亞熱帶針闊混交林土壤-植物-大氣連續體(SPAC)中水穩定同位素特征
2021-09-09 04:47:16王銳章新平戴軍杰羅紫東賀新光關華德
生態環境學報 2021年6期
關鍵詞:植物
王銳,章新平, ,戴軍杰,羅紫東,賀新光, ,關華德
1. 湖南師范大學資源與環境科學學院,湖南 長沙 410081;2. 湖南師范大學/地理空間大數據挖掘與應用湖南省重點實驗室,湖南 長沙 410081;3. National Centre for Groundwater Research and Training, Flinders University, Adelaide, SA 5001, Australia
森林生態系統是陸地水分調節和物質交換的重要場所,對于維護區域生態平衡和環境保護具有重要的意義(婁源海等,2016;李龍等,2020)。大氣降水作為水分的重要輸入項對整個系統水分補給過程具有重要影響,而土壤中儲存的水分又通過蒸發、下滲和植物蒸騰等方式進行重新分配,進而實現區域森林水循環過程的動態平衡(Luo et al.,2019;Dai et al.,2020;鄧文平等,2017)。近年來,隨著穩定同位素技術的逐漸成熟,利用穩定同位素示蹤的方法可從微觀上提取不同水體的水同位素信息,準確地揭示水分在區域森林生態系統中的運移規律(Dawson et al.,2002;Robertson et al.,2006)。相關研究表明,一個地區降水中穩定同位素變化特征與區域氣候特征緊密相關,具體表現在降水、溫度、濕度、風向風速、水汽輸送過程以及云下二次蒸發方面的特征(吳華武等,2012;黃一民等,2013;章新平等,2015;周慧等,2018)。受降水補給的影響,土壤水很大程度上保留了降水中穩定同位素信息。與此同時,降水在入滲過程中受蒸發和前期土壤水分混合的影響,穩定同位素值發生明顯變化,這種變化隨深度增加逐漸減小(王銳等,2020a)。
植物作為聯系土壤和大氣的介質,是區域森林系統水循環過程中最重要的環節。了解植物水中氫氧穩定同位素變化有助于揭示其在局地水體氫氧穩定同位素循環中的貢獻(Snyer et al.,2010;Stahl et al.,2013;羅倫等,2013)。研究發現,除少數鹽生或旱生植物外,大部分植物在蒸騰之前體內水中穩定同位素不發生分餾,當各水源中穩定同位素存在差異時,通過比較植物莖桿水和各水源中穩定同位素組成可實現對植物水分利用來源的判別(Ehleringer et al.,1992;Brunel et al.,1995)。在植物蒸騰過程中,葉片中的水分通過氣孔與外界水汽發生穩定同位素的交換,較輕的水分子會優先逸出葉片,使得葉片中水穩定同位素富集(Dongmann et al.,1974;Farquhar et al.,2003;Butt et al.,2010)。婁源海等(2016)研究發現,相較于濕潤的雨季,高溫低濕的旱季植物葉片水中δ18O更富集。李龍等(2020)研究證明,不同植物由于葉面積指數和葉齡的差異,葉片水中穩定同位素也存在較大的差異。植物葉片水中穩定同位素變化除了與環境水分狀況和植物自身生理特征有關外,還受各氣象因子影響(溫學發等,2008;周盼盼等,2016)。王小婷等(2016)通過比較季節尺度和日尺度上玉米葉片水中穩定同位素變化與各氣象因子的相關性發現,在不同時間尺度上影響葉片水中穩定同位素變化的因子也存在顯著的差異。目前,盡管關于森林系統內部水循環過程中穩定同位素變化的研究較為豐富,但主要集中在干旱半干旱地區(Che et al.,2019;黨宏忠等,2020),對濕潤地區森林系統內部水循環過程中穩定同位素變化還研究較少,且相關研究多基于短時間歷時的觀測數據,無法發現各水體中穩定同位素長時間動態變化特征及其細節。
本研究以亞熱帶濕潤地區長沙為研究區域,以針闊混交林內樟樹(Cinnamomum camphora)和刺杉(Cunninghamia lanceolate)作為研究對象,基于2017年3月—2019年9月期間降水、土壤水、植物莖桿水和葉片水中穩定同位素數據以及兩次典型晴日的植物莖桿水和葉片水中穩定同位素的逐時數據,并結合相關氣象因子,分析了降水、土壤水和植物水中穩定同位素的季節變化特征以及植物水中穩定同位素的日內變化特征。研究目的在于:(1)揭示區域森林系統內不同水體穩定同位素變化的影響機制;(2)結合各氣象因子,明確在不同時間尺度下影響植物葉片水中穩定同位素富集變化的主要因子。
1 材料與方法
1.1 研究區概況
研究場地位于湖南省長沙市望城區八曲河村(28°22′09″N—112°45′43″E),平均海拔在 50 m 左右。受亞熱帶季風的影響,研究區氣候四季分明,雨熱同期。多年平均降水量1415 mm,年內降水存在明顯季節差異,在 3—7月初,受夏季風影響,降水豐富;7月中旬至9月,在副熱帶高壓控制下,伏旱嚴重。多年平均氣溫17.4 ℃,大于0 ℃的多年平均年積溫5457 ℃,全年無霜期約272 d,植物生長期在330 d左右。研究場地的地下水深達18 m,由于人工栽植樹木根系主要分布在1 m以內,因此地下水難以被利用。土壤以紅色的粉砂質黏壤土為主。樣地林分主要是以 15年生常綠闊葉喬木樟樹(Cinnamomum camphora)和常綠針葉喬木刺杉(Cunninghamia lanceolate)、馬尾松(Pinus massoniana)組成的人工混交林,郁閉度為 0.65—0.80,林分密度為 1000 plant·hm?2。雖為常綠混交林,但生長季初期伴隨新葉生長出現大量枯葉掉落,地表常年枯落物質堆積厚度可達5 cm。此外,混交林外圍分布有稀疏低矮灌木梔子花(Gardenia jasminoides)和野茶花(Camellia japonica),覆蓋度在40%左右。
1.2 樣品采集
于2017年3月—2019年9月采集植物莖桿和葉片樣品。為避免降水干擾,樣品采集一般在降水后3—10 d進行,采樣頻率為每月1—3次,共69次。采集植物樣品時,分別選擇3棵長勢良好的樟樹和刺杉作為固定樣樹,用于長期連續取樣,其中,刺杉取樣于2018年9月開始。采集枝條樣品時,剪取冠層下部已木質化的小枝(直徑0.3—0.5 cm,長4—6 cm),并將剪下枝條迅速去皮后裝入 10 mL玻璃瓶內,并用Parafilm封口膜密封、編號、冷凍保存,待測定其水穩定同位素比率。采集葉片樣時,為避免新生葉和衰老葉造成的同位素差異,一般剪取不同方位的成熟葉片,并將其混合后迅速裝入10 mL玻璃瓶,并用Parafilm封口膜密封、編號、冷凍保存,待測定其水穩定同位素比率。每次植物樣品采集至少取3個重復,并記錄完整的采樣時間,以對應相應時刻環境因子的變化。此外,兩種植物日內動態變化過程的取樣(枝條、葉片)于典型無風晴日的2019年5月23—24日和2019年8月28—29日進行,采樣間隔為2 h,采集方式與上面相同。
土壤樣品采集與植物樣品采集同步進行,在樣樹附近利用手持式土壤取樣鉆機(科力SD-1,北京吉奧科技有限公司)鉆取土壤樣,每次鉆取2個平行土樣。鉆孔深度100 cm,0—20 cm取樣間隔為5 cm,20 cm以下取樣間隔為10 cm。采集的土壤樣品密封于10 mL的玻璃瓶中用于測定土壤水中穩定同位素比率。
降水取樣分別在降水日(降水量≥0.1 mm)的北京時間08:00和20:00進行。將采集的降水樣直接裝入30 mL的聚乙烯樣品瓶內,密封、編號且低溫保存,待測定其穩定同位素比率。參照氣象部門的規范,將當日20:00(代表當日08:00—20:00的累積降水量)和次日08:00(代表當日20:00至次日08:00的累積降水量)收集的降水量之和記為當日的降水量,相應時段的降水中穩定同位素值也記為當日。
1.3 環境因子的觀測
土壤體積含水量采用布設在樣地中心位置的云智能管式土壤水分溫度監測儀(RWET-100,智墑,中國)測定。儀器探頭位于0—100 cm每隔10 cm處,監測精度為±2.5%,監測時間自 2017年 2月起,采集頻率為每60 min一次。土壤含水量監測數據利用烘干稱質量法進行水分校正(王銳等,2020b)。
氣溫 T/℃、相對濕度 RH/%、太陽輻射RS/(W·m?2)、風速 U/(m·s?1)等氣象數據均由設置在實驗場地外 50 m處的微型自動氣象站(232,WeatherHawk,美國)獲取。監測時間自 2016年12月起,監測頻率為每30分鐘一次。此外,根據監測的T和RH可計算出林地附近的飽和水汽壓差VPD/kPa,其計算公式為:

1.4 樣品處理與測試
利用全自動真空冷凝抽提系統(LI-2100,北京理加聯合有限公司)從土壤、植物莖桿和葉片中抽提出土壤水、植物莖桿水和葉片水。所有水樣的氫、氧穩定同位素比率均利用氣-液兩用型穩定同位素分析儀(DLT-IWA-35EP,美國Los Gatos Research公司)測定。被測試水樣中的穩定同位素豐度用相對于維也納標準平均海洋水(V-SMOW)穩定同位素比率的千分差值(‰)表示:
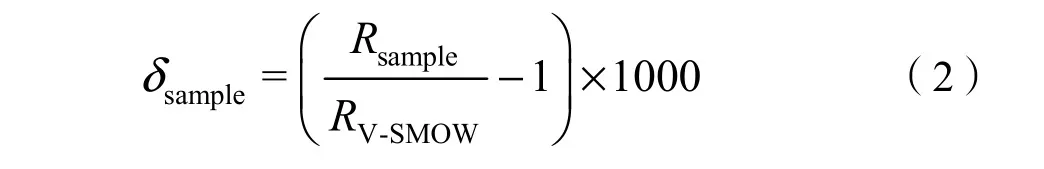
式中:
Rsample和 RV-SMOW分別代表水樣和標準平均海洋水中穩定同位素比率(D/1H或18O/16O)。
在測試植物葉片水中穩定同位素組成時選擇正標樣以提高測試結果準確度。δD和δ18O的測試精度分別為±0.6‰和±0.2‰。本文中,除降水和土壤水穩定同位素的平均值分別為相應時段內降水量和土壤含水量的加權平均值外,其他所有要素的平均值均指時段內的算術平均值。
由于植物樣品在抽提過程中存留有機物,而有機物中的甲醇和乙醇會引起光譜干擾并造成被測定的植物莖桿水和葉片水中穩定同位素值出現偏差,因此需對水樣中的醇類污染進行穩定同位素比率的校正(劉文茹等,2013)。由于所選植物水樣中僅發現甲醇類污染,所以通過實驗建立的甲醇類污染誤差校正公式為:

式中:
ΔδD和Δδ18O分別代表由甲醇類污染導致的植物水樣中穩定同位素值誤差;
NB為窄帶系數且隨植物類型而變化,該參數主要反映水樣中甲醇的污染程度,由 LGR公司的穩定同位素分析處理軟件提供。
校正后的穩定同位素值為實測值減去誤差值。
1.5 植物葉片水中穩定同位素富集程度
盡管植物根系吸收的水分在植物體內運輸的過程中不發生穩定同位素分餾,但是植物蒸騰作用卻會使得葉片水中穩定同位素富集(Farquhar et al.,2003)。葉片水中穩定同位素相較于源水(植物莖桿水)中穩定同位素的富集程度用ΔδL來表示,其計算公式為:

式中:
ΔδL表示葉片水中穩定同位素相較于莖桿水中穩定同位素的富集值;
δA代表植物葉片水中δD(或δ18O);
δB代表植物莖桿水中δD(或δ18O)。
1.6 時期的劃分
通過查閱相關文獻資料,長沙地區雨季主要集中在(3—6月),在7—9月受伏旱影響,降水少,干旱嚴重;而在10月至次年2月,雖然該時期降水少,但溫度低,相對濕度較大,干旱發生頻率小(王文等,2016;劉仲藜等,2020)。基于此,將觀測期分為濕潤期(10月至次年6月)和干旱期(7—9月)兩個時段,深入探討不同水分條件下研究區森林系統內部各水體中水穩定同位素的變化特征。
2 結果與分析
2.1 降水量與土壤含水量變化
圖1給出了2017年3月—2019年9月期間日降水量和土壤含水量隨時間的變化。根據統計,2017、2018和2019年的雨季(3—6月)降水日數分別為64、38和56 d,累積降水量分別為1095.0、366.0和758.9 mm。各年旱季(7—9月)降水日數分別為21、27和10 d,累積降水量分別為287.0、515.0和157.8 mm。此外,2017年10月至次年2月的總降水日數為52 d,累積降水量為290.9 mm;2018年10月至次年2月期間的總降水日數達83 d,累積降水量為 473.2 mm,較上年同期降水量增加62.7%。
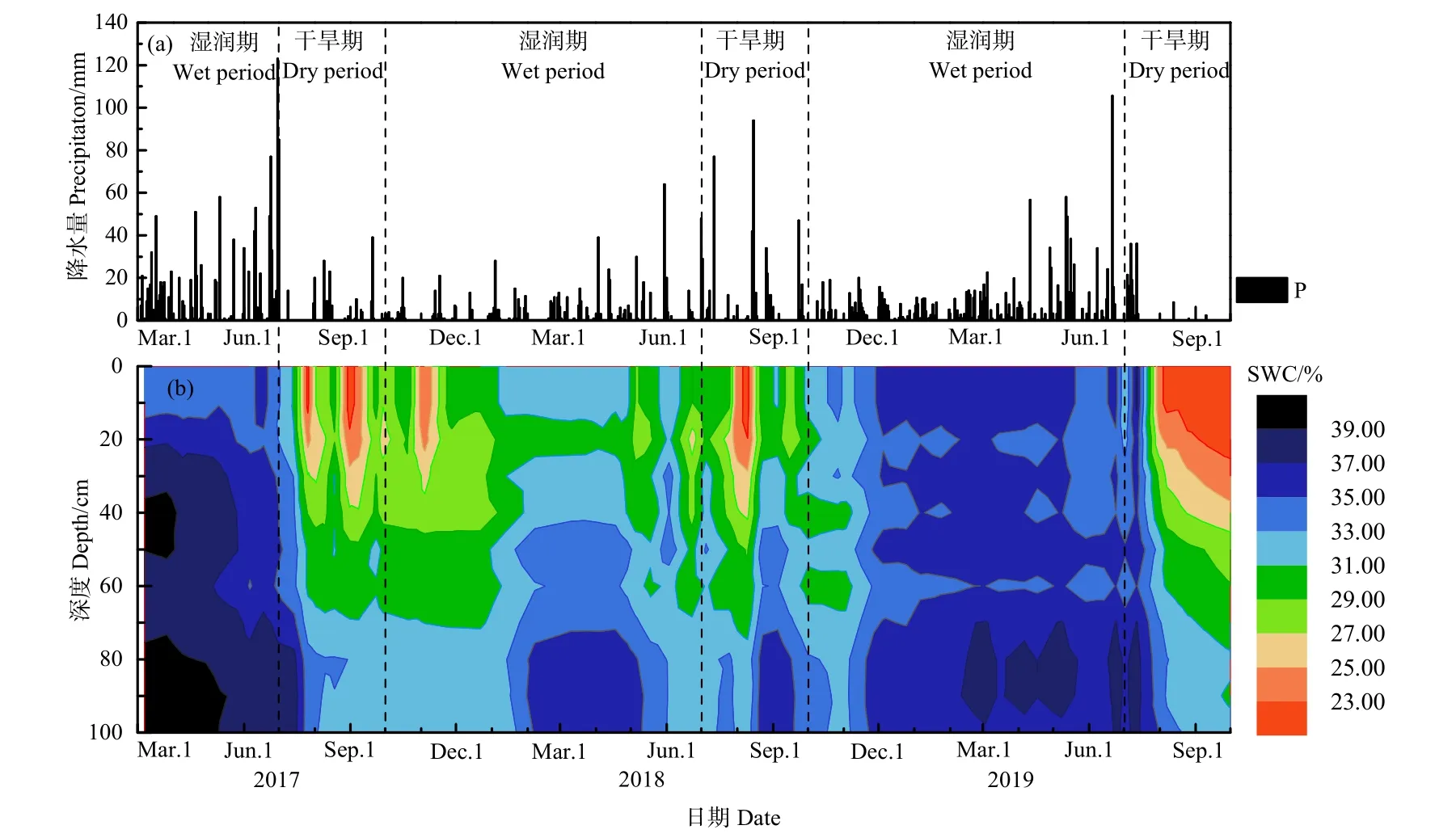
圖1 研究區日降水量(P)和土壤含水量(SWC)隨時間的變化Fig. 1 Temporal variations of precipitation and soil water content in study area
觀測期間土壤含水量在 20.9%—39.9%范圍內變化,并隨深度的增加逐漸遞增。在時間變化上,表層0—10 cm土壤含水量的高值出現在2017年3—6月和2018年10月至次年6月,平均值分別為34.4%和 35.3%,這與兩時期偏多的降水有關。在2017年10月至次年6月,由于降水較2017年3—6月和2018年10月至次年6月偏少,表層土壤含水量明顯低于兩時期,平均值為30.0%。比較發現,在2017、2018和2019年的7—9月,受較少降水補給和較強蒸散發的影響,表層土壤含水量較年內其它時段明顯偏低,分別只有 26.7%、27.9%和21.9%,表明該時期研究區受干旱脅迫程度偏重。
2.2 降水和土壤水中δ18O的變化
降水作為重要的水源輸入項,其穩定同位素組成直接或間接影響著土壤水和植物水中穩定同位素變化。由圖2可知,觀測期間降水中δ18O的變化范圍為?13.32‰—2.85‰,最大值和最小值的出現的時間分別為4—5月和7—9月。根據表1的統計,降水中穩定同位素組成存在明顯的季節差異。在2017年3—6月、2017年10月至次年6月和在2018年10月至次年6月的濕潤期,降水中δ18O的平均值±標準差分別為 (?6.69‰±3.45‰)、(?5.29‰±2.74‰)和 (?4.81‰±2.46‰)。相較于濕潤期,2017、2018和2019年的干旱期內降水中δ18O明顯偏負,平均值±標準差分別為 (?10.38‰± 2.35‰)、(?8.93‰±2.06‰) 和 (?8.64‰±2.19‰)。降水中穩定同位素的季節差異主要與該地區水汽來源的季節變化有關。在濕潤期,長沙地區水汽主要來自于西風環流所攜帶的大陸性氣團,該氣團具有濕度低,蒸發性強的特點,且冬季降水較少,一般以小雨為主,氣團中穩定同位素在運輸過程中被貧化程度低,因此長沙地區降水中穩定同位素更富集。在干旱期,來自熱帶海洋上的氣團,在輸送過程中受強烈對流作用,降水頻繁,且降水量較大,剩余氣團中同位素組成逐漸被貧化,因此長沙地區降水中穩定同位素較為貧化(吳華武等,2012)。
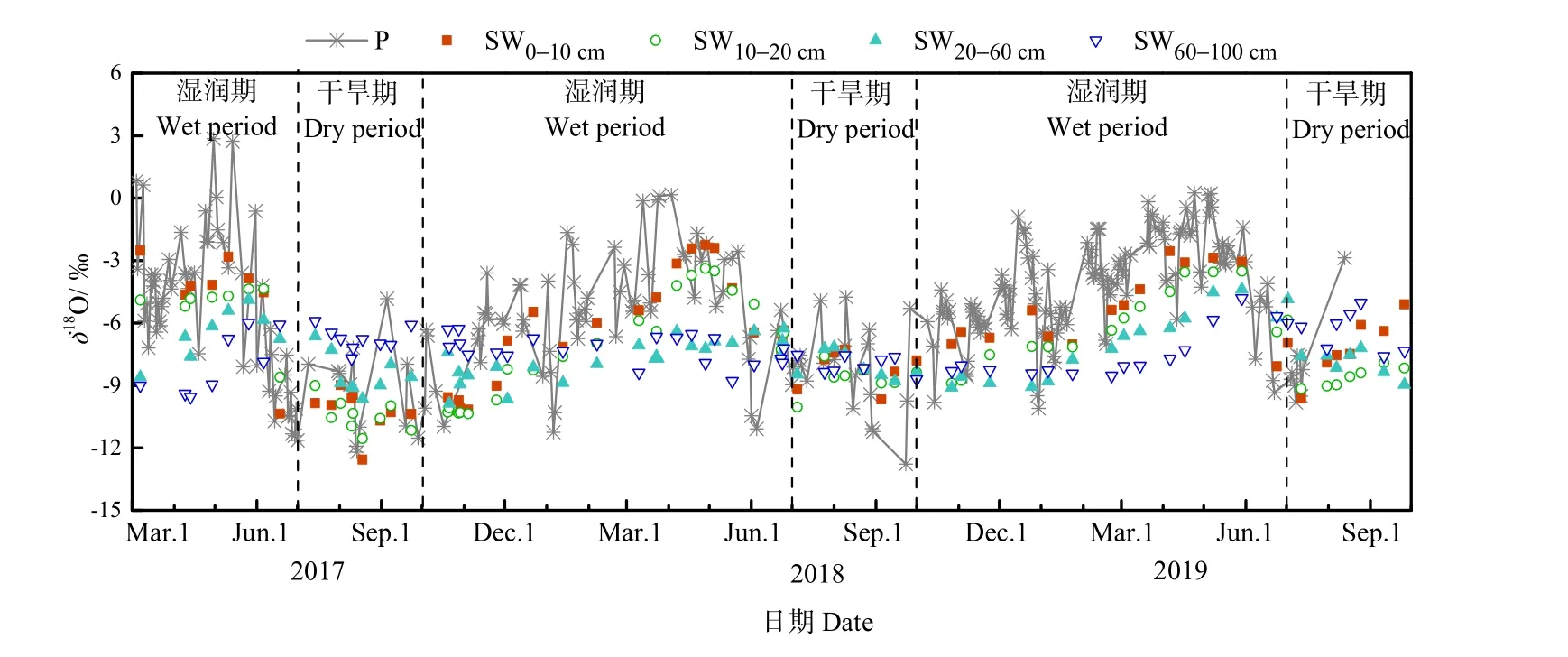
圖2 觀測期內降水(P)、土壤水(SW)中δ18O隨時間的變化Fig. 2 Temporal variations of δ18O of precipitation (P) and soil water (SW) during the observation period
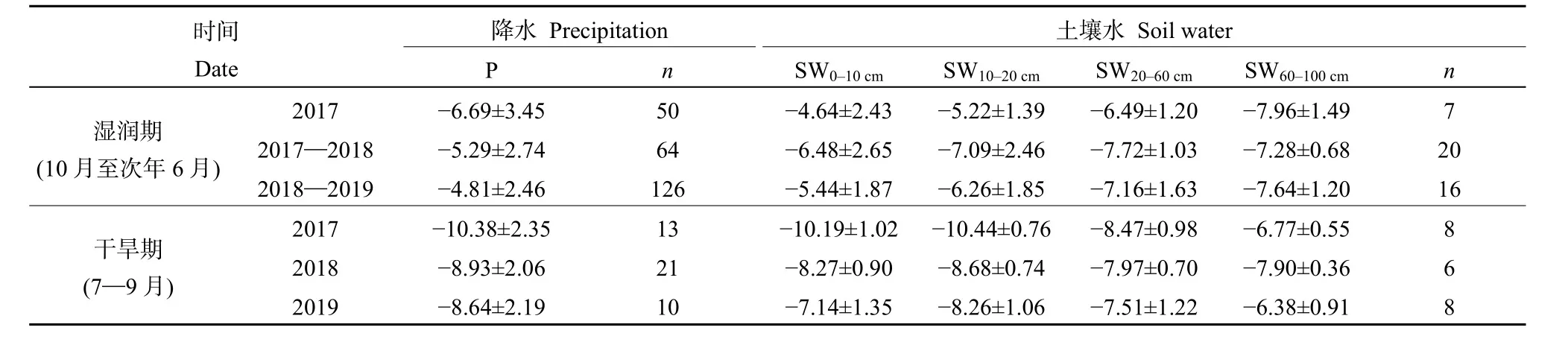
表1 干旱期、濕潤期內降水(P)和土壤水(SW)中δ18OTable 1 δ18O in precipitation (P) and soil water (SW) in wet and dry periods ‰
為進一步分析不同深度土壤水變化特征和確定植物水分利用特征,根據不同深度土壤含水量和土壤水中穩定同位素組成的特點,將土壤含水量和穩定同位素組成相近的相鄰土層合并為同一層次,因此將0—100 cm土壤剖面分為0—10、10—20、20—60和60—100 cm 4個層次。土壤水中δ18O的變化范圍在表層 0—10 cm 最大,為?12.56‰—?2.25‰,并隨深度的增加逐漸減小。可以看到,0—10、10—20、20—60 cm土壤水中δ18O達到最大值和最小值的時間與降水一致,分別為4—5月和7—9月。而60—100 cm土壤水中δ18O達到最大值和最小值的時間較降水存在滯后,分別為 6—7月和9—10月。這表明0—60 cm土壤水受降水直接補給影響較大,而60—100 cm深層土壤水對降水響應存在1—2個月的滯后。
與降水中穩定同位素變化相似,土壤水中δ18O存在明顯季節變化,且隨深度的增加季節變化逐漸減小(表1)。在2017年3—6月、2017年10月至次年6月和2018年10月至次年6月的濕潤期,表層0—10 cm土壤水中δ18O明顯偏正,平均值±標準差分別為 (?4.64‰±2.43‰)、(?6.48‰±2.65‰)和 (?5.44‰±1.87‰),且隨深度的增加土壤水中δ18O逐漸偏負。在2017、2018和2019年的干旱期,表層土壤水中δ18O較各濕潤期偏負,平均值±標準差分別為 (?10.19‰±1.02‰)、(?8.28‰±0.90‰)和(?7.14‰±1.35‰),隨深度的增加土壤水中δ18O 整體呈偏正的趨勢。不同時期土壤水中δ18O隨深度的變化主要與降水在土壤剖面中的補給過程有關,降水優先補給淺層土壤水,在淺層土壤水達到飽和后推動前期土壤水向下逐層補給深層土壤水。
2.3 植物水中δ18O變化
2.3.1 植物水中δ18O的季節變化
圖3給出了取樣期間樟樹和刺杉莖桿水和葉片水中δ18O隨時間的變化,表2則統計了各濕潤期和各干旱期內兩種植物莖桿水、葉片水中δ18O和葉片水穩定同位素富集程度Δ18OL的平均值。
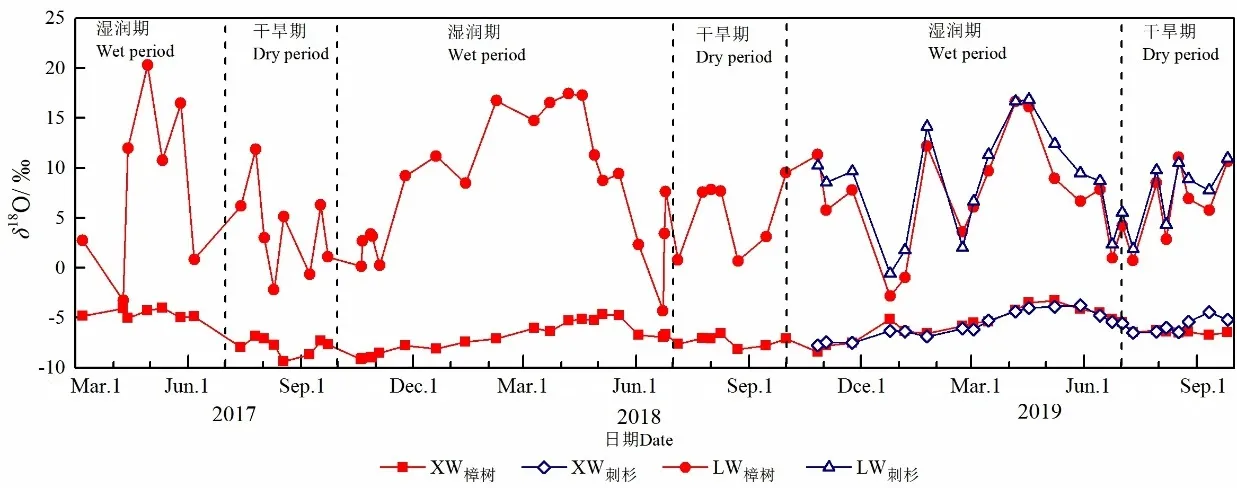
圖3 樟樹和刺杉莖桿水(XW)、葉片水(LW)中δ18O隨時間的變化Fig. 3 Temporal variations of δ18O of xylem water (XW) and leaf water (LW) of C. camphora and C. lanceolate
取樣期間樟樹和刺杉莖桿水中δ18O變化范圍分別為?9.40‰— ?3.30‰和?8.35‰— ?3.73‰。與降水相似,兩種植物莖桿水中δ18O的最大值和最小值分別出現在4月和8月。時間變化上,在2017年3—6月、2017年10月至次年6月和2018年10月至次年6月的濕潤期,樟樹莖桿水中δ18O明顯偏正,平均值±標準差分別為 (?4.59‰±0.43‰)、(?6.99‰±1.49‰)和 (?5.66‰±1.54‰);而在 2017、2018和2019的干旱期,樟樹莖桿水中δ18O較各濕潤期偏負,平均值±標準差分別為 (?7.84‰±0.85‰)、(?7.39‰±0.58‰)和 (?6.34‰±0.37‰)。比較發現,刺杉莖桿水中δ18O在2018年10月至次年6月的濕潤期和在2019年7—9月的干旱期均與樟樹莖桿水中δ18O接近,平均值±標準差分別為(?5.72‰±1.36‰)和 (?5.73‰±0.73‰),這可能與兩時期內樟樹與刺杉水分利用來源相似有關。結合表1中不同時期4層土壤水中δ18O比較發現,在各濕潤期,兩種植物莖桿水中δ18O與0—20 cm土壤水中δ18O接近;而在各干旱期,兩種植物莖桿水中δ18O則主要與20—60 cm和60—100 cm的土壤水中δ18O相似。這表明兩種植物在各濕潤期主要利用0—20 cm的淺層土壤水,而在各干旱期則主要利用20—60 cm和60—100 cm的土壤水。
相較于莖桿水,葉片水中穩定同位素明顯富集,且變幅更大。樟樹和刺杉葉片水中δ18O的變化范圍分別為?4.32‰—20.29‰和?1.60‰—15.84‰。由表2可知,各濕潤期樟樹和刺杉葉片水中δ18O均較各干旱期偏正,兩種植物葉片水Δ18OL也在濕潤期大于干旱期。根據Craig et al.(1961)對大氣水線的定義,將植物葉片水中δD和δ18O之間的線性關系定義為葉片水線(LWL)。研究表明,LWL斜率的大小可指示植物葉片水中同位素動力分餾程度的強弱,斜率越小表示葉片水中穩定同位素組成受動力分餾作用越強(Yakir et al.,1990;Yepez et al.,2003)。結合表3,濕潤期內樟樹和刺杉 LWL 分別為δD=3.06δ18O?21.74(R2=0.68)和δD=2.44δ18O?16.84(R2=0.63)。干旱期內兩種植物LWL 分別為δD=3.26δ18O?39.70(R2=0.67)和δD=2.61δ18O?27.72(R2=0.71),其斜率均大于濕潤期內兩種植物 LWL斜率。從時間變化上看,無論是樟樹還是刺杉,葉片水中穩定同位素的富集程度和動力分餾程度均在濕潤期大于干旱期。在濕潤期,土壤水分充足,兩種植物采取揮霍型水分利用策略,植物葉片水與大氣水汽交換頻繁。在干旱期,由于土壤水分虧缺嚴重,兩種植物用水趨于保守,植物通常選擇關閉部分氣孔減少蒸騰耗水,植物葉片水與大氣水汽交換受阻(羅紫東等,2016)。因此,在濕潤期,樟樹和刺杉葉片水中δ18O、Δ18OL較干旱期大,而LWL斜率則明顯小于干旱期。
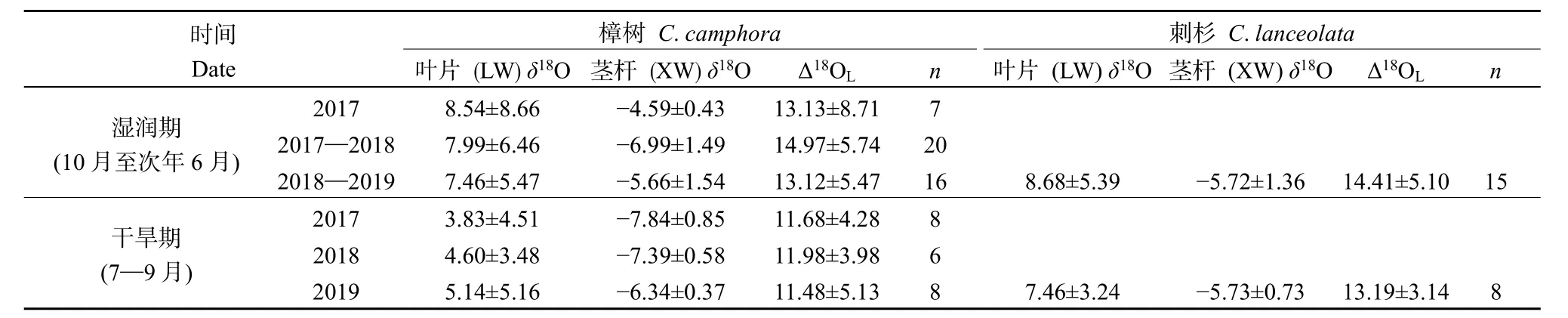
表2 干旱期、濕潤期內樟樹與刺杉莖桿水中δ18O、葉片水中δ18O和Δ18OLTable 2 δ18O in xylem water, δ18O and Δ18OL in leaf water of C. camphora and C. lanceolata in dry and wet periods ‰

表3 不同時間尺度上樟樹和刺杉葉片水線方程(LWL)Table 3 Leaf water line (LWL) of C. camphora and C. lanceolata on different time scales
2.3.2 植物水中δ18O的日內變化
圖4給出了濕潤期和干旱期典型晴日內兩種植物莖桿水、葉片水中δ18O的逐時變化。在所選濕潤期的典型晴日,樟樹和刺杉莖桿水中δ18O的平均值±標準差分別為 (?3.58‰±0.31‰)和 (?3.80‰±0.32‰);在所選干旱期的典型晴日,兩植物莖桿水中 δ18O 平均值±標準差分別為 (?6.09‰±0.39‰)和(?6.63‰±0.34‰)。可以看出,樟樹和刺杉莖桿水中δ18O均無明顯的日內變化,這說明兩種植物水分利用來源相對穩定。
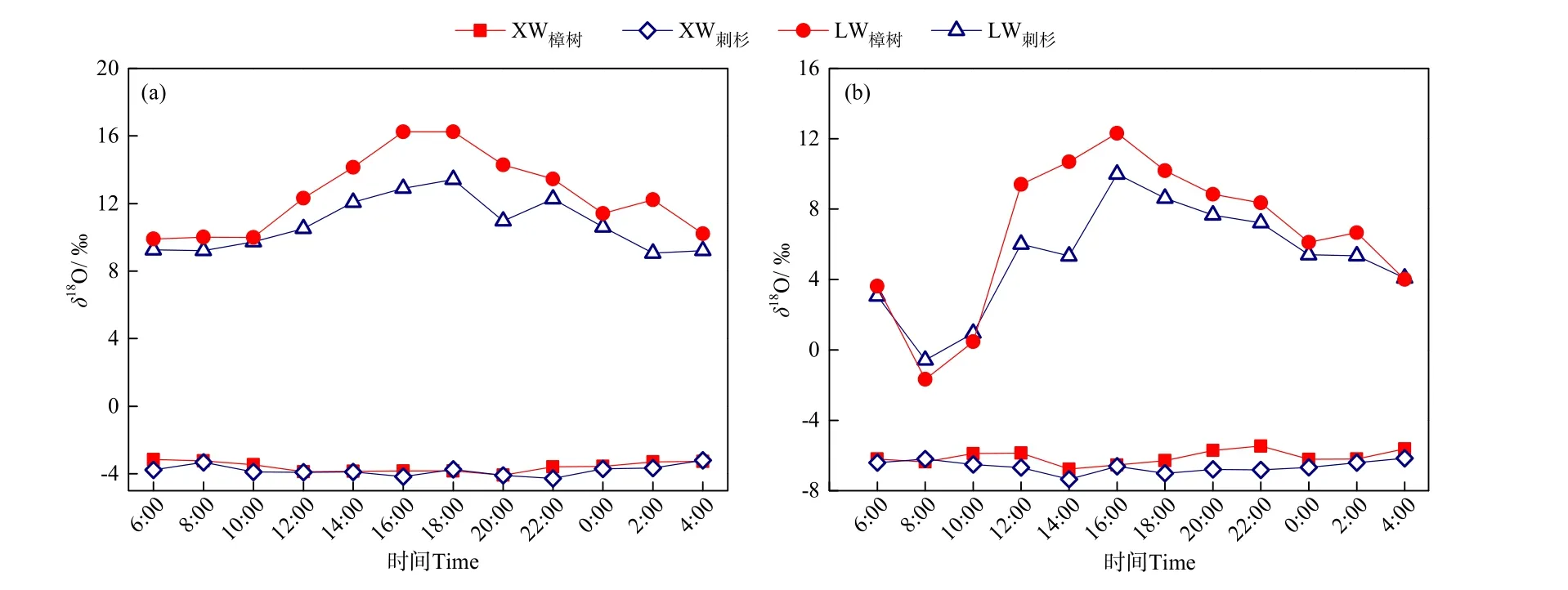
圖4 濕潤期和干旱期典型晴日內樟樹和刺杉莖桿水(XW)、葉片水(LW)中δ18O的逐時變化Fig. 4 Hourly variation of δ18O in xylem water (XW) and leaf water (LW) of C. camphora and C. lanceolata on typical sunny days during wet and dry periods
相較于莖桿水,樟樹和刺杉葉片水中δ18O存在明顯的日內變化。在濕潤期和干旱期的典型晴日,樟樹葉片水中 δ18O 分別在 9.90‰—16.25‰和?1.68‰—12.30‰內變化,刺杉葉片水中δ18O的變化范圍則分別為 9.07‰—13.42‰和?0.58‰—10.01‰。在時間變化上,兩種植物葉片水中 δ18O最大值均出現在16:00—18:00,而最小值則出現在06:00—08:00。與葉片水中δ18O對應,兩種植物葉片水Δ18OL均在16:00—18:00最大,在06:00—08:00最小。
兩時期典型晴日,樟樹和刺杉葉片水中穩定同位素組成存在明顯差異。在濕潤期典型晴日樟樹和刺杉葉片水中 δ18O偏正,平均值±標準差分別為(12.54‰±2.35‰)和 (10.77‰±1.56‰),對應的兩種植物葉片水 Δ18OL分別為 (16.12‰±2.60‰)和(14.57‰±1.77‰)。在干旱期典型晴日,樟樹和刺杉葉片水中 δ18O偏負,平均值±標準差分別為(6.58‰±4.26‰)和 (5.26‰±3.06‰),對應的葉片水Δ18OL也小于濕潤期典型晴日,平均值±標準差分別為 (12.68‰±4.35‰)和 (12.20‰±3.25‰)。結合表3,樟樹和刺杉 LWL在濕潤期典型晴日分別為δD=1.05δ18O+17.24(R2=0.66)和 δD=2.17δ18O+1.03(R2=0.79)。在干旱期典型晴日,兩種植物LWL分別為 δD=1.94δ18O?29.48(R2=0.94)和 δD=2.44δ18O?27.71(R2=0.92),其斜率大于濕潤期典型晴日兩種植物 LWL的斜率。上述結果表明,兩種植物葉片水中穩定同位素富集程度和動力分餾程度在濕潤期典型晴日更大,而在干旱期典型晴日更小,這可能與兩晴日內植物莖桿水中 δ18O大小及其微氣象環境有關。在濕潤期典型晴日,由于兩種植物葉片水水源(莖桿水)中δ18O較干旱期典型晴日明顯偏正,在相同的蒸騰作用下,兩種植物葉片水中δ18O 更富集。Butt et al.(2010)和 Lai et al.(2008)研究發現,在暖濕的環境中,植物葉片水內外水汽壓差較小,不利于葉片水與大氣進行水汽交換,植物葉片水中穩定同位素的分餾程度較弱。濕潤期典型晴日T、RH和VPD平均值分別為23.8 ℃、49.5%和1.5 kPa;而干旱期典型晴日,T、RH和VPD均高于濕潤期,平均值分別為 31.9 ℃、61.5%和 1.8 kPa,因此兩種植物葉片水中穩定同位素的富集程度和動力分餾程度在濕潤期典型晴日更大。
2.4 影響植物Δ18OL變化的氣象因子
植物葉片水中穩定同位素富集變化過程主要受區域微生境的氣象條件的影響(羅倫等,2013)。表 4給出了不同時間尺度上樟樹和刺杉葉片水Δ18OL與各氣象因子(溫度T、相對濕度RH、太陽輻射RS、飽水汽壓差VPD)相關系數。
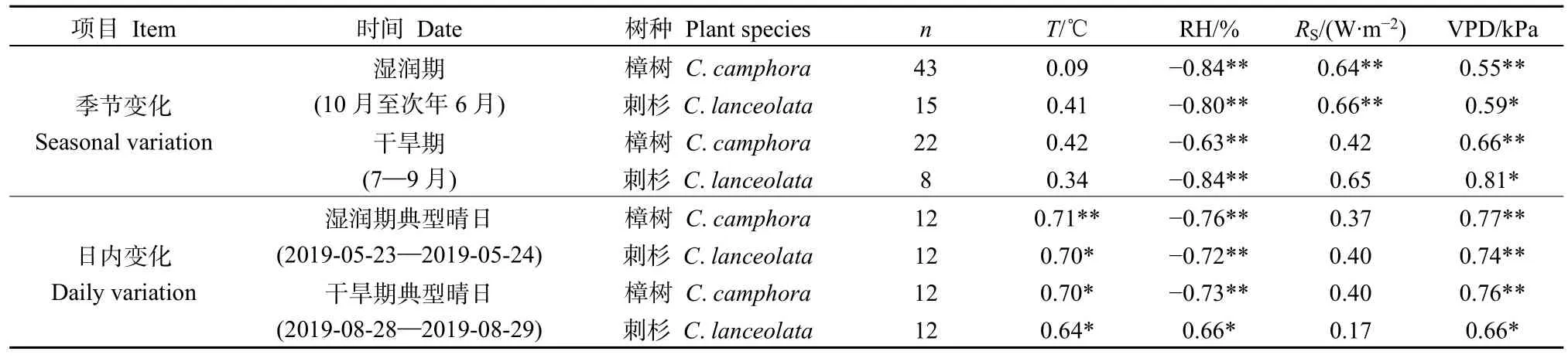
表4 不同時間尺度下樟樹和刺杉Δ18OL與各氣象因子的相關關系Table 4 Correlation between leaf water Δ18OL and meteorological factors of C. camphora and C. lanceolata in different time scales
在季節變化上,無論是濕潤期還是干旱期,樟樹和刺杉葉片水 Δ18OL與 T均無顯著相關關系(P>0.05),與RH和VPD分別呈極顯著的負相關和正相關關系(P<0.01)。此外,在濕潤期,兩植物葉片水Δ18OL與RS表現出極顯著正相關;在干旱期,樟樹葉片水Δ18OL與RS呈顯著的正相關,刺杉由于觀測樣本量較少,葉片水中Δ18OL與RS表現出不顯著的正相關。這在一定程度上說明,在年內兩種植物葉片水中穩定同位素富集程度主要受 RH、RS和VPD影響,而受T的影響較小。在濕潤期和干旱期的典型晴日內,樟樹和刺杉葉片水Δ18OL與T均表現出顯著的正相關(P<0.05),與RH和VPD分別表現出呈極顯著負相關和正相關。此外,在兩時期典型晴日內,兩種植物葉片水Δ18OL與RS均無顯著相關關系。這表明在日內逐時變化中,T、RH和 VPD是影響兩種植物葉片水中穩定同位素富集程度的主要因子,而RS對兩種植物葉片水中穩定同位素富集程度的影響較小。
RH和VPD控制著植物蒸騰速率和氣孔導度的變化,進而影響著植物葉片水Δ18OL(Farquhar et al.,2003;Butt et al.,2010;Barbour et al.,2000)。即當RH較大時,VPD則越小,葉片內外水汽壓接近,氣孔導度減小,植物蒸騰作用減弱,葉片水與冠層水汽的交換減弱,葉片水中穩定同位素富集程度低;反之,葉片水中穩定同位素富集程度低較大。因此,無論是季節變化還是日內變化,兩種植物葉片水Δ18OL與RH和VPD相關性極為顯著。
RS通過影響植物光合作用,進而影響葉片水中穩定同位素分餾。植物進行光合作用時,通過氣孔進入葉片的一部分 CO2在與水合成葡萄糖的過程中,羰基氧與H2O分子中氧原子發生交換,使得葉片水氧同位素發生分餾(Guy et al.,1993);而另一部分CO2與葉片水在碳酸酐酶催化下,經過穩定同位素平衡反應后返回大氣,葉片水中氧同位素同樣發生分餾(羅倫等,2013)。因此在季節變化上,樟樹和刺杉葉片水Δ18OL與RS的相關性顯著;而在日內變化中,由于夜間RS幾乎接近0,但兩種植物葉片水 Δ18OL在夜間仍較大,故兩種植物葉片水Δ18OL與RS相關性不顯著。
除RH、VPD和RS影響外,樟樹和刺杉葉片水中Δ18OL變化還與T有關。T通過調節冠層溫度改變葉片水中穩定同位素的分餾過程,一般情況下,溫度越高時,蒸發越強,剩余水體中穩定同位素越富集(Yang et al.,2015;Hua et al.,2019;Dai et al.,2020)。基于該原理,在日內變化中,兩植物葉片水中Δ18OL隨T的升高而逐漸增大。在季節變化中,各次取樣主要在天氣晴朗的早上 08:00—09:00,濕潤期(或干旱期)內取樣時刻的T值較為接近,因此可能導致Δ18OL與T的相關性不顯著。
3 結論
受水汽來源的季節變化影響,研究區降水中δ18O表現出在濕潤期(10月至次年6月)偏正,在干旱期(7—9月)偏負的季節變化。表層土壤受降水直接補給,土壤水中δ18O與降水中δ18O的季節變化趨勢一致,且隨深度的增加土壤水中δ18O季節變化逐漸減小。在濕潤期,由于樟樹和刺杉主要利用的0—20 cm土壤水,莖桿水中δ18O偏正;在干旱期,兩種植物主要利用的20 cm以下土壤水且該層土壤水中δ18O偏負,因此莖桿水中δ18O較濕潤期偏負。與莖桿水中δ18O較大的季節變化不同,在日內兩種植物莖桿水中δ18O均不存在明顯變化。
季節變化上,在環境水分充足時期,植物揮霍型水分利用策略加強了植物葉片水與大氣的水汽交換,使得葉片水中穩定同位素富集程度和強動力分餾程度加大,即葉片水中δ18O和Δ18OL值更高。而在水分虧缺嚴重時期,植物通過關閉部分氣孔減弱蒸騰耗水,使得葉片水中穩定同位素富集程度和動力分餾程度變小,即葉片水中δ18O和Δ18OL值明顯更小。日內變化上,樟樹和刺杉葉片水中δ18O和Δ18OL均在16:00—18:00達到最大,在06:00—08:00最小。相較于暖濕的天氣(干旱期典型晴日),在干燥的天氣(濕潤期典型晴日)植物葉片水中穩定同位素富集程度和動力分餾程度更大,即植物葉片水中δ18O和Δ18OL更大。
T、RH、VPD和 RS分別通過調節植物冠層溫度、葉片氣孔導度、蒸騰速率以及光合速率等方式影響植物蒸騰過程,使得植物葉片水中穩定同位素發生明顯分餾。不同時間尺度上影響植物葉片水中穩定同位素富集變化的氣象條件存在差異。在季節變化上,兩種植物葉片水中δ18O的富集程度主要受RH、RS和VPD的影響;在日內變化上,影響兩種植物葉片水 δ18O富集程度的主要氣象因子分別為T、RH和VPD。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
