6株芽孢桿菌的分離鑒定和生物學特性
2021-09-12 13:20:48田照輝徐紹剛董穎胡紅霞王巍東天孫愛
江蘇農業(yè)科學 2021年13期
田照輝 徐紹剛 董穎 胡紅霞 王巍 東天 孫愛
摘要:為了篩選適合漁用微生態(tài)制劑的益生菌,從池塘邊發(fā)酵魚肉湯中分離芽孢桿菌,結合形態(tài)學、生理生化特征、16S rRNA及gyrA、gyrB、rpoB和purH基因序列比對分析對其進行鑒定,并對其產(chǎn)淀粉酶、產(chǎn)蛋白酶能力及生長情況進行研究。結果表明,分離篩選的6株芽孢桿菌分屬于4個種,1株為蠟樣芽孢桿菌(Bacillus cereus),1株為阿氏芽孢桿菌(B. aryabhattai),2株為貝萊斯芽孢桿菌(B. velezensis),2株為解淀粉芽孢桿菌(B. amyloliquefaciens),6株菌在LB液體培養(yǎng)基中生長良好,均具有產(chǎn)蛋白酶能力,其中4株具有產(chǎn)淀粉酶能力。
關鍵詞:芽孢桿菌;分離鑒定;生物學特性;產(chǎn)淀粉酶能力;產(chǎn)蛋白酶能力
中圖分類號: S917.1 ?文獻標志碼: A ?文章編號:1002-1302(2021)13-0157-05
一直以來,抗生素和藥物殘留是食品安全的重要威脅,2018年農業(yè)農村部組織制定了《獸用抗菌藥使用減量化行動試點工作方案》,推行減抗行動。微生態(tài)制劑能夠提高動物的免疫力和生產(chǎn)性能,為綠色飼料添加劑,在水產(chǎn)養(yǎng)殖中,微生態(tài)制劑還能有效調控水質,是實現(xiàn)高效綠色養(yǎng)殖模式的有效措施之一[1]。
芽孢桿菌是一類重要的微生態(tài)制劑,可以形成芽孢,耐高溫,具有較強的生存能力。在缺乏營養(yǎng)或不良環(huán)境時,芽孢桿菌可以生成芽孢來增強抗逆性進入消化道,調節(jié)腸道菌群,抑制病原菌生長。芽孢桿菌還能促進機體免疫器官及組織的成熟,可以促進淋巴細胞數(shù)量增加,從而提高機體的體液免疫、細胞免疫水平,芽孢桿菌在動物、人體的微生態(tài)調節(jié)中得到了高度重視和廣泛應用,是較好的微生態(tài)制劑菌種[2]。劉文亮等在飼料中添加蠟樣芽孢桿菌投喂凡納濱對蝦(Litopenaeus vannamei),發(fā)現(xiàn)可以改變凡納濱對蝦的腸道微生物組成,提高其生長速度,增強其抗病能力[3],將蠟樣芽胞桿菌添加到凡納濱對蝦養(yǎng)殖水體中,可以提高凡納濱對蝦抗白斑綜合征病毒(white spot syndrome virus,WSSV)和感染能力[4]。孫盛明等在飼料中添加枯草芽孢桿菌,發(fā)現(xiàn)能夠提高團頭魴幼魚的生長性能、抗氧化能力并改變其腸道菌群結構[5]。張歡歡等在生物絮團對蝦養(yǎng)殖系統(tǒng)中添加其分離到的芽孢桿菌,發(fā)現(xiàn)能夠改善菌群結構,抑制弧菌生長[6]。芽孢桿菌、乳酸菌益生菌在卡他扇貝(Argopecten ventricosus)早期發(fā)育過程中能夠促進卡他扇貝生長、抵抗溶藻弧菌病[7]。在飼料中添加枯草芽孢桿菌后,草魚(Ctenopharyngodon idellus)腸道和肝胰臟消化酶的活性顯著提高,腸道芽孢桿菌、假單胞菌數(shù)量極顯著提高,致病性弧菌、大腸稈菌數(shù)量極顯著減少[8]。芽孢桿菌能產(chǎn)生胞外酶,如高蛋白酶、高淀粉酶活性,可以高效降解養(yǎng)殖殘餌中的蛋白和淀粉,改善修復養(yǎng)殖環(huán)境。將芽孢桿菌類應用于水產(chǎn)養(yǎng)殖,進行水質調控,還可作為有害菌的拮抗菌替代或減少抗生素的使用。枯草芽孢桿菌 HAINUP40可顯著降低水體中的亞硝酸鹽、氨氮和化學需氧量,不僅可以作為水質改良劑改善水體環(huán)境,還可作為飼料添加劑提高羅非魚的疾病抵抗力,預防無乳鏈球菌疾病的發(fā)生[9]。短小芽孢桿菌(Bacillus pumilus)能夠抑制多種水產(chǎn)病原菌,對水體環(huán)境的耐受能力強,具有防治水產(chǎn)養(yǎng)殖病原菌的應用潛力[10]。高艷俠等篩選的貝萊斯芽孢桿菌具有較強的廣譜拮抗水產(chǎn)常見病原菌功能,能有效抑制水產(chǎn)養(yǎng)殖魚類常見病原菌如無乳鏈球菌、 海豚鏈球菌等[11]。張皎皎等篩選的甲基營養(yǎng)型芽孢桿菌(B. methylotrophicus)對嗜水氣單胞菌的抑制效果顯著[12]。
有益微生物在水產(chǎn)養(yǎng)殖中能有效改善水質,其應用越來越廣泛,但仍存在一些問題,如有些菌種的效果不高,穩(wěn)定性差,不能快速成為優(yōu)勢菌群等[13],微生態(tài)制劑菌種還具有種屬特異性和地域性,一般從微生物生活的環(huán)境中分離和篩選特定功能益生菌,因此從相應的水產(chǎn)養(yǎng)殖環(huán)境分離菌株,用于篩選水產(chǎn)微生態(tài)制劑菌種更具有合理性[14-16]。
1 材料與方法
1.1 樣品的采集
2018年夏將池塘中零星死亡的魚收集起來,在塘邊煮成肉糜,任其自然發(fā)酵,待發(fā)酵肉湯無色無臭后,取回至北京市水產(chǎn)科學研究所實驗室進行菌種分離。
1.2 培養(yǎng)基
1.2.1 營養(yǎng)瓊脂培養(yǎng)基 10 g/L蛋白胨、3 g/L牛肉膏、5 g/L氯化鈉、2%瓊脂粉,121 ℃高壓滅菌 15 min ,倒平板上備用[17]。
1.2.2 產(chǎn)蛋白酶菌株篩選培養(yǎng)基 12 g/L脫脂奶粉、20 g/L營養(yǎng)瓊脂,113 ℃高壓滅菌15 min,倒平板上備用。
1.2.3 產(chǎn)淀粉酶菌株篩選培養(yǎng)基 可溶性淀粉 5 g/L、氯化鈉5 g/L、pH值7.2,將除瓊脂外的各成分溶解于蒸餾水中,校正pH值,加入2%瓊脂粉,于115 ℃高壓滅菌15 min,倒平板備用。
1.3 菌種分離
在無菌條件下,于滅菌后的三角瓶中加入 90 mL 無菌水和10 mL發(fā)酵肉湯,混勻后取10 mL懸浮液置于無菌試管中,在80 ℃保溫15 min殺死非芽孢菌體,之后取1 mL進行梯度稀釋制成懸液,濃度依次為發(fā)酵肉湯原液的10-2、10-3、10-4、10-5、10-6。分別取50 μL各稀釋液涂布于營養(yǎng)瓊脂培養(yǎng)基,37 ℃恒溫培養(yǎng)24 h后,挑選單菌落,傳代3次以上,待菌落穩(wěn)定后,挑取單菌落用營養(yǎng)瓊脂液體培養(yǎng)基在37 ℃、150 r/min下培養(yǎng)24 h,進行革蘭氏染色、芽孢染色,在顯微鏡下觀察,選取有芽孢的G+菌,滅菌甘油保存于-80 ℃。
1.4 生理生化鑒定
利用HIBacillusTM Identification Kit和參照微生物試驗中的糖酵解試驗進行鑒定。
1.5 分離純化芽孢菌株產(chǎn)蛋白酶活力和產(chǎn)淀粉酶活力測定
挑選單菌落點種于“1.2.2”節(jié)所述培養(yǎng)基,于30 ℃恒溫培養(yǎng)17 h,取出后4 ℃放置6 h,以芽孢桿菌利用底物產(chǎn)生透明圈直徑和菌體直徑的比值表示產(chǎn)酶活性。
挑選單菌落點種于“1.2.3”節(jié)所述培養(yǎng)基,于30 ℃恒溫培養(yǎng)17 h,取出后4 ℃放置6 h,滴加魯哥氏碘液顯色,以芽孢桿菌利用底物產(chǎn)生透明圈直徑和菌體直徑的比值表示產(chǎn)酶活性。
1.6 分子生物學鑒定
挑選純化的菌株參照曹鳳明等的方法[17-18]設計引物,將純化的菌株送北京擎科生物技術有限責任公司進行16S rRNA、gyrA、gyrB、rpoB、purH基因測序,測序引物序列見表1。
1.7 生長曲線的測定
將保存的芽孢桿菌在營養(yǎng)瓊脂培養(yǎng)基平板上復蘇,挑單克隆在LB培養(yǎng)基中于37 ℃、150 r/min過夜培養(yǎng),將過夜培養(yǎng)的芽孢桿菌按1%的量接種于100 mL LB液體培養(yǎng)基中,于37 ℃、150 r/min振蕩培養(yǎng),每隔2 h使用伯樂(Bio-Rad)公司的SmartSpec Plus分光光度計檢測菌液D600 nm,以測定D600 nm為縱坐標、以時間為橫坐標繪制生長曲線。
2 結果與分析
2.1 菌株的分離純化
經(jīng)過初篩、復篩,選取6株分離得到的菌株,將其編號為2019B1~2019B6,各菌株的菌落形態(tài)、革蘭氏染色和芽孢產(chǎn)生情況見表2。
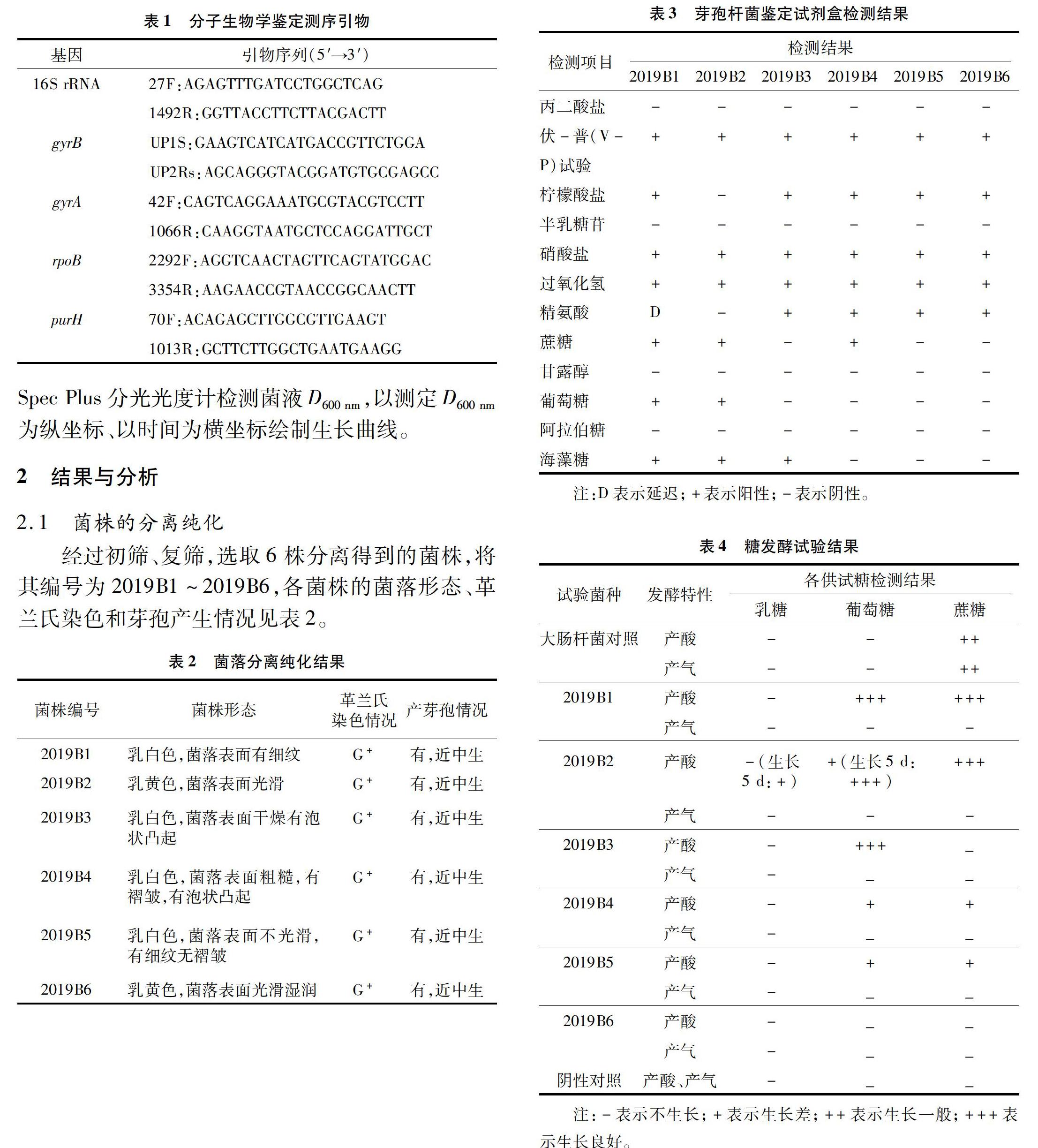
2.2 生理生化反應的鑒定
利用芽孢桿菌鑒定試劑盒HIBacillusTM Identification Kit的檢測結果見表3,糖酵解試驗結果見表4。
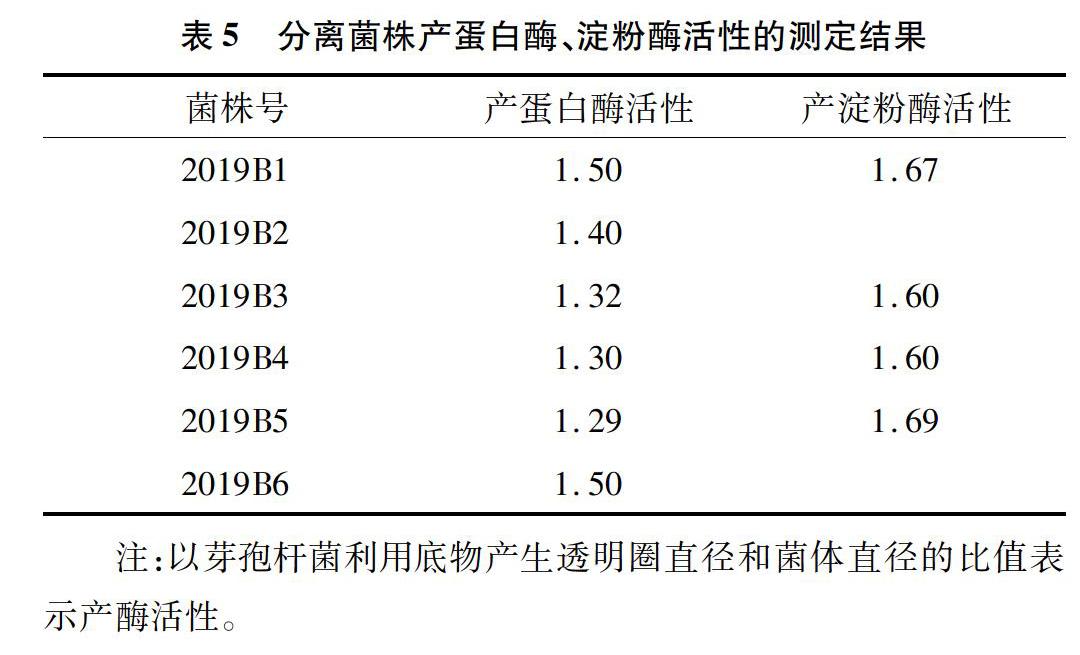
2.3 產(chǎn)淀粉酶、蛋白酶活性
用透明圈直徑/菌落直徑的值表示產(chǎn)酶能力[19]。由表5可以看出,6株菌株均具有產(chǎn)蛋白酶能力,其中2019B1和2019B6產(chǎn)蛋白酶能力最強;2019B2、2019B6無產(chǎn)淀粉酶能力,其他4株具有產(chǎn)淀粉酶能力,其中2019B5產(chǎn)淀粉酶能力最強。
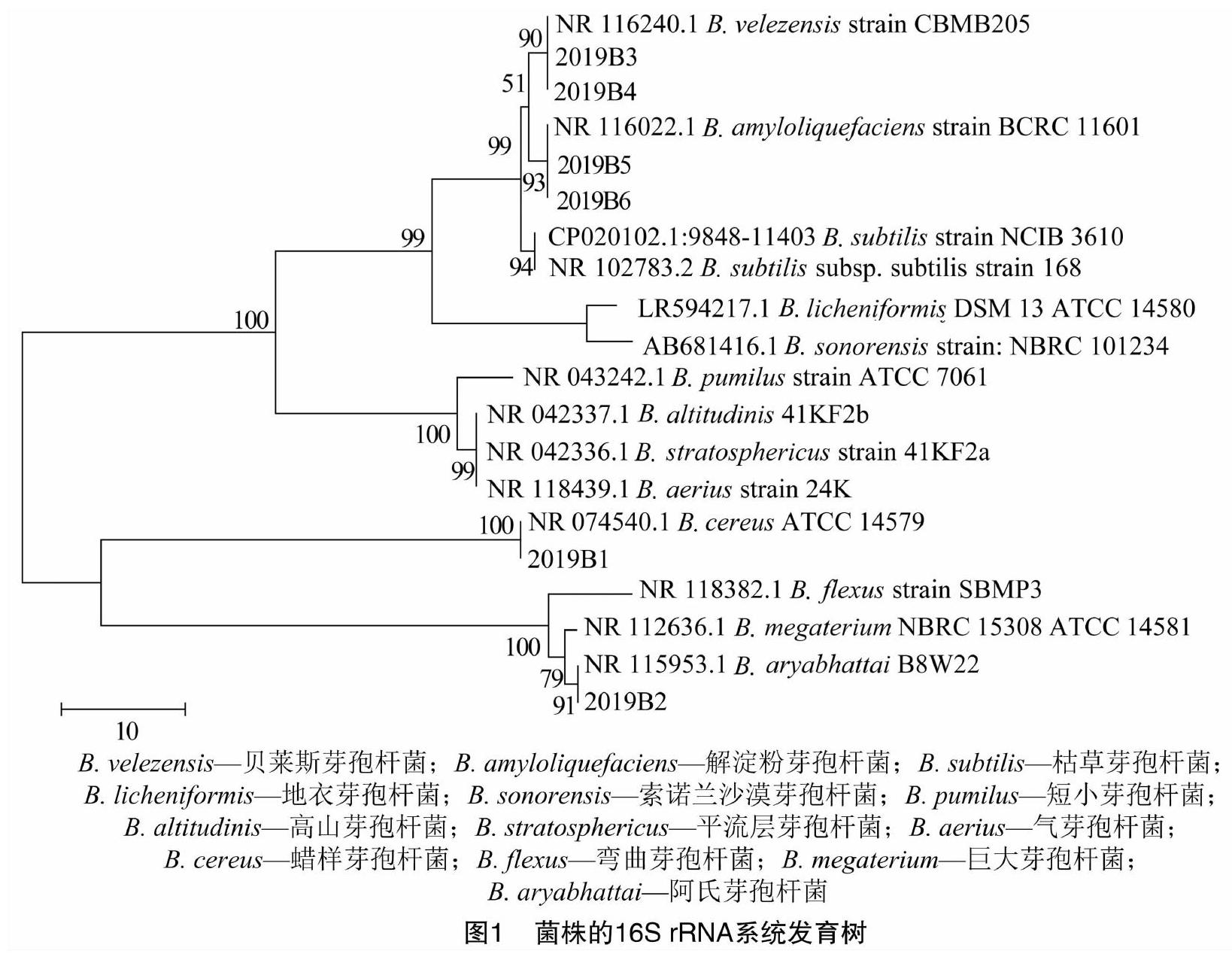
2.4 分子生物學鑒定結果和系統(tǒng)發(fā)育樹
將各菌株的16S rRNA核苷酸序列在GenBank中進行BLAST比對,發(fā)現(xiàn)每株都有多個與其同源性達99%以上的種,選取GenBank中與6株菌株同源性高的菌株的模式菌株的16S rRNA序列,用MEGA 6軟件進行同源性比對并構建鄰接(NJ)樹,自展次數(shù)為1 000次(圖1)。由系統(tǒng)發(fā)育樹可見,2019B1與蠟樣芽孢桿菌(B. cereus)聚為一支,2019B2與阿氏芽孢桿菌(B. aryabhattai)聚為一支,2019B3、2019B4與貝萊斯芽孢桿菌(B. velezensis)聚為一支,2019B5、2019B6與解淀粉芽孢桿菌(B. amyloliquefaciens)聚為一支。
除利用16S rRNA序列進行測序比對外,還對各菌株進行gyrA、gyrB、rpoB和purH基因的測序,并在GenBank中進行BLAST比對,發(fā)現(xiàn)2019B1的purH基因序列比對結果為蠟樣芽孢桿菌,2019B3、2019B4的gyrA、purH基因序列的比對結果分別為貝萊斯芽孢桿菌、解淀粉芽孢桿菌,rpoB基因序列的比對結果均為貝萊斯芽孢桿菌,2019B5的gyrA基因序列比對結果為解淀粉芽孢桿菌,2019B6的rpoB、purH基因序列比對結果為解淀粉芽孢桿菌。
2.5 菌株鑒定結果
結合形態(tài)學、生理生化和分子生物學檢測結果可知,2019B1為蠟樣芽孢桿菌(B. cereus),2019B2為阿氏芽孢桿菌(B. aryabhattai),2019B3、2019B4為貝萊斯芽孢桿菌(B. velezensis),2019B5和2019B6為解淀粉芽孢桿菌(B. amyloliquefaciens)。
2.6 生長曲線
由圖2可以看出,6株芽孢桿菌均能在37 ℃的LB液體培養(yǎng)基中正常生長,達到較高濃度。其中,2019B2長勢最好,培養(yǎng)13 h時已接近穩(wěn)定期;2019B1、2019B2在培養(yǎng)2 h時就開始進入快速增長期, 在培養(yǎng)6.3 h后濃度增長放緩, 2019B1尤其明顯;2019B4在培養(yǎng)4 h才開始生長,并逐漸結束遲緩期,在培養(yǎng)6.3 h進入快速生長期;2019B6的生長情況比較平穩(wěn),2019B3、2019B5的生長曲線比較一致,在培養(yǎng)4 h內和2019B6的長勢相似,培養(yǎng)2 h內為遲緩期,培養(yǎng)2~4 h時生長開始增快,培養(yǎng)4 h后2019B3、2019B5快速增長,遠遠超過2019B6。
3 討論與結論
枯草芽孢桿菌于1835年被正式命名,隨著研究技術的進步,人們發(fā)現(xiàn)枯草芽孢桿菌是一個表型相似近緣種群(簡稱枯草群),其形態(tài)特征和生理生化特征是早期進行各種群分類與鑒定的基礎,16S rRNA基因含有豐富的信息量,已經(jīng)成為細菌系統(tǒng)發(fā)育和分類鑒定的標準,但是將分離篩選的菌株進行16S rRNA基因序列同源性檢索時發(fā)現(xiàn),多數(shù)菌株與GenBank數(shù)據(jù)庫中幾種芽孢桿菌的同源性達99%以上,這是因為枯草芽孢桿菌是一個表型相似近緣種群(簡稱枯草群),枯草群多個種類的16S rRNA基因序列相似性均在99.5%以上,目前NCBI(美國國家生物技術信息中心)核酸數(shù)據(jù)庫中,16S rRNA、gyrB、gyrA、rpoB、 purH等基因已涵蓋了枯草群的各個種,可利用這些基因進行相關菌株的鑒定和鑒別[18],因此本研究在分離篩選芽孢桿菌時,不僅進行了生理生化鑒定,還進行了分子生物學鑒定,在進行分子生物學鑒定時除進行16S rRNA測序建樹,各菌株還進行了gyrA、gyrB、rpoB和purH基因的測序同源比對,將傳統(tǒng)方法與分子生物學方法相結合,互為補充和驗證。
在生產(chǎn)過程中微生態(tài)制劑對于篩選益生菌尤其關鍵,種屬特異性是篩選合適益生菌的一個先決條件,不同來源的芽孢桿菌在不同動物胃腸道內的黏附繁殖能力及相關生理功能有所不同[16]。水產(chǎn)養(yǎng)殖戶在池邊將零星死亡的魚煮成肉糜自然發(fā)酵,待其無臭無味后潑灑使用,能有效地改善水質,池塘養(yǎng)殖魚未出現(xiàn)不良反應,說明發(fā)酵后的魚肉湯是安全的,可能含有能有效凈化水質的微生物,從這種發(fā)酵的魚肉湯中分離的菌株,可能來源于魚體或周邊環(huán)境,更適合在本區(qū)域池塘養(yǎng)殖使用。
本研究分離鑒定得到了6株芽孢桿菌,分屬于4種,即1株蠟樣芽孢桿菌,1株阿氏芽孢桿菌,2株解淀粉芽孢桿菌,2株貝萊斯芽孢桿菌,均具有產(chǎn)蛋白酶能力,其中4株具有產(chǎn)淀粉酶能力,6株菌在LB液體培養(yǎng)基中生長良好,為之后進一步研究其環(huán)境耐受性、體外抑菌試驗、水質凈化作用及對魚類的免疫作用研究奠定了基礎。
參考文獻:
[1]王秋菊,崔一喆. 微生態(tài)制劑及其應用[M]. 北京:科學工業(yè)出版社,2014:1-37.
[2]石水琴,蔣 雯,袁 林,等. 4株芽孢桿菌的分離鑒定與生物學特性分析[J]. 江蘇農業(yè)科學,2018,46(2):112-115.
[3]劉文亮,許 華,唐 楊,等. 飼料中補充蠟樣芽孢桿菌(Bacillus cereus)生物膜對凡納濱對蝦(Litopenaeus vannamei)生長、抗病力及其腸道微生物組成的影響[J]. 漁業(yè)科學進展,2017,38(4):90-98.
[4]王春迪,宋曉玲,張曉靜,等. 養(yǎng)殖水體中添加蠟樣芽孢桿菌PC465對凡納濱對蝦抗病力的影響[J]. 中國水產(chǎn)科學,2016,23(1):146-155.
[5]孫盛明,蘇艷莉,張武肖,等. 飼料中添加枯草芽孢桿菌對團頭魴幼魚生長性能、肝臟抗氧化指標、腸道菌群結構和抗病力的影響[J]. 動物營養(yǎng)學報,2016,28(2):507-514.
[6]張歡歡,王秀華,李 晨,等. 一株芽孢桿菌的分離鑒定及在生物絮團對蝦養(yǎng)殖中的應用[J]. 漁業(yè)科學進展,2016,37(2):111-118.
[7]Abasolo-Pacheco F,Campa-CórdovaI,Mazón-Suástegui J M,et al. Enhancing growth and resistance to Vibrio alginolyticus disease in catarina scallop (Argopecten ventricosus) with Bacillus and Lactobacillus probiotic strains during early development[J]. Aquaculture Research,2017,48(9):1-11.
[8]李衛(wèi)芬,沈 濤,陳南南,等. 飼料中添加枯草芽孢桿菌對草魚消化酶活性和腸道菌群的影響[J]. 大連海洋大學學報,2012,27(3):221-225.
[9]張峰峰,謝鳳行,趙玉潔,等. 枯草芽孢桿菌水質凈化作用的研究[J]. 華北農學報,2009,24(4):218-221.
[10]趙彩春,陳國明,張家學. 短小芽胞桿菌HLK8-1的分離鑒定及抑菌特性分析[J]. 微生物學雜志,2016,36(2):33-38.
[11]高艷俠,張德鋒,可小麗,等. 羅非魚源無乳鏈球菌腸道拮抗芽孢桿菌的篩選及其生物學特性[J]. 微生物學報,2019,59(5):926-938.
[12]張皎皎,馬富平,熊 波,等. 一株新型嗜水氣單胞菌拮抗菌的篩選及鑒定[J]. 西南大學學報(自然科學版),2017,39(12):18-23.
[13]李步先. 蝦蟹養(yǎng)殖池塘益生菌的分離篩選及降解特性初步研究[D]. 青島:中國海洋大學,2015:7.
[14]李洪鵬,李秋芬,張 艷,等. 淺海養(yǎng)殖環(huán)境復合生態(tài)凈化菌群的篩選及其凈化功能研究[J]. 漁業(yè)科學進展學進展,2009,30(2):46-53.
[15]巴翠玉,張林波,張培軍,等. 2株枯草芽孢桿菌的分離鑒定及特性研究[J]. 華南農業(yè)大學學報,2017,38(3):46-51.
[16]趙 東,牛廣杰,彭志云,等. 包包曲中5株枯草芽孢桿菌的分離與初步鑒定[J]. 中國釀造,2010 (8):65-68.
[17]曹鳳明,楊小紅,馬鳴超,等. 枯草芽孢桿菌近緣種群鑒定方法研究進展[J]. 微生物學通報,2014,41(5):968-974.
[18]王 佩,武亞方,盧江麗,等. 不同位置土壤中枯草芽孢桿菌產(chǎn)蛋白酶活力的比較[J]. 湖北畜牧獸醫(yī),2014,35(3):12-13.
