芒果葉綠體基因組密碼子使用偏好性分析
2021-09-14 02:37:23唐玉娟趙英黃國弟付海天宋恩亮李日旺金剛
熱帶作物學報 2021年8期
唐玉娟 趙英 黃國弟 付海天 宋恩亮 李日旺 金剛


摘 ?要:本研究利用CodonW、SPSS、Excel軟件對芒果葉綠體基因組55條蛋白編碼序列的密碼子開展了中性繪圖、PR2-plot、ENC-plot和最優密碼子分析。結果表明,芒果葉綠體基因的有效密碼子(ENC)偏好性不強,分布在35.38~56.56;GC3s含量26.29%,說明密碼子末位堿基以A/T結尾為主;ENC與GC3極顯著相關。相對同義密碼子分析,RSCU>1的密碼子有31個,其中,以A和T結尾的29個,表明A和T是基因組偏愛的密碼子。中性繪圖、PR2-plot和ENC-plot分析表明,芒果葉綠體基因組密碼子使用偏好受突變和選擇等因素共同作用的影響。通過高頻率密碼子和高表達優越密碼子相結合的方法進行分析,篩選出主要以A和T結尾的芒果葉綠體基因組最優密碼子19個,分別是GCT、TGT、GAA、TTT、GGT、CAT、ATT、AAA、TTA、CCT、CAA、AGA、CGA、CGT、AGT、TCT、ACT、GTA 和CTT。
關鍵詞:芒果;葉綠體基因組;密碼子偏好性;最優密碼子
Abstract: Codon usages of 55 chloroplast genes in mango were analyzed to obtain to the results of neutrality plot, PR2-plot ENC-plot and optimal codons by using CodoW, SPSS and SPSS softwares. The results showed that the effective number of codons (ENC) ranged from 35.38 to 56.56, indicating that the condon bias was weak. GC base pair content in the third condon position of chloroplast genomes in mango was 26.29%, indicating that the third base in the codon was terminated by A/T. Significant correlation was found between ENC and GC3.There were 31 codons with relative synonymous codon usage greater than 1, of which 29 codons ending with A and T, indicating that the third codon preferred A and T.According to neutrality plot analysis, PR2-plot and ENC-plot analysis, mutational pressure combined with selection influenced the pattern of synonymous codon usage across genes in mango chloroplast genomes. In addition 19 codons were affirmed as the optimal codons by using analysis of high-frequency codons and the high expression codons, ending with A and T. There were GCT, TGT, GAA, TTT, GGT, CAT, ATT, AAA, TTA, CCT, CAA, AGA, CGA, CGT, AGT, TCT, ACT, GTA and CTT.
Keywords: Mango; chloroplast; codon usage bias; optimal codons
遺傳密碼又稱密碼子,編碼同種氨基酸的密碼子稱為同義密碼。同義密碼子提供了基因表達和蛋白質進化間的聯系,維持著生物體遺傳密碼的功能完整性。密碼子的使用頻率不同,有的密碼子使用頻率高于其他同義密碼子,形成了密碼子偏好性[1]。不同物種密碼子偏性不同,但同一種物種或者親緣關系相近的物種間卻使用了相似的偏好性[2]。密碼子偏好性的使用在基因表達水平、共翻譯蛋白質折疊、mRNA的穩定性等[3],反映了翻譯優化中突變偏好與自然選擇的平衡[4],因此研究物種密碼子偏好性,為探究物種的進化模式與其在系統發育史上的地位提供依據。
葉綠體是植物特有的細胞器,具有高度的保守性及較低的進化率,是研究植物條形碼最有潛力的基因序列[5-6],在植物系統進化、物種鑒定等方面廣泛應用[7-8]。完整的葉綠體基因組為認識進化和自然選擇提供機理性的啟示。不同基因組有其特有的密碼子使用模式,密碼子使用模式的分析有助于理解生物體適應性及其在系統發育進化史的地位[9],有益于基因表達水平的調控與基因的改造[10]。
芒果(Mangifera indicate L.)是著名的熱帶水果,享有“熱帶水果之王”之美稱[11]。外觀秀麗、果香四溢,含有豐富的維生素、蛋白質、胡蘿卜素等,營養價值極高,深受大眾的喜愛,是南方熱區果農增值創收的主要水果之一[12]。目前為止尚未發現芒果密碼子使用偏好性相關方面的研究報道,本研究通過對芒果葉綠體基因組密碼子組成及相關參數進行分析,首次闡明芒果密碼子使用偏好性的特點,確定芒果葉綠體基因組的最優密碼子及密碼子偏好性發生的因素,為芒果資源保護、性狀改良及葉綠體基因組的應用和研究提供參考依據。
1 ?材料與方法
1.1 ?基因序列
芒果葉綠體基因組GenBank登錄號為CM021858(https://www.ncbi.nlm.nih.gov/),共60個基因序列。為保證結果的準確性[13],剔除重復且長度小于300 bp的編碼序列,選擇以ATG為起始密碼子,以TAA、TAG、TGA為終止密碼子的序列,最終選出55條序列進行分析。
1.2 ?中性繪圖分析
統計密碼子3個位置上的GC含量,以GC12為縱坐標,GC3為橫坐標繪制散點圖進行相關性分析。
1.3 ?相對同義密碼子使用度分析
利用CodonW軟件對相對同義密碼子使用度(relative synonymous codon usage, RSCU)進行分析。RSCU表示一個密碼子的實際使用頻率與無偏好性時理論使用頻率間的比值。RSCU<1時,表示該密碼子的使用頻率低于其他同義密碼子;RSCU>1時,表示該密碼子的使用頻率高于其他同義密碼子;當RSCU=1時表示該密碼子沒有偏好性[14]。
1.4 ?ENC-plot繪圖分析
以有效密碼子ENC(effective number of condon)為縱坐標,GC3s為橫坐標,繪制散點圖,計算公式:ENC=2+GC3s+29/[GC3s2+(1-GC3s)2]。結合ENC比值頻率對差異進行量化分析[15]。
1.5 ?PR2-plot繪圖分析
計算各密碼子第3位上的A、T、C、G的含量,以A3/(A3+T3)為縱坐標,以G3/(G3+C3)為橫坐標進行PR2的偏倚分析。
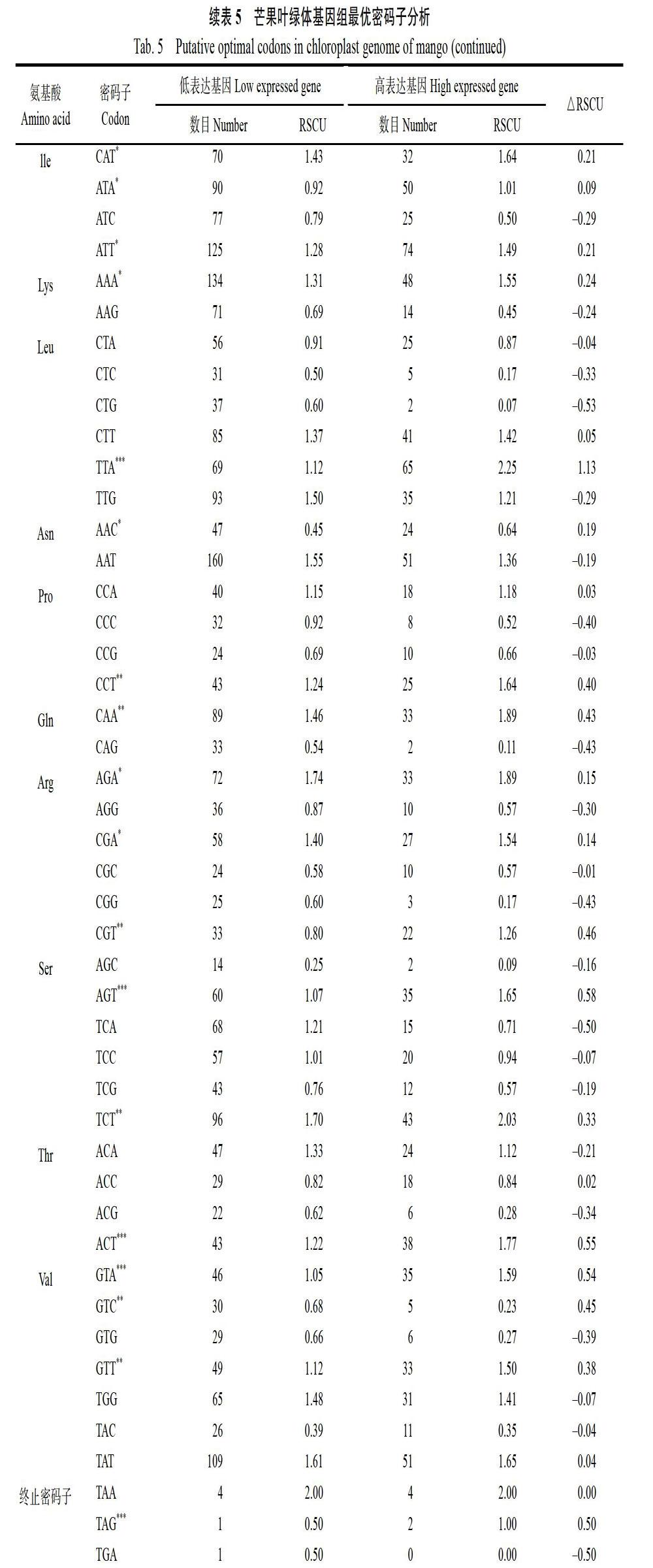
1.6 ?最優密碼子分析
最優密碼子以芒果ENC值為偏性參考標準,選擇最低和最高10%的基因構建高低偏性庫,分別計算兩組的RSCU值與△RSCU值,將RSCU>1的密碼子確定為高頻率密碼子,△RSCU>0.08的密碼子定義為高表達密碼子。同時滿足上述2種條件的密碼子定義為最優密碼子[16]。
2 ?結果與分析
2.1 ?密碼子使用的總體特征
對55條芒果葉綠體基因組的密碼序列的堿基組成進行分析(表1),葉綠體基因組密碼子3個位置上的GC平均含量為37.94%。從密碼子不同位置的堿基組成來看,GC在密碼子的分布不均勻,GC1、GC2、GC3含量分別為47.14%、39.73%和29.35%,第3位上的GC含量低于前兩位,分布趨勢為GC1>GC2>GC3。GC3s的含量26.29%,表明密碼子中的第3位堿基以A/T結尾為主。
ENC表示偏性的強弱,其理論取值范圍為21~61,其值越小表示同義密碼子的偏性越強,反之同義密碼子的偏性越弱。當ENC值為20時,表示同義密碼子完全偏倚;當ENC值為60時,表示同義密碼子不偏倚[13]。芒果葉綠體基因組編碼序列的ENC值均(48.14)值大于40,分布于35.38~56.56之間,由此可以判斷芒果葉綠體基因組密碼子的偏性不強。
芒果葉綠體基因密碼子參數的相關分析顯示GCall和GC1、GC2、GC3極顯著相關(表2)。GC1和GC2、GC3顯著相關,說明芒果葉綠體基因密碼子3個位置上的堿基組成相似。ENC值與GC2顯著負相關,與GC3極顯著相關,表明第2、3位上的堿基組成,特別是第3位對密碼子的偏好性影響較大。
2.2 ?中性繪圖分析
芒果葉綠體基因組GC12的取值范圍在0.33~0.56之間,GC3的取值范圍在0.18~0.32之間,基因全部落在中線對角線上方(圖1)。GC12與GC3的相關系數0.279(表2),雙尾檢測相關性不顯著,回歸系數(曲線斜率)為0.3544,相關性較弱。結合圖1和表2表明芒果葉綠體基因組密碼子第1、2位和第3位堿基之間關聯性不大,自然選擇在芒果葉綠體基因組密碼子偏好使用中起主要作用。
2.3 ?ENC-plot繪圖分析
ENC-plot繪圖以ENC比值[(預期ENC值~實際ENC值)/預期ENC值]和ENC比值頻數判斷偏好性影響因素,揭示ENC與GC3s的關系,反映堿基組對密碼子偏好性的影響程度[15-19]。基因坐標點離標準曲線的距離近且數量較多說明偏好性主要受到突變的影響;反之說明偏好性受選擇的影響較大。圖2結合表3顯示60%共33個基因分布在–0.05~0.05之間,這33個基因的實際ENC值與預期ENC值接近,與標準曲線的距離較近,;分布在–0.05~0.05之外的基因有22個,這22個基因與預期ENC值較遠,在標準曲線較遠的位置。說明芒果基因組密碼子偏好性與GC3的差異有關,突變對芒果葉綠體基因組密碼子偏好性的影響較大。
2.4 ?PR2-plot繪圖分析
PR2-plot繪圖通過分析密碼子第3位堿基的組成關系,揭示密碼子使用偏好性的影響因素。由圖3可以看出芒果基因在4個區域內分布不均勻,多數分布于平面圖的左下方。T和C在密碼子第3位出現的頻率較高,G與C、A與T的使用頻率不相等,T的使用頻率高于A,C的使用頻率高于G,嘧啶的使用頻率高于嘌呤,說明突變不是影響芒果密碼子使用偏好性的唯一因素,選擇等其他因素對芒果密碼子使用偏好性也產生一定的影響。
2.5 ?相對同義密碼子使用度分析
利用codonW 軟件對芒果55條基因序列的RSCU值進行計算(表4)。結果顯示RSCU>1的密碼子共31個,出現頻率相對較高的是以A(13個)或T(16個)結尾的密碼子,是芒果基因組偏愛;以G(1個)或C(1個)結尾的密碼子出現的頻率較低且RSCU值多小于1,說明這些是基因組非偏好的密碼子。
2.6 ?最優密碼子分析
芒果葉綠體基因組,以△RSCU≥0.08為標準確定了24個高表達密碼子,以A結尾的8個,以T結尾的12個,以C結尾的3個,以G結尾的1個。結合芒果葉綠體基因的相對同義密碼子使用度和同義密碼子相對使用度(表4和表5),確定19個最優密碼子,分別是GCT、TGT、GAA、TTT、GGT、CAT、ATT、AAA、TTA、CCT、CAA、AGA、CGA、CGT、AGT、TCT、ACT、GTA、CTT。其中以A結尾的7個,以T結尾的12個,暗示芒果葉綠體基因組偏愛使用以A和T堿基結尾的密碼子。
3 ?討論
隨著分子生物學的飛速發展,測序技術的持續更新,基因組數據的應用為密碼子的偏性研究提供了支持。研究發現GC含量可反映密碼子使用偏性突變的整體趨勢[20],密碼子第3位堿基GC3含量可作為密碼子偏好性分析的主要依據[21]。本研究通過對篩選出的芒果葉綠體基因組的55條密碼序列進行分析發現,GC3s含量為26.29%,暗示密碼子偏好使用末位堿基為A/T的密碼子。結果與劍麻[22]、杉木[23]偏好使用末位堿基為A/T的密碼子相似。ENC可判斷基因表達量的高低,當其大于35時基因表達水平較低,在編碼氨基酸時,對密碼子的選用偏好性也弱。芒果葉綠體基因組ENC均值為48.14,表明葉綠體基因密碼子偏好性不強。ENC越小,基因的密碼子使用偏好性越強,越偏好使用A/T結尾的密碼子[24],這與本實驗RSCU等數據揭示的芒果葉綠體基因組第3位密碼子富含A/T的結論相一致。突變和自然選擇是影響密碼子偏好性使用的主要因素,本研究通過中性繪圖分析顯示,選擇是密碼子偏好性的主要影響因素;通過ENC-plot繪圖分析則發現,突變是密碼子偏好性的主要影響因素,本研究認為芒果葉綠體基因組密碼子的偏好性受突變和選擇2種因素的影響應該是相對均衡的。
同義密碼子的偏好使用影響基因的表達,最優密碼子可以提高翻譯的效率和準確率[25]。有研究認為,雙子葉植物偏好使用A/T結尾的密碼子,單子葉植物偏好使用G/C結尾的密碼子[26],但也有不同的觀點,如有偏好使用A/T結尾的單子葉植物如糜子[27]、霍山石斛[28]等。芒果屬于雙子葉植物,本研究通過高表達優越密碼子和高頻密碼子相結合的方法篩選出的19個最優密碼子均以A或T結尾,表明芒果葉綠體基因組偏愛使用以A和T堿基結尾的密碼子,結論與雙子葉偏好使用A/T結尾的密碼子類似。
本研究系統的對芒果葉綠體基因組密碼子的偏好性及影響因素進行了研究,發現芒果葉綠體基因組偏好使用以A/T結尾密碼子;突變和選擇均衡的影響密碼子的偏好性使用;研究還確定了19個芒果葉綠體基因組最優密碼子。研究結果為今后芒果分子系統進化、遺傳轉化、葉綠體基因工程等方面的研究奠定基礎,為芒果在葉綠體基因組水平上進行分類、性狀改良等提供參考依據。
參考文獻
Grantham R, Gautier C, Gouy M. Codon frequencies in 119 individual genes confirm consistent choices of degenerate bases according to genome type[J]. Nucleic Acids Research, 1980, 8(9): 1893-1912.
Grantham R, Gautier C, Gou M. et al. Codon catalog usage and the genome hypothesis[J]. Nucleic Acids Research, 1980, 8(1): 49-62.
文 ?言. 川桑密碼子使用模式與密碼子分析工具的開發[D]. 重慶: 西南大學, 2017.
Wu X M, Wu S F, Ren D M, et al. The analysis method and progress in the study of codon bias[J]. Hereditas, 2007, 29(4): 420-426.
Gielly L, Taberlet P. The use of chloroplast DNA to resolve p;ant phylogeny: noncoding versus rbcL sequences[J]. Molecular Biology and Evolution, 1994(11): 769-777.
Shaw J, Lickey E B, Beck J T, et al. The tortoise and the hare:relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis[J]. American Jourmal of Botany, 2005(92): 142-166.
李 ?汐, 祝 ?銘, 孫延霞, 等. 基于葉綠體rps16基因和核基因ITS片段研究肉蓯蓉屬系統位置[J]. 植物科學學報, 2012, 30(5): 431-436.
李丹丹, 郭水良, 于 ?晶, 等. 基于4個葉綠體基因識別蓑蘚屬(Macromitrium)植物的可行性研究[J]. 植物科學學報, 2013, 31(1): 23-33.
張月榮. 禾本科葉綠體基因組密碼子使用模式及紫莖澤蘭葉綠體RNA編輯分析[D]. 楊凌: 西北農林科技大學, 2013.
劉 ?柱, 胡新文, 劉志昕. 基因及表達調控中遺傳密碼子選擇的偏愛性[J]. 生命科學研究, 2011, 5(3): 106-109.
付海天, 趙 ?英. 芒果SRAP-PCR反應體系的優化與退火溫度的篩選[J]. 福建農業學報, 2014, 29(4): 345-349.
唐瑩瑩, 黃國弟, 陳永森, 等. 杧果新品種‘桂熱杧3號對炭疽病和細菌性黑斑病的抗性評價初報[J]. 中國南方果樹, 2020, 49(3): 35-39, 45.
Rosenberg M S, Subramanian S, Kumar S. Patterns of transitional mutation biases within and among mammalian genomes[J]. Molecular Biology and Evolution, 2003, 20(6): 988-993.
馮海悅, 張 ?羽, 吳正常, 等. 13種哺乳動物MUC4基因密碼子使用模式分析[J]. 農業生物技術學報, 2018, 26(9): 1546-1556.
Sueoka N. Near homogeneity of PR2-bias fingerprints in the human genome and their implications in phylogenetic analyses[J]. Journal of Molecular Evolution, 2001, 53(4-5): 469-476.
原曉龍, 李云琴, 張勁峰, 等. 降香黃檀葉綠體基因組密碼子偏好性分析[J/OL]. 廣西植物, 2021, 41(4): 622-630.
劉 ?慧, 王夢醒, 岳文杰, 等. 糜子葉綠體基因組密碼子使用偏性的分析[J]. 植物科學學報, 2017, 35(3): 362-371.
Sueoka N. Directional mutation pressure and neutral molecular evolution[J]. Proceedings of the National Academy of Sciences of the United States of America, 1988, 85(8): 2653-2657.
李慧姬, 吉雪花, 朱冉冉, 等. 10種植物PSY基因密碼子使用偏好性分析, 西北農業學報, 2020,29(2): 276-284.
Zhou M, Long W, Li X. Analysis of synomous codon usage in chloroplast genome of Populus alba[J]. Journal of Forestry Research, 2008, 19(4): 293-297.
馮瑞云, 梅 ?超, 王慧杰, 等. 籽粒莧葉綠體基因組密碼子偏好性分析[J]. 中國草地學報, 2019, 41(4): 8-15.
金 ?剛, 覃 ?旭, 龍凌云, 等. 劍麻葉綠體基因組編碼序列密碼子的使用特征[J]. 福建農林大學學報, 2018, 47(6): 705-710.
鄭薇瑋, 陳金慧, 郝兆東, 等. 杉木等5種針葉樹葉葉綠體密碼子偏好性分析[J]. 分子植物育種, 2016, 14(5): 1091-1097.
雷 ?慧, 李 ?鴿, 王娜玉. 金蓮花葉綠體基因組密碼子偏好性分析[J]. 山西農業科學, 2019, 47(8): 1300-1305, 1445.
Sharp P M, Matassi G. Codon usage and genome evolution[J]. Current Opinion in Genetics & Development, 1994, 4(6): 851-860.
Wang L J, Roossinck M J. Comparative analysis of expressed sequences reveals a conserved pattern of optimal codon usage in plants[J]. Plant Molecular Biology, 2006, 61(4-5): 699-710.
劉 ?慧, 王夢醒, 岳文杰, 等. 糜子葉綠體基因組密碼子使用偏性的分析[J]. 植物科學學報, 2017, 3(3): 362-371.
謝析穎, 李 ?蓉, 林玉玲, 等. 霍山石斛轉錄因子MYB1R1密碼子選用偏好性分析[J]. 分子植物育種, 2017, 15(5): 1683-1690.
責任編輯:白 ?凈
