三七PnAAE41基因的克隆、表達分析及原核表達
2021-08-26 21:34:39閆靜向貴生馮壘楊梅郭敏張廣輝
熱帶作物學報 2021年7期
閆靜 向貴生 馮壘 楊梅 郭敏 張廣輝

摘? 要:草酰輔酶A合成酶參與了三七主要止血活性成分三七素的生物合成,研究三七草酰輔酶A合成酶基因(PnAAE41)的分子特征及表達特征,從而完善三七素生物合成途徑以及成分積累調控機制等方面的研究。以三七為實驗材料,在前期轉錄組研究基礎上,設計特異性引物,通過RT-PCR擴增獲得1個三七AAE基因的cDNA全長。采用生物信息學方法分析其分子特性,通過qRT-PCR技術分析PnAAE41基因的時空表達特異性。通過異源表達探索進行PnAAE41最佳蛋白最佳的誘導條件的探索,隨后以顏色反應作為定性研究的基礎。序列分析結果表明,PnAAE41的開放閱讀框長度為1572 bp,編碼523個氨基酸,登錄號為MW242343。系統進化分析顯示,PnAAE41蛋白與擬南芥AtAAE蛋白親緣關系最近。預測PnAAE41蛋白主要定位于細胞質膜上,不含信號肽序列,二級結構中α-螺旋和無規則卷曲占大部分比例。qRT-PCR分析表明,PnAAE41基因主要在根和莖中優勢表達。異源表達最佳誘導條件為0.5 mmol/L異丙基-β-D-硫代半乳糖苷(IPTG),16 ℃誘導9 h,該蛋白對草酸具有催化作用。本研究結果為進一步深入研究AAE在三七素生物合成途徑中的功能提供理論基礎。
關鍵詞:三七;草酰輔酶A合成酶;生物信息學分析;時空表達;原核表達
中圖分類號:S567.23+6????? 文獻標識碼:A
Clone and Prokaryotic Expression Analysis of Acyl-activating Enzyme Gene from Panax notoginseng
YAN Jing1,2,3, XIANG Guisheng1,2, FENG Lei1,2,3, YANG Mei3, GUO Min3, ZHANG Guanghui1,2*
1. National & Local Joint Engineering Research Center on Germplasms Utilization & Innovation of Chinese Materials in Southwest/ , Yunnan Agricultural University, Kunming, Yunnan 650201, China; 2. Yunnan Provincial Key Laboratory of Medicinal Plant Bio?logy, Yunnan Agricultural University, Kunming, Yunnan 650201, China; 3. College of Agriculture and Biotechnology, Yunnan Agricultural University, Kunming, Yunnan 650201, China
Abstract: Acyl-activating enzyme (PnAAE41) of Panax notoginseng is involved in the biosynthesis of dencichine,the main hemostatic active ingredient. The molecular characteristics and expression characteristics of Acyl-activating enzyme was explored to provide a basic theory for further research on the biosynthesis pathway of dencichine and the regulation mechanism of component accumulation. The full length of PnAAE41 gene was obtained by RT-PCR. The molecular properties was analyzed by bioinformatics methods. The spatio-temporal expression specificity of PnAAE41 gene was determined by real-time quantitative PCR. The heterologous expression was used to explore the optimal induction condition. The color reaction was used as the basis for qualitative research. The sequence analysis results showed that PnAAE41 was 1572 bp, encoding 523 amino acids, and the login number was MW242343. Phylogenetic analysis showed that PnAAE41 was most closely related to Arabidopsis AtAAE. PnAAE41 was predicted to be mainly located on the plasma membrane, which did not contain signal peptide sequence, and its secondary structure was mainly composed of alpha helix and random coil. The optimal induction condition was 0.5 mmol/L IPTG and 16 ℃ induction for 9 h. The protein had a catalytic effect on oxalic acid. The study would provide a theoretical basis for the further research on the role of AAE in the biosynthesis of dencichine.
Keywords: Panax notoginseng; acyl-activating enzyme; bioinformatics analysis; spatio-temporal expression; prokaryotic expression
DOI: 10.3969/j.issn.1000-2561.2021.07.008
三七[Panax notoginseng (Burk) F. H. Chen]為五加科人參屬多年生草本植物,以根及根莖入藥,是我國重要的中藥材大品種。三七具有散瘀止血、消腫定痛、防止腦缺血、止血、降脂和保護肝臟等功效,與人參共享“北參南七”的美譽。三七總皂苷廣泛用于治療冠心病、心絞痛、腦血管后遺癥和高血壓等。云南省一半以上的制藥企業需要三七為原料,其加工產品產值占云南醫藥產值的“半壁江山”,因此三七是云南乃至我國的中藥產業最重要的支柱品種。前期對于三七活性成分的研究主要集中于三萜皂苷,其他藥用成分則鮮有關注。三七素(dencichine)是一種非蛋白游離氨基酸,化學名稱為β-N-草酰-L-α,β-二氨基丙酸(β-N-oxalyl-L-α,β-diaminopropionic acid,β-ODAP),是云南白藥、三七止血片等藥品的有效成分,具有增加血小板數量、縮短凝血和出血時間的功效[1-2]。除三七外,三七素還存在于人參(Panax. Ginseng C. A. Meyer)、西洋參(Panax. Quinquefolius L.)等其他人參屬(Panax)植物以及山黧豆屬(Lathyrus)、豬屎豆屬(Crotalaria)等豆科植物中[3-6]。盡管三七素是三七的主要活性成分之一,但目前對其在三七中的生物合成機理、積累規律等研究未見相關報道。
Malathi等[7]在對家山黧豆(Lathyrus sativus L.)的研究中發現,草酰輔酶A(Oxalyl coenzyme A)與L-α,β-二氨基丙酸(L-α,β-diaminopropanoic acid, DAP)在ODAP合成酶作用下產生β-ODAP,這是三七素生物合成的最后一步,也是最為關鍵的一步反應。現已證實,家山黧豆ODAP合成酶屬于BAHD酰基轉移酶基因家族,能夠催化上述反應,但其基因序列并未公布[8]。草酰輔酶A是由草酸和輔酶A在草酰輔酶A合成酶的作用下生成的,因此,草酰輔酶A合成酶也是三七素生物合成的關鍵酶之一。研究發現草酰輔酶A合成酶屬于酰基激活酶(Aacyl-activating enzyme,AAE)超家族,其中還包括長鏈酰基輔酶A合成酶(Llong-chain acyl-CoA synthetases, LACSs),4-香豆酸輔酶A連接酶(4-coumarate CoA ligases,4CLs)和乙酰輔酶A合成酶(Aacetyl-CoA synthetase)[9-10]。
植物體內合成草酰輔酶A是草酸分解代謝的一部分,能夠避免植物受到草酸鈣積累的傷害,并且在植物的非生物和生物脅迫反應中都能發揮作用[11-12]。有研究從擬南芥(Arabidopsis thaliana L. Heynh.)[13]、蒺藜苜蓿(Medicago truncula L.)[14]、赤小豆(Vigna umbellate Thunb.)[15]與酵母(Saccharomyces cerevisiae)[16]中均克隆到1條草酰輔酶A合成酶基因,并對其功能進行了驗證,但在三七中的研究及其在三七素合成中的功能則未見報道。本研究基于三七轉錄組數據克隆到的1條草酰輔酶A合成酶基因PnAAE41,對其結構與功能進行了初步探索,為進一步研究三七素的生物合成與積累調控機制等方面奠定基礎,為深入了解三七品質形成機理提供了理論依據,也為在其他物種中三七素的生物合成提供數據線索。
1? 材料和方法材料與方法
1.1? 實驗材料與試劑
三七材料采自于云南省文山壯族苗族自治州硯山縣文山苗鄉三七科技有限公司(23°06′N,103°43′ E)。
二硫蘇糖醇(DTT)、磷酸烯醇丙酮酸(PEP)、肌激酶、還原型輔酶I(NADH)、1-甲氧基-5-甲基-吩嗪硫酸甲酯和四唑硝基藍購自麥克林公司;丙酮酸激酶、乳酸脫氫酶、腺嘌呤核苷三磷酸(ATP)購自索萊寶公司;輔酶A(CoA)為MedChemExpress公司產品;pET-28a載體、BL21(DE3)感受態購自上海唯地。
1.2? PnAAE41基因的鑒定與克隆
以33年生三七不同組織(根、莖、葉)轉錄組數據為基礎,從Pfam 33.1數據庫(http://pfam. xfam.org/)下載AAE基因家族種子文件PF00501,利用HMMER v3.0軟件從三七轉錄組數據中提取AAE同源序列,使用NCBI CDD(https://www.ncbi. nlm.nih.gov/Structure/cdd/wrpsb.cgi)工具進行手工校正,應用ORF Finder(https://www.ncbi.nlm. nih. gov/orffinder/)工具獲得基因的開放閱讀框,隨后同Pfam 33.1和SMART[17]數據庫進行結構域預測,獲得最終候選基因AAE。本研究中其他物種的氨基酸序列均來自于NCBI數據庫。
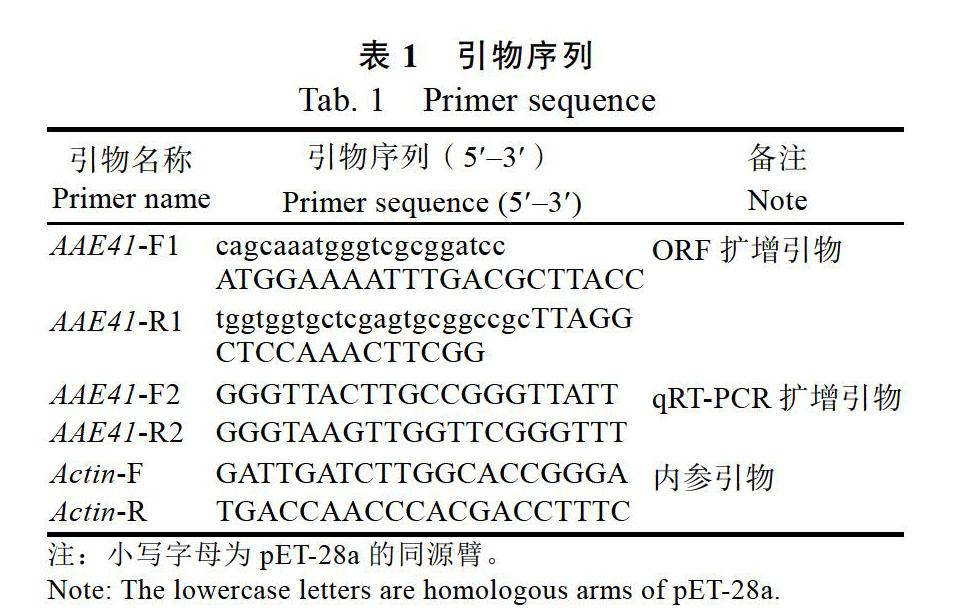
采集33年生三七的不同部位(根、莖、葉),參照難提植物總RNA小提試劑盒(MaGen)的操作說明提取三七總RNA,隨后使用逆轉錄試劑盒(TakaraTaKaRa)進行逆轉錄反應,將獲得的cDNA于–80 ℃備用。設計特異性引物AAE41-F1和AAE41-R1對PnAAE41全長進行擴增(表1),反應體系:cDNA 2.0 μL,上下游引物各1.6 μL,Q5 2×Master Mix 25.0 μL,加ddH2O補至總體積為50.0 μL;反應程序:98 ℃ 30 s;98 ℃ 10 s,50 ℃ 30 s,72 ℃ 50 s(35個循環);72 ℃ 2 min。1.0%的瓊脂糖凝膠電泳切膠回收。將PnAAE41基因回收產物與pET-28a線性化載體進行連接,熱激轉化,挑取陽性克隆菌株測序。
1.3? PnAAE41蛋白的生物信息學與系統進化分析
利用Expasy Protparam(https://web.expasy. org/ protparam)分析蛋白的理化性質;使用TMHMM(http://www.cbs.dtu.dk/services/TMHMM)預測蛋白的跨膜結構域;通過PSORT(http://psort1.hgc. jp/form.html)進行亞細胞定位預測;利用SignalP(http://www.cbs.dtu.dk/ services/SignalP0)預測蛋白質有無信號肽;使用GORIV(https://npsa-prabi. ibcp.fr/cgi-bin/ secpred_ gor4.pl)預測蛋白二級結構;利用SWISS-MODEL(https://swissmodel. expasy.org/ interactive/Sbb BSU/ models)預測蛋白的三維結構,并構建三維結構模型。利用MAFFT v7.471軟件進行氨基酸序列比對;IQ-TREE v2.1.1軟件用于構建ML系統進化樹(模型:LG+F+R5),并使用EvolView 3(https://www.evolgenius.info/ evolview/)進行進化樹編輯與美化。
1.4? PnAAE41基因的時空表達特異性分析
以Actin為內參基因[18],使用Primer 5.0工具設計qRT-PCR特異性引物AAE-F2和AAE-R2,Actin-F和Actin-R(表1)。按照ChamQ Universal SYBR qPCR Master Mix(Vazyme)試劑盒的反應體系:cDNA 1.0 μL,上下游引物各0.4 μL,SYBR Mix 10.0 μL,加ddH2O補充至總體積為20.0 μL。反應程序:95 ℃ 30 s;95 ℃ 15 s,60 ℃ 30 s(40個循環);72 ℃ 30 s。采用2–△△CT法計算目的基因的相對表達量[19]。
1.5? PnAAE41重組蛋白的誘導及純化
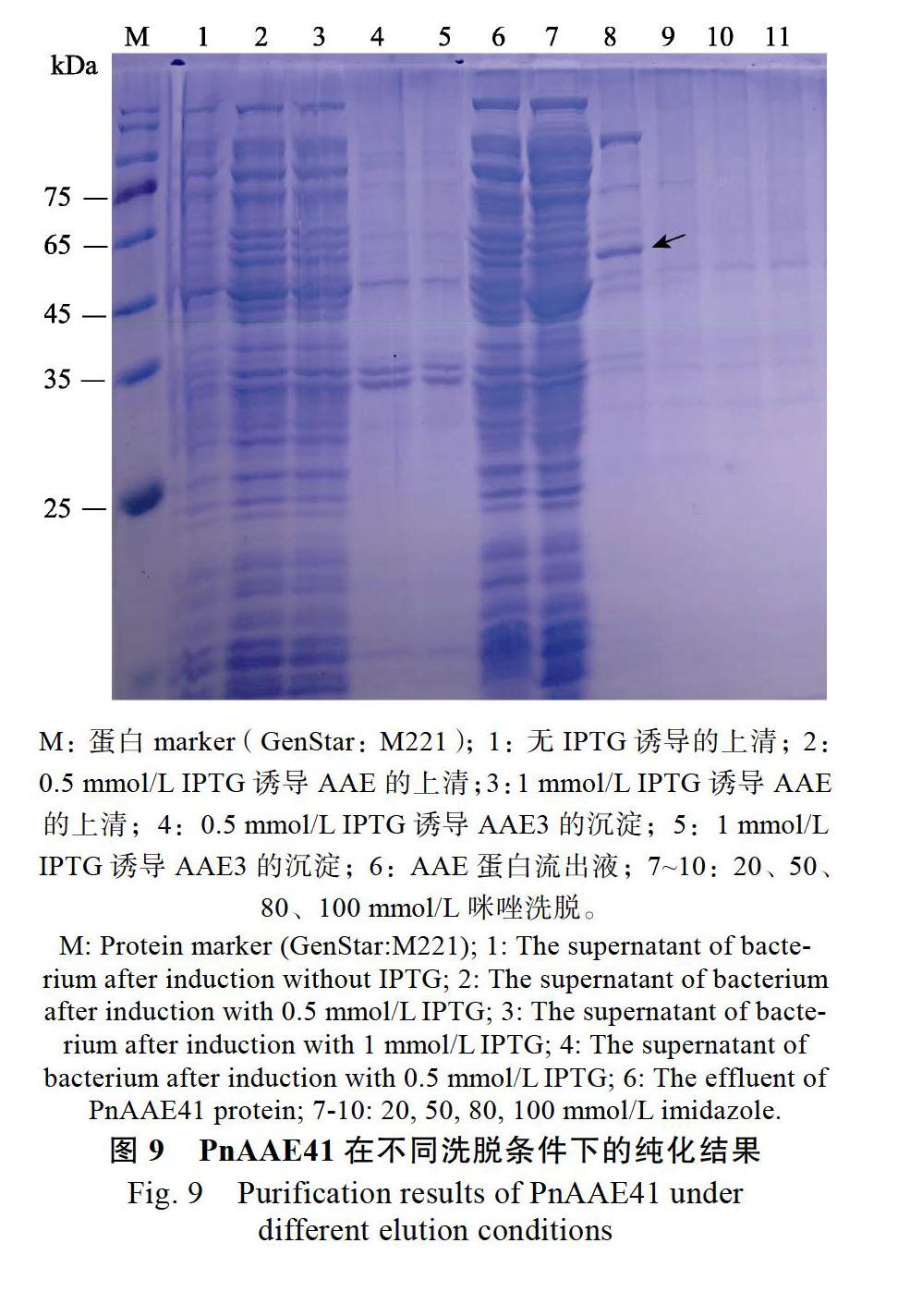
將通過測序驗證的陽性菌株接種到含50 mg/L Kan+的LB液體培養基中,并在37 ℃ 200 r/min條件下過夜培養,按1∶100(V/V)的接種量接種后,同條件下培養至A600為0.6~0.8,隨后對不同誘導溫度(16、37、25 ℃)、不同誘導濃度(0.1、0.5、1.0 mmol/L)異丙基-β-D-硫代半乳糖苷(IPTG)與不同誘導時間(8、12、16 h)條件下蛋白誘導的最佳條件進行了篩選。在最佳的誘導條件下進行擴大培養,4 ℃ 5000 ×g離心20 min收集菌體,用PBS(pH 8.0)重懸菌體;低溫高壓破碎細胞至菌液澄清透明,離心收集上清與沉淀。上清用0.45 μm濾膜過濾,然后按照天地人和Ni NTA Beads預裝柱純化說明書對其進行純化;沉淀用PBS(pH 8.0)重懸;隨后對其進行SDS-PAGE檢測,并分析蛋白表達情況。
1.6? PnAAE41蛋白催化活力檢測
反應體系緩沖液中含有0.479 mg重組蛋白、NaPO4(pH 8.0)、DTT、MATP、MgCl2、CoA、NADH、PEP,以及肌激酶、丙酮酸激酶、乳酸脫氫酶各10個單位,使用不同濃度(0.0、0.5、1.0、2.0、5.0、8.0、10.0 μmol/L)的草酸溶液為底物進行催化反應,在室溫下孵育10 min后加入顯色劑1-甲氧基-5-甲基-吩嗪硫酸甲酯和四唑硝基藍。二硫化四唑鹽被用作NADH的顯色指示劑,如果PnAAE41蛋白能以測試的有機酸作為底物,那么隨著NADH的消耗其顏色會從紫色變為淡黃色[20],基于此,可以作為定性研究的基礎。
2? 結果與分析
2.1? PnAAE41基因的鑒定與克隆
根據Pfam和SMART數據庫結構域預測共獲得49條AAE同源基因,根據植物基因的命名方法將其命名為PnAAE1~PnAAE49,其中39條含有完整的開放閱讀框(open reading frame,ORF)。以4條已驗證的具有草酰輔酶A合成酶功能的基因為參考序列[13-16],利用IQ-TREE軟件將其與49條三七AAE同源基因構建系統發育樹,結果顯示PnAAE41與已驗證有功能的基因聚為一支,其中PnAAE41與AtAAE親緣關系最近,與OsAAE親緣關系較遠;單子葉和雙子葉的AAE蛋白被分成兩個不同的分支蛋白被分成2個不同的分支(圖1)。為了進一步揭示PnAAE41是否具有AAE型基因的典型結構特征,將其與上述4條參考序列進行了多序列比對,結果顯示,它們具有較高的序列相似性;其中PnAAE41具有典型的AMP結合域和乙酰輔酶A合成酶結構域,并且在結構域及其附近具有高度的保守性(圖2)。
以33年生葉的cDNA為模板,利用特異性引物AAE41-F1和AAE41-R1經RT-PCR擴增與單克隆測序分析,序列結果顯示,其大小與預期結果一致,長1572 bp,編碼523個氨基酸。
2.2? PnAAE41的生物信息學分析
理化性質分析結果顯示,PnAAE41蛋白相對分子質量56.32 kDa,等電點6.14,不穩定指數33.51,平均親水性–0.024,屬于親水性蛋白。信號肽預測結果顯示,整個氨基酸肽鏈上沒有信號肽的存在,推測該蛋白屬于非分泌蛋白(圖3)。亞細胞定位預測結果顯示,PnAAE41蛋白具有多個亞細胞定位,其主要分布于細胞質膜。跨膜結構預測結果顯示,在第50~100個氨基酸之間有2個跨膜螺旋區,推測該蛋白允許存在跨膜結構域及其位點呈現(圖4)。
2.3? PnAAE41蛋白的二級與三級結構分析
PnAAE41蛋白的二級結構進行預測結果顯示(圖5),PnAAE41蛋白主要以無規則卷曲和α螺旋為主,其中無規則卷曲的結構類型所占比例最高,為47.99%;α螺旋的結構類型比例為35.56%;延伸鏈所占比例最低,為16.44%。
通過SWISS-MODEL在線軟件預測PnAAE41蛋白的三維結構并構建其三維結構模型,以草酰輔酶A連接酶[5ie2.1.A]為參考模型,結果顯示(圖6),與模板蛋白序列的一致性為79.69 %,GMQE值為0.93,QMEAN為0.51。
2.4? PnAAE41基因的時空表達特異性
以Actin為內參基因,對PnAAE41基因在三七不同生長年限、不同組織的相對表達水平進行了分析。結果顯示(圖7),PnAAE41基因在1年生三七葉中的相對水平最低,在根和莖中的相對表達水平較高,在22年生三七中,根中的表達水平較高,其次為莖,其中葉中表達水平最低。在33年生三七中,PnAAE41在莖中的表達水平高于根和葉。
2.5? 重組蛋白表達與催化活力分析
對測序驗證的陽性菌株進行了表達條件的篩選,當添加終濃度為0.5 mmol/L IPTG并在16 ℃、200 r/min下誘導9 h后,收集菌體并對其進行破碎、純化、SDS-PAGE檢測,結果發現,在此條件下誘導表達的蛋白量最多,其分子質量約56 kDa,與預期的重組蛋白大小一致(56.32 kDa),其中上清中出現大小一致的蛋白特異條帶(圖8~圖9)。使用BCA蛋白質定量試劑盒對濃縮后重組蛋白的濃度進行了測定(479 μg/mL),并利用顏色可視化法來檢測其對草酸的催化性,結果顯示,PnAAE41蛋白催化草酸消耗了NADH,顏色從紫色變成淡黃色(圖10)。
3? 討論
三七素屬于非蛋白質氨基酸,可通過增加體內血小板數量并縮短凝血和出血的時間達到止血效果,被廣泛用于治療外傷出血,是云南白藥等
多種傳統藥方的有效成分。鄭鄧毅男等[21]對三種人參屬對3種人參屬(三七、人參、西洋參)植物都進行了三七素含量檢測,結果發現三七素的含量以三七最高,人參次之,西洋參最低。三七素可能是人參屬植物特征性成分,該化合物容易被檢測到,因此可以作為人參屬植物化學分類的依據。前人對于三七素的研究僅涉及三七素液相質譜檢測與提取方法,并且多集中在豆科植物家山黧豆中[22-26]。
關于草酰輔酶A合成酶在植物中代謝途徑的提出可以追溯到1961年[27],但直到2012年這種酶才被發現。Foster等[22]描述了擬南芥酰基激活酶(AAE)基因,該基因編碼1種草酰輔酶A合成酶,能夠催化草酸分解代謝途徑的第一步,接著在蒺藜苜蓿、水稻、赤小豆與酵母中也發現了草酰輔酶A合成酶。在三七素合成途徑中,草酰輔酶A在最后一步中作為酰基供體進行酰基轉移反應,與L-α,β-二氨基丙酸在BAHD酰基轉移酶作用下產生三七素,這是最為關鍵的一步反應。我們基于三七轉錄組數據篩選到了1條草酰輔酶A合成酶基因PnAAE41并成功克隆,隨后對其理化性質、表達特征進行了分析。與已知的4種草酰輔酶A合成酶進行氨基酸序列比對發現PnAAE41具有AAE型基因典型的AMP結合域和乙酰輔酶A合成酶結構域,與擬南芥AtAAE具有86.05%的一致性。接著為了確定PnAAE41是否編碼草酰輔酶A合成酶,構建了該蛋白的His標記載體并在大腸桿菌中進行了異源表達,接著通過親和層析對獲得的蛋白進行純化并利用顏色反應檢測其催化功能。
AAE在草酸降解途徑中的作用是一個復雜的多信號互作過程,除了遺傳因素,還受到生長階段,組織部位等因素的影響。通過研究PnAAE41基因的組織時空特異性,發現該基因主要在三七根和莖中表達,與段紹鳳等[28]在研究三七素在三七植株的分布規律結果一致。本研究基于課題組前期對三七素的生物合成途徑做出的有根據的推測,成功克隆了候選基因PnAAE41,為深入研究三七素的生物合成提供了理論基礎。下一步將開展三七素合成途徑中所參與的BAHD酰基轉移酶基因的異源表達及功能分析,繼續完善三七中三七素的生物合成途徑,為三七素生物合成途徑的闡明提供參考。
參考文獻
[1]? Kosuge T, Yokota M, Ochiai A. Studies on antihemorrhagic principles in the crude drugs for hemostatics. II. Onantihemorrhagic principle in Sanchi Ginseng Radix [J]. Yakugaku Zasshi, 1981, 101(7): 629-632.
[2]? 趙國強, 王秀訓. 三七止血成分dencichine [J]. 中草藥, 1986, 17(6): 34-35, 20.
[3]? 焦成瑾, 楊玲娟, 趙菲佚, 等. 三七素檢測方法研究進展[J]. 氨基酸和生物資源, 2015, 37(3): 15-20.
[4]? Koh H L, Lau A J, Chan E C. Hydrophilic interaction liquid chromatography with tandem mass spectrometry for the determination of underivatized dencichine (beta-N-oxalyl- L-alpha, beta-diaminopropionic acid) in Panax medicinal plant species[J]. Rapid Commun Mass Spectrom, 2005, 19(10): 1237-1244.
[5]? Kuo Y H, Ikegami F, Lambein F. Neuroactive and other free amino acids in seed and young plants of Panax ginseng[J]. Phytochemistry, 2003, 62(7): 1087-1091.
[6]? Qiao C F, Liu X M, Cui X M. High-performance anion-exchange chromatography coupled with diode array detection for the determination of dencichine in Panax notoginseng and related species[J]. Journal of separation Separation scienceScience, 2013, 36(15): 2401-2406.
[7]? Malathi K, Padmanaban G, Sarma P S. Biosynthesis of β-N-oxalyl-l-α,β- diaminopropionic acid, the Lathyrus sativus neurotoxin[J]. Phytochemistry, 1970, 9(7): 1603-1610.
[8]? Martin P Peter E. Genetic improvement of grass pea (Lathyrus sativus) for low β-L-ODAP content[D]. Norwich: University of East Anglia, 2017.
[9]? Staswick P E, Tiryaki I, Rowe M L. Jasmonate response locus JAR1 and several related Arabidopsis genes encode enzymes of the firefly luciferase superfamily that show activity on jasmonic, salicylic, and indole-3-acetic acids in an assay for adenylation[J]. Plant Cell, 2002, 14(6): 1405-1415.
[10]????? Jay M, Shockey, Martin S, et al. Arabidopsis contains a large superfamily of acyl-activating enzymes. phylogenetic and biochemical analysis reveals a new class of Acyl-Coenzyme A Synthetases [J]. Plant Physiology, 2003, 132(2): 1065-1076.
[11]????? [11]Nakata P A. Plant calcium oxalate crystal formation, function, and its impact on human health[J]. Front Biology, 2012, 7(3): 254-266.
Foster J, Kim H U, Nakata P A, et al. A previously un-known oxalyl-CoA synthetase is important for oxalate ca-tabolism in Arabidopsis[J]. Plant Cell, 2012, 24(3): 1217-1229.
[12]蔡曉鋒, 徐晨曦, 王小麗. 植物中的草酸: 合成、降解及其積累調控[J]. 植物生理學報, 2015, 51(3): 267-272.
[11]
[12]????? 蔡曉鋒, 徐晨曦, 王小麗. 植物中的草酸: 合成、降解及其積累調控[J]. 植物生理學報, 2015, 51(3): 267-272.
Foster J, Kim H U, Nakata P A, et al. A previously unknown oxalyl-CoA synthetase is important for oxalate catabolism in Arabidopsis[J]. Plant Cell, 2012, 24(3): 1217-1229.
[12]????? 272.
[13]????? Foster J, Kim H U, Nakata P A, et al. A previously un-known oxalyl-CoA synthetase is important for oxalate ca-tabolism in Arabidopsis[J]. Plant Cell, 2012, 24(3): 1217-1229.
[13]????? Foster J, Luo B, Nakata P A. An oxalyl-CoA dependent pathway of oxalate catabolism plays a role in regulating calcium oxalate crystal accumulation and defending against oxalate-secreting phytopathogens in Medicago truncatula[J]. PLoS One, 2016, 11(2): e0149850.
[14]????? Foster J, Luo B, Nakata P A. An oxalyl-CoA dependent pathway of oxalate catabolism plays a role in regulating calcium oxalate crystal accumulation and defending against oxalate-secreting phytopathogens in Medicago truncatula[J]. PLoS One, 2016, 11(2): e0149850.
[14]????? Lou H Q, Fan W, Xu J M, et al. An oxalyl-CoA synthetase is involved in oxalate degradation and aluminum tolerance[J]. Plant Physiology, 2016, 172(3): 1679-1690.
[15]????? Lou H Q, Fan W, Xu J M, et al. An oxalyl-CoA synthetase is involved in oxalate degradation and aluminum tol-erance[J]. Plant Physiology, 2016, 172(3): 1679-1690.
[15]????? Foster J, Nakata P A. An oxalyl-CoA synthetase is im-portant for oxalate metabolism in Saccharomyces cerevisiae [J]. FEBS Letters, 2014, 588(1): 160-166.
[16]????? Foster J, Nakata P A. An oxalyl-CoA synthetase is im-portant for oxalate metabolism in Saccharomyces cerevisiae [J]. FEBS Letters, 2014, 588(1): 160-166.
[16]????? Chen H, Kim H U, Weng H, et al. Malonyl-CoA synthetase, encoded by ACYL ACTIVATING ENZYME13, is essential for growth and development of Arabidopsis[J]. Plant Cell, 2011, 23(6): 2247-2262.
[17]????? Chen C, Chen H, Zhang Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data[J]. Molecular Plant, 2020, 13(8): 1194-1202.
[18]????? Li X J, Yang J L, Hao B, et al. Comparative transcriptome and metabolome analyses provide new insights into the molecular mechanisms underlying taproot thickening in Panax notoginseng[J]. BMC Plant Biology, 2019, 19(1): 451.
[19]????? Livak K J, Schmittgen T D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method[J]. Methods, 2001, 25(4): 402-408.
[20]????? Ishiyama M, Miyazono Y, Sasamoto K, et al. A highly water-soluble disulfonated tetrazolium salt as a chromogenic indicator for NADH as well as cell viability[J]. Talanta, 1997, 44(7): 1299-1305.
[21]????? 鄭毅男,李向高. 人參屬植物止血成分比較分析[J]. 吉林農業大學學報, 1989, 11(1): 24–-27.
[22]????? Foster J, Kim H U, Nakata P A, et al. A previously unknown oxalyl-CoA synthetase is important for oxalate catabolism in Arabidopsis[J]. Plant Cell, 2012, 24(3): 1217-1229.
[23]????? Xie G X, Qiu Y P, Qiu M F, et al. Analysis of dencichine in Panax notoginseng by gas chromatography-mass spectrometry with ethyl chloroformate derivatization[J]. J Pharm Biomed Anal, 2007, 43(3): 920-925.
[24]????? 李? 琳, 王承瀟, 曲? 媛, 等. 反相離子對色譜法測定三七中的三七素含量[J]. 中國中藥雜志, 2015, 40(20): 4026-4030.
[25]????? 楊玲娟, 焦成瑾, 高二全. 三七中三七素及其異構體的高效液相色譜檢測[J]. 中藥材, 2015, 38(2): 311-314.
[26]????? Ji W H, Xie H K, Zhou J, et al. Water-compatible molecularly imprinted polymers for selective solid phase extraction of dencichine from the aqueous extract of Panax notoginseng[J]. Journal of Chromatography B Analytical Technologies in the Biomedical and Life Sciences, 2016, 1008: 225-233.
[27]????? Giovanelli J, Tobin N F. Adenosine triphosphate-and coenzyme A-dependent decarboxylation of oxalate by extracts of peas[J]. Nature, 1961, 190: 1006-1007.
[28]????? 段紹鳳, 陳? 庚, 閆? 靜, 等. 三七素提取工藝及檢測方法的優化研究[J]. 熱帶作物學報, 2019, 40(11): 2255-2260.
責任編輯:黃東杰
