他類群自交系組配,育出優良組合的頻率會增加,此為群間選系組配模式[2]。該模式的難點是在群間選系后,尋找可組配形成優勢雜交組合類群,或改良創新類群。在不同的生態類型區,如黃淮海區、東華北區、西北區和西南區,改良創新類群和組配模式可能會有所變化。目前較成功的群間選系組配創新模式有[改良瑞德(BSSS)×Lancaster]×(黃改×旅系)、[改良瑞德(BSSS)×Lancaster]×(黃改×歐洲硬粒)、[改良瑞德(BSSS)×Lancaster]×Suwan1、[改良瑞德(BSSS)×Lancaster]×[Suwan1×Iodent]及[Tuxpeo×改良瑞德(BSSS)]×(Suwan1×Lancaster)等。
筆者所在團隊用PB群的自交系9195與屬于改良瑞德群的丹9046雜交,從F1自交后代中育出優良自交系QB506。QB506與屬于Suwan1類群自交系T32和QR273組配,分別育出優良雜交種金玉818和金玉506,在西南地區被大面積推廣應用。因此,(PB群×改良瑞德)× Suwan1是適宜西南地區玉米自交系和雜交種的選育模式。
3 討 論
對雜種優勢形成的機理研究顯示,顯性效應、超顯性效應和上位性均可能對雜好優勢有貢獻,并且均有試驗和實驗數據支撐。但都不能很好地解釋整個雜種優勢的形成機理。
徐云碧[17]認為,雜合性極其相關的基因互作是解釋雜種優勢的主要遺傳學基礎,因為雜交種對于親本間有差異的所有基因位點均為雜合的。因此,雜種優勢的程度取決于何種基因位點是雜合的,以及基因位點內等位基因和基因位點間等位基因彼此間如何互作。基因位點內等位基因的互作導致顯性、部分顯性或超顯性,理論上的顯性程度為0(無顯性)至大于1(超顯性)。基因位點間等位基因的互作導致上位性。遺傳作圖結果表明,雜種優勢和其他數量性狀中涉及的大多數QTL具有顯性效應,隨著能夠更有效地估計上位性方法的出現,上位性已被更頻繁地發現,并被證明是數量性狀遺傳控制中的一個普遍現象。然而,對于不同雜交組合、物種或性狀,其雜種優勢須通過不同程度的顯性結合所有可能的基因位點間互作來解釋。對雜種優勢的充分理解將依賴于與雜種優勢有關的所有基因的克隆和功能分析。
現今使用的雜交種是通過雜交2個高度近交的系,即自交系而產生的F1雜種。無論采取何種育種方式,關注點都集中在F1代比2個親本更優。多年來提出的4種解釋:一是2個親本間顯性或部分顯性基因的分散;二是在各個基因位點上的超顯性;三是2個基因位點相斥相有利等位基因緊密連鎖而導致的擬超顯性;四是上位性[18]。這些解釋相互不排斥,因此需確定其在任何特定雜交種中的相對重要性,但實踐證明很難確定,說明這些遺傳成分在雜種優勢形成的遺傳基礎上不是相互排斥的。因而雜種優勢形成雖有多個假說,但一直存在爭議[19]。
隨著對雜種優勢研究的深入,對雜種優勢的理解越來越復雜,很難將已有研究結果應用于育種實踐。通過對前人研究結果的分析認為,對雜種優勢的形成,基因的加性效應、顯性效應、超顯性和位點間基因的上位性都有作用,會因材料遺傳背景不同、位點不同、基因不同而在這3個方面的基因作用方式上存在差異。基因的加性效應和顯性效應普遍存在,而位點間的上位性因特定的材料和位點才顯著存在,并非普遍現象;或是與加性效應和顯性效應相比,其效應相對較低,大多數情況下似乎可以忽略不計。
為玉米指導育種實踐,對雜種優勢的理解應盡可能簡單化,抓主要的或普遍存在的,并且在遺傳方差成分中占比較大的部分。雜種優勢利用的最終目的是育出產量高的F1雜交種,因此,加性效應的重要性是第一位的,它決定雙親的平均值;其次,顯性效應也非常重要,決定F1雜交種的雜種優勢,不管是中親優勢還是超親優勢,是顯性效應還是超顯性起作用,均是不同性狀或是同一性狀不同位點的差別而已。上位性對雜種優勢的影響不可預見,且隨遺傳背景的差異,在育種中可以暫時忽略。雙親群體基因頻率差異對雜種優勢的影響顯而易見,如基因頻率差異y為零,則無雜種優勢;相反,y值達最大的1時,雜種優勢最大。因此認為,中親優勢和超超親優勢的數學表達式MPHF1≈Σdy2和HPHF1≈Σy(d-a)能很好地用于指導玉米育種實踐。
4 結 論
在綜述前者研究成果的基礎上,結合多年的玉米自交系和雜交種選育實踐,對超親優勢數量遺傳學理論的推導結果顯示,基因型尺度模型下的中親優勢和超親優勢的數學表達式分別為MPHF1≈Σdy2及HPHF1≈Σy(d-a),均能很好地用于指導玉米育種實踐。
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
科學大眾(2020年17期)2020-10-27 02:49:10
紅土地(2018年11期)2018-12-19 05:10:56
意林·全彩Color(2018年9期)2018-11-13 22:49:38
中學物理·高中(2016年12期)2017-04-22 11:53:03
中國衛生(2016年4期)2016-11-12 13:24:14
中國衛生(2014年4期)2014-12-06 05:57:14
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
