黔西北黃杉群落物種多樣性的海拔梯度格局
2021-09-09 04:47:10何斌李青陳群利李望軍游萍
生態環境學報 2021年6期
何斌,李青,陳群利,李望軍,游萍
貴州省普通高等學校生物資源開發與生態修復特色重點實驗室/貴州工程應用技術學院生態工程學院,貴州 畢節 551700
物種多樣性是研究植物群落結構與功能的物質基礎(Curtis et al.,1951;張曉龍等,2017),不僅能夠反映植物群落對生態環境的響應,而且可以有效評價生態系統的穩定性(劉維暐等,2012)和健康狀況(Rad et al.,2009)。物種多樣性的空間分布格局及其潛在機制一直是生態學和植物地理學研究的熱點問題之一(Whittaker,1960;Macarthur,1972),受多個生態梯度(如經緯度、海拔、研究尺度等)的影響(Palmer,1992;黃建輝,1994),其中海拔梯度被認為是一個關鍵性的影響因子(Debano et al.,1990;Zimmerman et al,1999)。因此,在全球氣候變暖的背景下,加強物種多樣性的海拔梯度格局研究,對于揭示物種多樣性的環境梯度變化規律具有重要意義(郝建鋒等,2014)。
植物群落物種多樣性的海拔梯度格局常常與研究區的地理位置、環境條件、研究對象和研究尺度等有關(Ghimire et al.,2006;方精云等,2004;吳昊等,2012)。已有研究表明,不同地區山地物種多樣性的海拔梯度格局不同,熱帶地區以單調下降趨勢為主,亞熱帶地區以先上升再下降趨勢為主,溫帶地區變化趨勢不確定(Ohsawa,1991;Ohsawa,1995;Liu et al.,2007;方精云,2004;沈澤昊等,2004;王志恒等,2004)。低海拔地區降水和人為干擾是限制物種多樣性的關鍵因子(Wang et al.,2007),高海拔地區則為溫度(Whittaker et al.,1975)。Beck et al.(2008)認為山地水熱條件的綜合效應是造成物種多樣性單峰格局的主要原因,而Begon et al.(1986)認為溫度是造成物種多樣性隨海拔梯度單調下降的主要原因;Lomolino(2001)和Rahbek(2005)認為物種多樣性的海拔梯度格局存在不確定性,取樣時的最低海拔梯度和不同海拔梯度的地形變化均可導致物種多樣性單調下降的海拔梯度格局現象;Wu et al.(2014)研究認為進化因素也是造成物種多樣性海拔格局的因素之一。由此可見,目前有關山地物種多樣性的海拔梯度格局以及機制還沒有形成統一的定論。因此,開展不同植物類群、不同山體和不同區域物種多樣性海拔格局的案例研究,對闡明物種多樣性與海拔之間的關系具有重要意義,有利于更好地理解山地植物對環境變化的適應(Kitayama,1992;Grytnes et al.,2002)。
隨著全球經濟活動的加劇,人類對植物資源的過渡利用和生態環境的破壞,導致大量物種瀕于滅絕,特別是近幾十年來,許多瀕危物種的分布區迅速縮小。在這種背景下,瀕危物種多樣性的研究得到了眾多生態學家的關注,并成為重點研究對象。黃杉(Pseudotsuga sinensis)屬松科(Pinaceae)黃杉屬(Pseudotsuga),為第三紀孑遺植物,中國特有種,被列為國家Ⅱ級重點保護植物,在全國分布范圍狹窄,僅在貴州、滇東北、滇中、川東、川西南、湘西、鄂西等地呈零星分布(熊斌梅等,2017)。在貴州省威寧縣分布的黃杉純林,面積約373.2 hm2,是國內罕見的保存比較完整、面積較大的黃杉群落。由于黃杉林是黔西北地區喀斯特山地森林生態系統的一個重要組成部分,研究黃杉群落物種多樣的垂直分布格局,不僅對探討其瀕危機制及生物多樣性保護具有重要的意義,而且有利于揭示亞熱帶喀斯特山地森林生態系統的結構、演替和功能。目前,有關黃杉的研究較少,且主要集中在群落結構(左家哺,1995;熊斌梅等,2017)、種群動態(孟廣濤等,2008)和生態評價(胡文彬,2015)等方面。基于此,本文以黃杉群落為研究對象,通過典型群落樣地調查,分析群落物種組成、植物區系成分、物種多樣性對海拔變化的響應,旨在探討,(1)植物區系成分和物種多樣性的海拔梯度格局?(2)物種α多樣性各指數間是否存在相關性?(3)灌木層和草本層的植物多樣性格局是否一致?以期為喀斯特山地生物多樣性保護和瀕危植物的保育提供參考,也為研究喀斯特山地森林植物群落的穩定性機制奠定基礎。
1 研究區概況與方法
1.1 研究區概況
研究區位于貴州省威寧縣西南部的哲覺、黑石、麻乍等6個鄉鎮,屬中亞熱帶季風氣候,年均溫10.5—3.5 ℃,年均降雨量1000 mm左右,年總日照時數1530—1926 h。地形復雜、地勢起伏大,坡度10°—30°之間,土壤母質以紫色砂巖和砂頁巖為主,主要為黃壤、黃棕壤和紫色土,呈酸性(pH 5.5—6.0)。研究區內植被類型主要有黃杉林、云南松(Pinus yunnanensis)林、華山松(P. armandii)林、金絲桃(Hypericum monogynum)灌叢、馬桑(Coriaria nepalensis)灌叢、西南栒子(Cotoneaster franchetii)灌叢、滇榛(Corylus yunnanensis)灌叢、莢蒾(Viburnum dilatatum)灌叢和荒山草坡。
研究區內黃杉林分在20世紀50年代到80年代期間被當地群眾大肆砍伐,林地面積減少了88.95%、蓄積減少了89.71%(黃鶴先,1989)。現有林分中,黃杉以中、幼林分居多,根據野外調查,主要有以下4種類型:黃杉純林;黃杉、云南松混交林;黃杉、華山松混交林和黃杉散生木,且大部分為天然更新。
1.2 樣地設置與調查
在對黃杉分布區全面踏查的基礎上,結合實際情況,采用典型樣地法于2019年7—10月從海拔1800 m的金斗鄉到2290 m的哲覺鎮,分別在6個海拔梯度上選擇有代表性的黃杉林設置20 m×30 m的樣地,每個海拔梯度2個重復,共12塊樣地,對喬木層植物進行每木檢尺,同時記錄樣地的海拔、經緯度、坡度、地形等基本信息(表1)。在每個樣地內,按梅花形選取5個5 m×5 m的灌木層樣方和1 m×1 m的草本層樣方,分別調查灌木植物的種類、株數(叢數)、高度和冠幅和草本植物的種類、株數(叢數)、平均高度和蓋度。
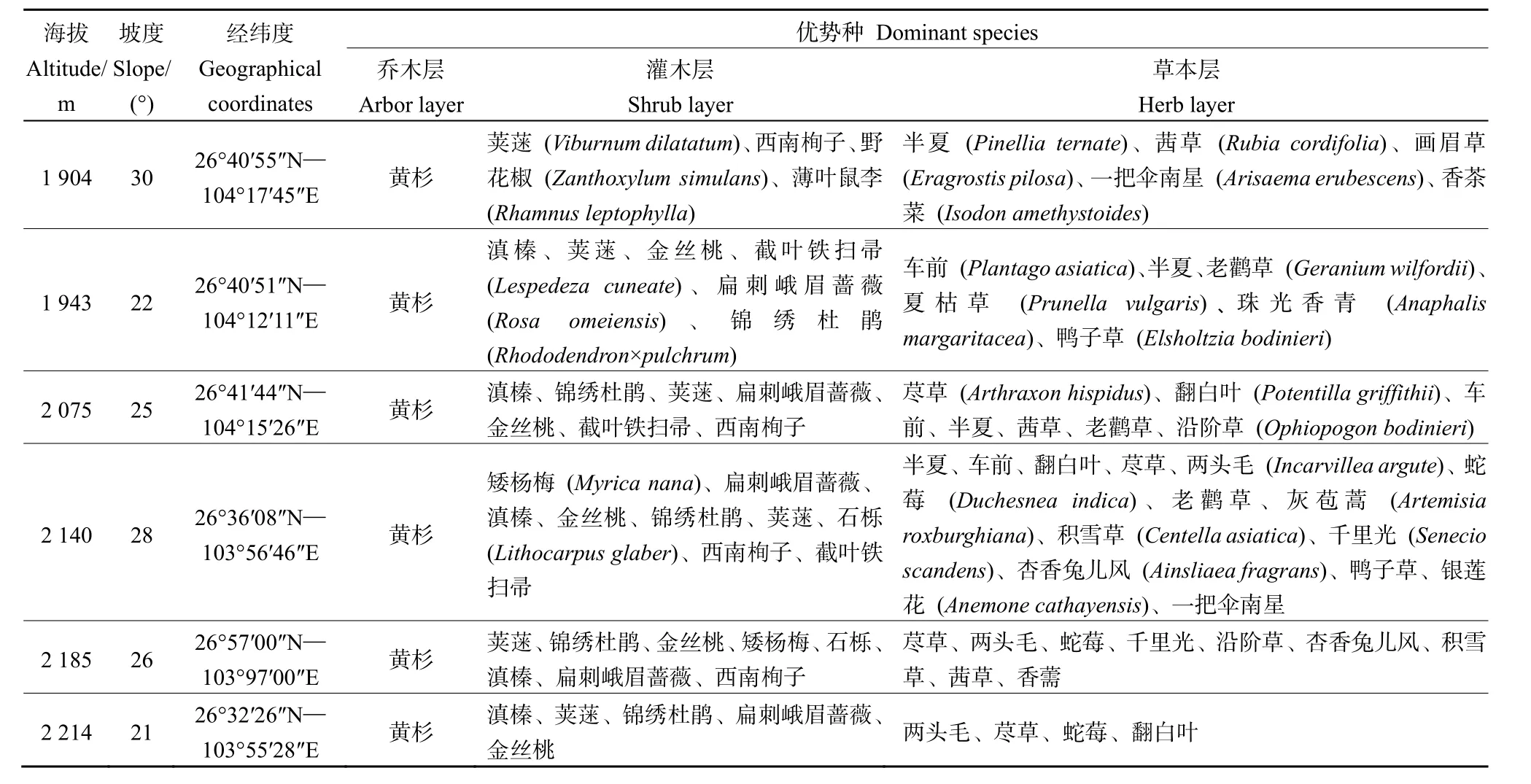
表1 研究區內黃杉林樣地概況Table 1 General characteristics of the sample plot of P. sinensis forest in the study area
1.3 植物區系分析
參照《世界種子植物科的分布區類型系統》(吳征鎰等,2003)和《中國種子植物屬的分布區類型》(吳征鎰,1991)對中國種子植物分布區類型的劃分,進行統計分析。
1.4 物種多樣性分析
參考馬克平等(1995)和方精云等(2004)的生物多樣性測度方法,本文從α多樣性和β多樣性兩個方面分析海拔梯度對物種多樣性的影響。
灌木層和草本層重要值(IV)計算公式為:

式中:
Hr為相對高度;
Cr為相對蓋度;
Fr為相對頻度。
(1)α多樣性指數:
物種豐富度指數(S):

式中:
N為樣地內出現的物種數。
Shannon-Wiener多樣性指數:

Simpson優勢度指數:

Pielou均勻度指數:

式中:
Pi為物種i的重要值。
(2)β多樣性指數:
S?renson物種相似性指數(Cs):

Cody物種替代指數(βc):

式中:
a、b分別為任意兩個海拔梯度的物種數;
c為兩海拔梯度間的共有物種數。
1.5 數據分析
運用SPSS 19.0統計軟件進行以下分析:(1)單因素方差分析(One-Way ANOVA),以海拔梯度為自變量,以不同層次的物種多樣性指數為因變量進行方差分析和顯著性檢驗;(2)黃杉群落不同層次物種多樣性的 Pearson相關性分析。應用Microsoft Excel 2016進行繪圖。
2 結果與分析
2.1 植物群落的物種組成
本次調查,樣地中共記錄種子植物50科95屬124種。由表2可知,含有10個以上物種的科有2科,占總物種數比例為 23.39%,分別為薔薇科(Rosaceae)16種和菊科(Compositae)13種;含5—9個物種的科共有 5科,占總物種數的比例為25.81%,分別為杜鵑花科(Ericaceae)8種、禾本科(Gramineae)7種、殼斗科(Fagaceae)6種、豆科(Leguminosae)6種和唇形科(Labiatae)5種。上述7個科僅占總科數的14%,但是所包括的物種占總物種數的49.2%,說明這7個科的植物在黃杉群落區系組成中起主導作用。含4個物種及以下的科有43科,其中31科只含有1個物種,占總物種數的50.8%,但是占總科數的86%。
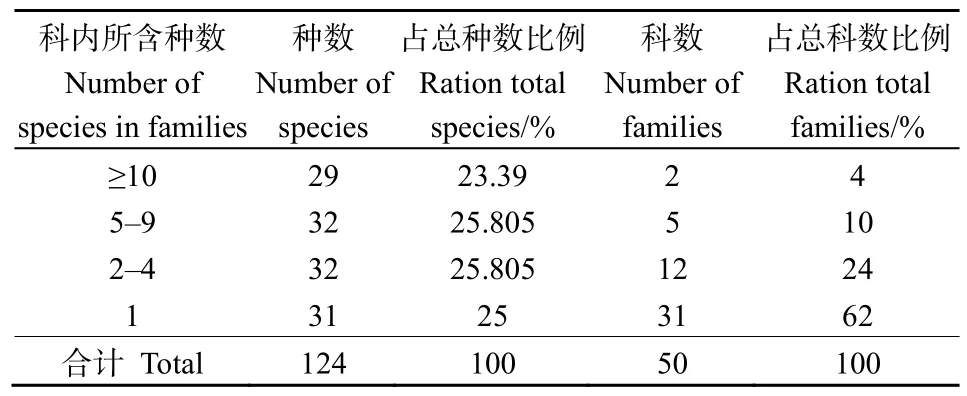
表2 黃杉群落種子植物科內種的組成Table 2 Composition of species in families of seed plants in the P. sinensis community
由表3可知,群落內各科所含屬數差異也很明顯。含有10屬以上的科只有1科,為菊科,占總科數的2%,占總屬數的11.58%;含有4—9屬的科共有5科,占總科數的10%,占總屬數的30.53%,分別為薔薇科9屬、禾本科6屬、豆科5屬、唇形科5屬和樺木科4屬;含3屬及以下的科共有44科,占總科樹的88%,占總屬數的57.89%,其中33科只有1屬。
基于語文核心素養要求的作文自改教學是一種行之有效的教學方法,對教學策略的探索也是有益的,但本文中的策略設計還不夠完美,對實際的教學問題的考慮也還不夠周全,希望能夠在日后的探索中精益求精,盡善盡美。我相信,只要方向是正確的,即便此時仍舊行走在羊腸小道,只要堅持開拓,前方終會迎來坦途。
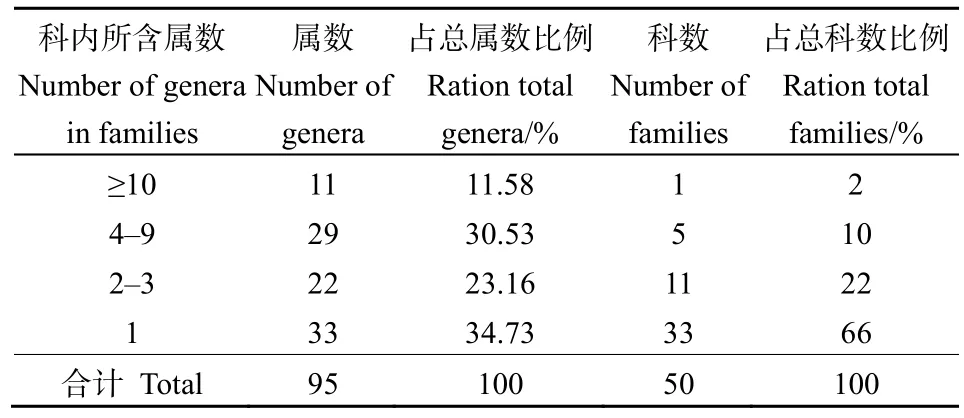
表3 黃杉群落種子植物科內屬的組成Table 3 Composition of genera in families of seed plants in the P. sinensis community
2.2 植物區系成分
由表4可知,黃杉群落內種子植物科的地理成分較少,僅有6種分布區類型,其中世界分布最多,有19科,占總科數的38%,有禾本科、薔薇科、菊科和唇形科等。熱帶地理成分共有20科,占總科數的40%,包括泛熱帶分布16科、東亞(熱帶、亞熱帶)及熱帶南美間斷分布2科、熱帶亞洲至熱帶非洲分布1科和熱帶亞洲(印度—馬來西亞)分布1科。溫帶地理成分中僅有北溫帶分布,共有11科,占總科數的22%。
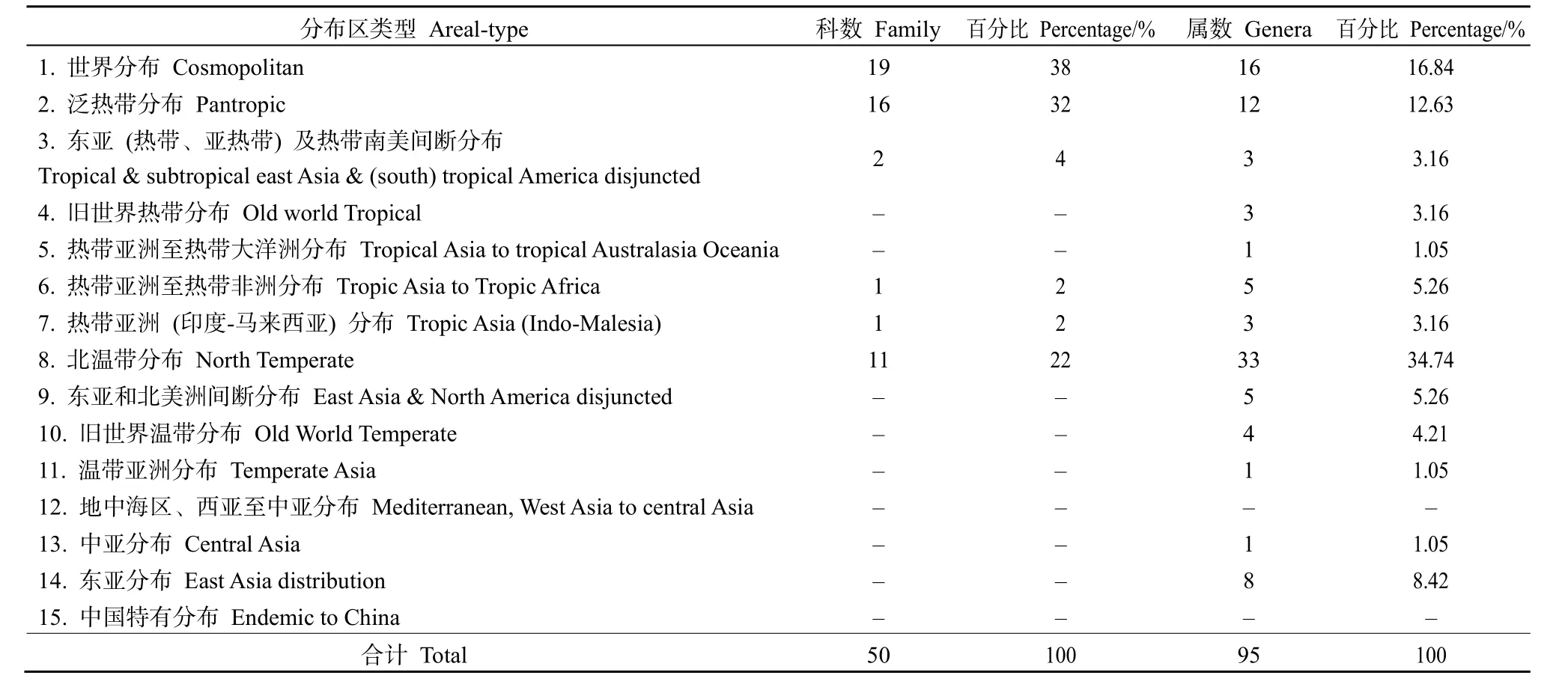
表4 種子植物科、屬的分布區類型Table 4 The distribution type of families and genera in the flora of seed plants
在 15種分布區類型中,屬的地理成分僅缺少了地中海區、西亞至中亞分布和中國特有分布,表明黃杉群落種子植物屬的分布區類型復雜、多樣。其中,北溫帶分布、世界分布和泛熱帶分布占據了群落的主體。世界分布型共有 16屬,占總屬數的16.84%,有金絲桃屬(Hypericum)、懸鉤子屬(Rubus)、車前屬(Plantago)和飛蓬屬(Erigeron)等。熱帶地理成分共有27屬,占總屬數的28.42%。其中,泛熱帶分布(12屬)占優勢,占熱帶成分的44.44%,有醉魚草屬(Buddleja)、花椒屬(Zanthoxylum)和薯蕷屬(Dioscorea)等;熱帶亞洲至熱帶非洲分布有5屬,占熱帶成分的18.52%,有鐵仔屬(Myrsine)和香茶菜屬(Rabdosia)等;東亞(熱帶、亞熱帶)及熱帶南美間斷分布有3屬,占熱帶成分的 11.11%,有柃屬(Eurya)和葎草屬(Humulus)等;舊世界熱帶分布有3屬,占熱帶成分的11.11%,有烏蘞莓屬(Cayratia)和千金藤屬(Stephania)等;熱帶亞洲(印度-馬來西亞)分布有3屬,占熱帶成分的11.11%,有潤楠屬(Machilus)和木荷屬(Schima)等;熱帶亞洲至熱帶大洋洲分布有 1屬,占熱帶成分的 3.71%,有栝樓屬(Trichosanthes)。溫帶地理成分多于熱帶地理成分,共有52屬,占總屬數的54.74%。北溫帶分布占優勢,有 33屬,占總屬數的 34.74%,有莢蒾屬(Viburnum)、馬桑屬(Coriaria)、小檗屬(Berberis)和栒子屬(Cotoneaster)等;東亞分布有8屬,占總屬數的 8.42%,有櫟屬(Quercus)和半夏屬(Pinellia)等;東亞和北美洲間斷分布有 5屬,占總屬數的5.26%,有黃杉屬(Pseudotsuga)和胡枝子屬(Lespedeza)等;舊世界溫帶分布有4屬,占總屬數的4.21%,有火棘屬(Pyracantha)和香薷屬(Elsholtzia)等;溫帶亞洲分布有1屬,占總屬數的1.05%,有杭子梢屬(Campylotropis);中亞分布有1屬,占總屬數的1.05%,有角蒿屬(Incarvillea)。
2.3 物種多樣性的海拔梯度格局
由于喬木層基本上為黃杉,偶爾有零星華山松(P. armandii)、云南松(P. yunnanensis)、川滇榿木(Alnus ferdinandi-coburgii)和栓皮櫟(Quercus variabilis)分布,故下面主要分析灌木層和草本層物種多樣性的海拔梯度格局。
2.3.1 α多樣性的海拔梯度格局
從黃杉群落灌木層和草本層物種多樣性的海拔梯度變化趨勢圖(圖 1)中可以看出,隨著海拔的升高,灌木層和草本層的物種豐富度指數 S、Shannon-Wiener多樣性指數H、Simpson優勢度指數D和Pielou均勻度指數J均呈單峰格局。其中,物種豐富度指數S、Shannon-Wiener多樣性指數H和Simpson優勢度指數D變動幅度較大,均在海拔2140 m處達到最大值;Pielou均勻度指數J變化較平緩,灌木層在海拔1943 m處達到最大值,而草本層在海拔2075 m處達到最大值。

圖1 灌木層和草本層物種多樣性的海拔梯度變化格局Fig. 1 Altitudinal gradient pattern of species diversity in shrub layer and herb layer
從圖2中可以看出,不同海拔間,灌木層的物種豐富度、Shannon-Wiener多樣性指數H、Simpson優勢度指數D差異顯著(P<0.05),均在海拔2140 m處出現最大值和海拔 1904 m處出現最小值;Pielou均勻度指數J差異不顯著(P<0.05),最大值出現在海拔1943 m處,最小值出現在海拔2214 m處。不同海拔梯度間,草本層的物種豐富度、Shannon-Wiener多樣性指數H、Simpson優勢度指數D差異顯著(P<0.05),均在海拔2140 m處出現最大值和海拔2214 m處出現最小值;Pielou均勻度指數J差異也顯著(P<0.05),最大值出現在海拔2075 m處,最小值出現在海拔1904 m處。
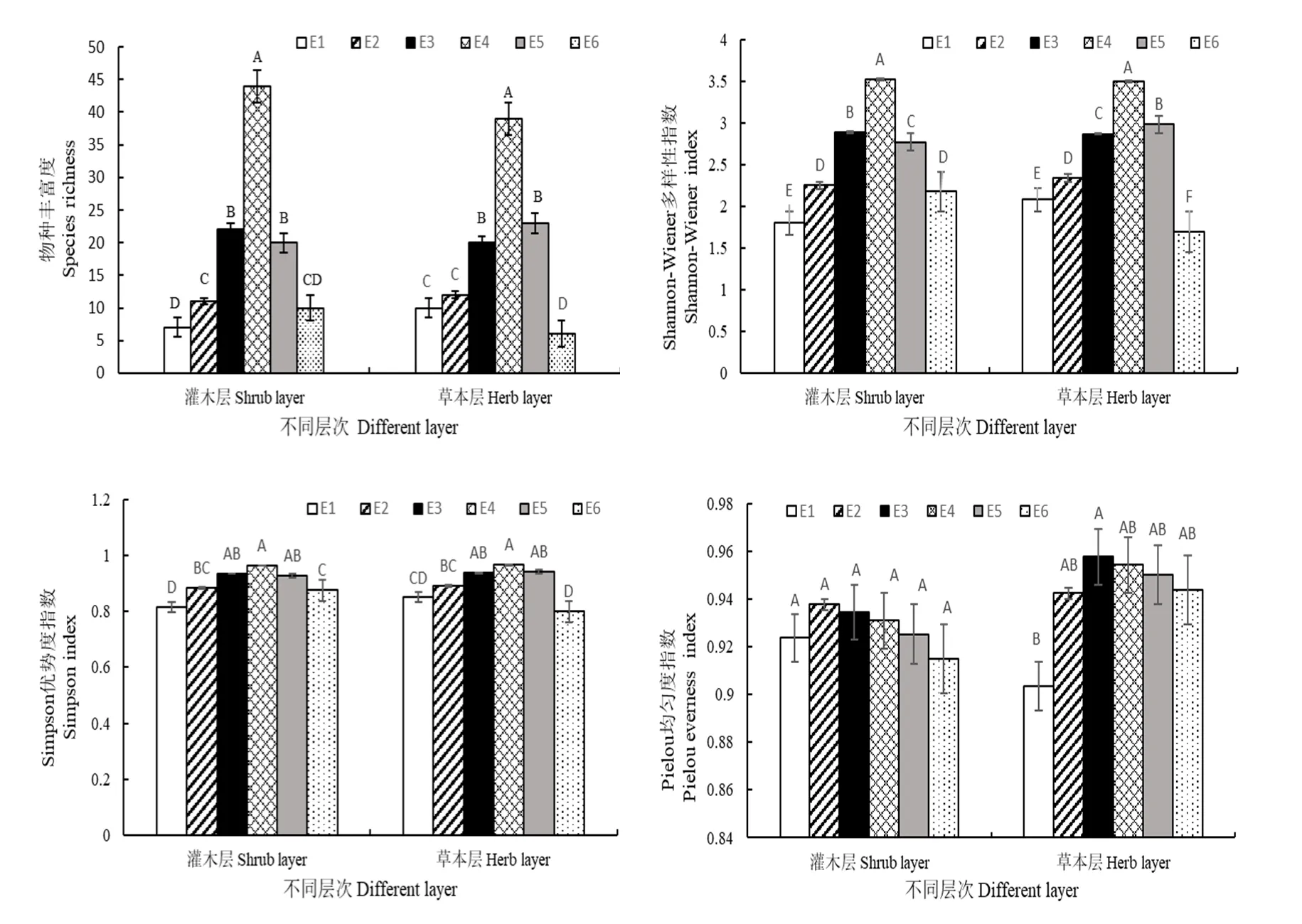
圖2 不同海拔灌木層和草本層物種多樣性的比較Fig. 2 Comparison of species diversity in shrub layer and herb layer at different elevation
2.3.2 不同海拔間α多樣性指數間關系
通過對黃杉群落物種α多樣性指數的相關性分析(表5)發現,灌木層和草本層的Shannon-Wiener多樣性指數H與物種豐富度指數S和Simpson優勢度指數D存在極顯著正相關性(P<0.01),物種豐富度指數S與Simpson優勢度指數D存在顯著正相關性(P<0.05),Pielou均勻度指數J與其他多樣性指數間相關性不顯著(P<0.05)。整體來看,草本層α多樣性指數間的相關性高于灌木層α多樣性指數間的相關性。

表5 植物群落α多樣性指數相關性分析Table 5 Correlation analysis of α diversity index of plant community
圖3為相鄰海拔梯度間的β多樣性指數變化趨勢,結果表明,隨著海拔的增加,灌木層的S?renson指數變化較大,呈雙峰曲線,分別在 1943—2075 m和 2140—2185 m處達到最大值;草本層的S?renson指數呈單峰曲線,在2140—2185 m處最大。隨著海拔的上升,灌木層和草本層的Cody指數均呈單峰變化趨勢,均在2075—2140 m達到最大值,但是灌木層的Cody指數變化幅度更大。上述分析表明,隨著海拔梯度的增加,灌木層間物種的變化比草本層間物種的變化大。
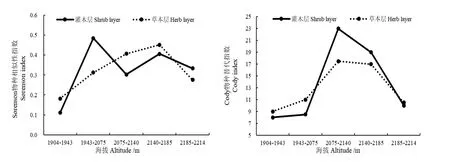
圖3 相鄰海拔梯度上灌木層和草本層的S?renson指數和Cody指數Fig. 3 The S?renson index and Cody index in shrub layer and herb layer of each two adjacent altitude gradient
由表 6可知,不同海拔梯度間灌木層物種的S?renson指數和Cody指數有一定的差異。不同海拔梯度上,灌木層植物 S?renson指數的平均值為0.3066,其中在海拔2075 m與海拔2214 m間最高,為 0.500;灌木層植物 Cody指數的平均值為13.2,其中在海拔2075 m和海拔2140 m間最高,為 23。但是,隨著海拔差的增加,灌木層植物的S?renson指數和Cody指數無明顯變化規律。

表6 不同海拔灌木層的S?renson指數和Cody指數Table 6 The S?renson index and Cody index of shrub layer in different elevations
由表 7可知,不同海拔梯度間草本層物種的S?renson指數和Cody指數也存在一定的差異。不同海拔梯度上,草本層植物S?renson指數的平均值為0.2018,其中在海拔2140 m與海拔2185 m間最高,為0.4516;隨著海拔差的增加,S?renson指數基本上呈逐漸下降的趨勢,甚至在海拔 2214 m與海拔1904 m和1943 m間以及海拔1943 m與海拔2185 m間S?renson指數為0。不同海拔梯度上,草本層植物Cody指數的平均值為14,其中在海拔2140 m與海拔1904 m間最高,為21.5;但是,隨著海拔差的增加,Cody指數無明顯變化規律。
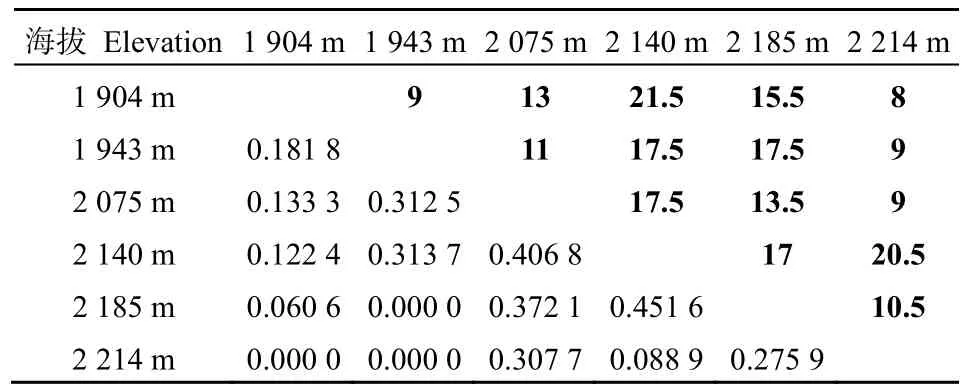
表7 不同海拔草本層的S?renson指數和Cody指數Table 7 The S?renson index and Cody index of herb layer in different elevations
3 討論
3.1 黃杉群落的植物地理成分
本研究調查表明,研究區內黃杉群落植物屬的地理成分復雜、多樣,北溫帶分布區類型占明顯優勢(34.74%),說明黃杉群落內植物區系具有明顯的溫帶性質,溫帶氣候對群落內植物的系統發育過程產生了嚴重的影響,與研究區內的氣候條件比較吻合,這與左家哺(1995)的研究結果基本一致。
3.2 海拔梯度對物種α多樣性的影響
在眾多生態因子中,海拔是山地環境中最為重要的因素之一,它可以通過影響太陽輻射、溫度、水分和土壤等資源的分配進而影響生態系統的生態水文過程,導致植物群落空間分布格局的不同(Gaston,2000;朱珣之等,2005;張樹斌等,2018)。許多研究表明,物種α多樣性的海拔梯度格局主要有單調下降、單調上升、先升高后降低、先降低后升高和無規律性5種類型,負相關和中間高度膨脹型是較為普遍的格局。本研究中,黃杉群落灌木層和草本層的物種豐富度指數S、Shannon-Wiener多樣性指數H、Simpson優勢度指數D和Pielou均勻度指數J均呈單峰格局變化趨勢,這與蘭斯安等(2016)、王飛等(2018)、吳紅寶等(2019)和蘇闖等(2018)的研究結果相似。這可能是由于以下原因造成的:首先,相對海拔是影響喀斯特山地水熱條件分布的主要因素,它通過影響光照、水分及土壤資源的再分配進而影響植物群叢的分布及結構,相對海拔較低的地帶,水分充足甚至過量,但光照條件較差,生長季節光照不足常常成為限制植物分布的主要因素;相對海拔較高的地帶,山體陡峭、土層較薄,氣溫劇烈變化產生了許多異質性的微生境,而且由于喀斯特地區特殊的雙層地質結構,水分會迅速從下層流失,形成了干旱缺水的嚴酷生境條件,不利于植物定居;中海拔地區,水分和熱量的組合、配比適宜,生境條件最好,資源的可利用率最高,優越的水熱條件能使更多的物種共存,物種多樣性達到最大。其次,人為干擾是另外一個原因,低海拔地區是村鎮的主要分布區域,人為干擾以及牛羊等家畜的踩踏、啃食現象嚴重,一些物種喪失了生存機會;隨著海拔的升高,人為干擾活動強度在逐漸降低,因干擾而缺失的部分物種得到自然釋放,重新分布(王宇超等,2013)。
王國宏(2002)認為,群落不同生活型植物(喬、灌、草植物)的物種多樣性對海拔梯度的響應不同。本研究結果表明,黃杉群落灌木層和草本層的物種α多樣性指數在不同海拔上存在顯著差異(P<0.05),但灌木層物種的Pielou均勻度指數不顯著(P≥0.05)。這與福建梅花山常綠闊葉林植物物種多樣性指數的特征相似(孔祥海等,2012)。這可能是由于環境條件(特別是水分和熱量)對不同生活型植物分布的影響不同,隨著海拔的變化,環境條件和資源配置發生了變化,導致了不同生活型植物的物種多樣性垂直分布存在差異(Bhattarai et al.,2003;Grytnes et al.,2006)。
3.3 海拔梯度對物種β多樣性的影響
本研究中,黃杉群落灌木層和草本層的物種 β多樣性指數在相鄰兩個海拔梯度間變化很大,變化趨勢不同(圖3);灌木層物種S?renson指數和Cody指數的變化幅度均比草本層大(表 6、7)。這表明研究區內灌木層物種組成在海拔梯度上差異更大,物種更替速率更快,可能與不同生活型植物的生活史策略和對環境響應不同有關(秦浩等,2019)。首先,木本植物在系統進化方面更加保守,其分布主要受歷史過程的影響,表現為親緣關系較近的物種共存;草本植物則表現為親緣關系較遠的物種共存(秦浩等,2019)。其次,木本植物受溫度的影響較大(Pausas et al.,2001),木本植物的β多樣性對環境差異具有相對較強的響應;草本植物受光照、水分、微地形、土壤理化性質和人為活動等影響更大(Qian et al.,2014),而且其它因子的修飾作用會削弱或加強特定生境的海拔梯度效應,草本植物的 β多樣性受環境差異和地理距離的共同影響。
目前,不同學者對植物群落物種β多樣性海拔梯度格局的結論不同。最普遍的格局為:隨海拔高差的增加,物種β多樣性指數減小。本研究表明,隨著海拔高差的增加,草本層物種的S?renson指數符合這種格局,但是,灌木層物種的S?renson指數和 Cody指數以及草本層物種的 Cody指數均無明顯變化規律。這種無明顯變化規律的格局可能是由以下原因造成的:一是研究區屬于喀斯特山地,黃杉群落樣地的生境異質性較高,但是異質性的增加與距離的增加并不完全一致;二是世界分布種的存在可能降低了擴散限制作用(林國俊等,2010)。
綜上所訴,本研究對黃杉群落多樣性的海拔梯度格局進行了探討,對保護黃杉種質資源、維持其持續更新具有重要意義。但是,研究中只對海拔梯度這一綜合因子對黃杉群落物種多樣性的影響進行了探討,沒有進一步深入研究水分、溫度、土壤和微生物等環境因子的海拔梯度變化、對黃杉群落生物多樣性的影響以及與黃杉生長發育之間的關系,以后應加強相關方面的研究。
4 結論
(1)隨著海拔的升高,黃杉群落灌木層和草本層物種的α多樣性指數均呈單峰格局;不同海拔上,灌木層和草本層的物種α多樣性指數均存在顯著差異(P<0.05),灌木層物種的Pielou均勻度指數除外。
(2)隨著海拔的增加,不同生活型植物的β多樣性表現出不同的變化趨勢,灌木層物種組成沿海拔梯度的變化更劇烈。隨著海拔高差的增加,除草本層物種的S?renson指數逐漸減小外,其他β多樣性指數變化規律不明顯。
(3)相對海拔導致的水熱條件差異是影響喀斯特山地物種多樣性垂直分布格局的主要因素,其中,中海拔地帶黃杉群落的物種多樣性最高,這對黃杉種質資源的保護和維持具有重要意義。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
