雅礱江官地水庫環境因子和浮游植物群落結構的時空變化及相互關系
2021-09-25 02:22:22李天才甘維熊姚維志蘇勝齊
西南農業學報 2021年8期
李 鋒,李天才,甘維熊,姚維志,付 梅,熊 波,蘇勝齊*
(1.西南大學水產學院,重慶 400700;2.西南大學漁業資源環境研究中心,重慶 400700;3.雅礱江流域水電開發有限公司,四川 成都 610000)
【研究意義】雅礱江官地水庫建于2013年,處于西昌市與鹽源縣交界處,上游連接雅礱江錦屏大河灣,下游毗鄰二灘水庫[1],是雅礱江下游由人煙稀少地區轉入人口密集城鎮的樞紐,也是長江上游重要生態屏障和水源涵養地的組成部分。因此,掌握官地水庫水質狀況和浮游植物群落結構可為管理、保護和改善區域水生態環境提供基礎資料和科學依據,有助于下游作為飲用水源的二灘水庫水質變化分析和提前管理。【前人研究進展】水體理化指標和營養成分含量是評判水庫水質等級的直接數據來源。浮游植物是水庫生態系統中重要初級生產者,是水庫生態系統食物網的基礎,其密度、結構直接影響高級捕食者的種群結構,繼而對整個生態系統產生重要影響[2-3]。藻類多數為單細胞生物,對環境因子的變化極為敏感,故而其種類組成、密度分布、多樣性及特殊種類可反映水體環境因子及污染狀況,是評價水庫生態系統的重要指標[4-6]。研究[7-8]表明,總體上湖泊型水庫污染程度高于河道型水庫,下游水庫又高于上游水庫;養殖型、灌溉型水庫污染程度高于發電型水庫,臨近城鎮或工業園區的水庫污染程度高于山區源頭水庫。關于環境因子和浮游植物的調查多數是因為水庫已出現不同程度的污染,前人研究目的大多為掌握水庫基本情況,提出針對性治理方案[7-10]。【本研究切入點】官地水庫地處偏僻,屬于山區源頭發電-河道型水庫[11],多年觀察初步判斷其水質良好。而本研究參照成熟的水庫環境因子和浮游植物調查方法,歷經1年4個季度的調查,旨在系統掌握官地水庫水質狀況和浮游植物群落結構。【擬解決的關鍵問題】分析存在的官地水庫環境因子和浮游植物群落的時空變化特征,并梳理相互之間的關系,為保護水質、預防惡化提供科學管理依據。
1 材料與方法
1.1 采樣時間與斷面
官地庫區回水長度近60 km,庫區河道狹窄,并無大的支流匯入,因此僅在干流主要位置設置采樣斷面即可。經現場勘察、分析,沿江自上游到下游設置A/B/C 3個采樣斷面(表1和圖1)。每個斷面采樣時采集左右兩岸水體和浮游植物樣本,以兩岸平均數作為調查結果。分別于2018年冬季(12月)和2019春季(3月)、夏季(7月)、秋季(10月)完成官地庫區周年水質及浮游植物采樣調查。
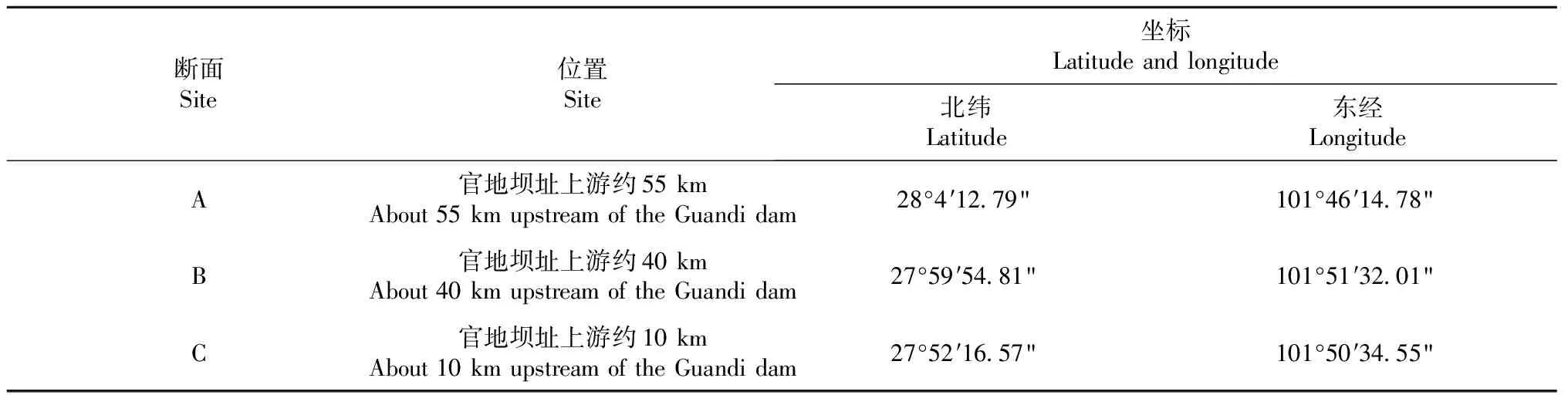
表1 官地庫區采樣斷面設置
1.2 水樣采集和環境因子測定
水體理化指標測定按照《水環境檢測規范》(SL 219-98)和《水庫漁業資源調查規范》(SL 167-96)進行。水溫(WaterTemperature, WT)和溶解氧(Dissolved Oxygen,DO)使用YSI55型溶氧儀現場測定;pH值使用便攜式pH計現場測定;電導率(Conductivity, CT)使用DOB33A型電導儀現場測定;透明度(Transparency, SD)使用塞氏盤直接現場測定。水體硬度(Hardness of Groundwater,HG)、化學需氧量(Chemical Oxygen Demand,COD)、總氮(Total Nitrogen, TN)、總磷(Total Phosphorus, TP)等環境因子參照《水和廢水監測分析方法(第四版)》[12]在西南大學漁業資源與環境中心實驗室進行測定。
1.3 浮游植物樣本采集和分析
用2 L有機玻璃采水器分別定量表層、30和50 cm水層水樣,均勻混合后取1 L水體作為浮游植物分析樣本。浮游植物水樣加15 mL 1.5%(V/V)魯哥(Lugol)氏液進行固定后帶回實驗室進行分析。浮游植物水樣用浮游生物沉淀器在實驗室沉淀24 h后吸出上清液,剩余20~50 mL濃縮水樣倒入浮游生物標本瓶中以待定量鑒定。定量鑒定時將濃縮水樣搖勻,取0.1 mL置于計數框內,在10×40倍鏡下觀察、鑒定、計數浮游植物。每瓶樣本計數取兩次觀測的平均值,同一樣本的兩次定量鑒定值不大于平均值的5%為有效觀測,否則繼續觀察直至符合要求。浮游植物種類鑒定主要參照《中國淡水藻類》[13]、《中國內陸水域常見藻類圖譜》[14]、《中國常見淡水浮游藻類圖譜》[15]等資料。
1.4 數據處理
采用 Excel 2016對水體環境因子和浮游植物群落結構進行匯總分析、作圖;采用SPSS 22.0進行Pearson相關性分析;采用Conaco 4.5進行聚類、冗余分析并作圖。
ρ=10nv
式中,ρ為浮游植物密度(個/L),n為兩次觀測到藻類數量的平均值,v為1 L水樣經沉淀濃縮后的體積(mL);
Pi=Ni/N
式中,Pi為第i個類群在群落中的數量占比,N為群落個體總數,Ni為第i個類群的數量;
H′=-∑Pi×log2Pi
式中,H′為香農威納指數(Shannon-WienerIndex);
J′=H′/log2S
式中,J′為均勻度指數(Pielou’s Evenness Index),S為群落種群數量;
式中,D′為辛普森多樣性指數(Simpson’s diversity index);
Y=Pi×fi
式中,Y為種群優勢度(大于0.02即為優勢種群),fi為第i個種群出現的頻率。
2 結果與分析
2.1 水體環境因子
2.1.1 水體理化性質及主要營養成分 官地庫區水溫空間上整體表現為上游低于下游;季節上則表現為冬季轉向夏季過程中水溫逐漸升高,而后再降低的循環;全年水溫10.9~22.6 ℃,平均值14.9 ℃。溶解氧空間上僅冬季各斷面差異較大,表現為上游高于下游,其它季節各斷面無明顯差異;季節上則整體表現為冬季逐漸降低至秋季后再升高的循環;全年溶解氧為8.5~16.2 mg/L,平均值為10.8 mg/L。冬、春和秋季各江段pH值均無明顯差異,在8.35~8.46小幅變化,僅秋季各江段明顯高于其它季節;全年pH呈弱堿性,變幅8.35~8.97,平均值8.51。透明度在空間上總體表現為自上而下逐漸升高的趨勢,僅秋季A斷面高于其它斷面;季節上均表現為自秋季逐漸降低至夏季后再升高的循環,其中夏季遠低于其它季節;全年透明度為21~534 cm,平均值為238 cm。硬度在空間上各斷面無大的差異,季節上則均表現為春季逐漸降低至秋季,而后再升高的循環;全年硬度為2.5~6.0 mmol/L,平均值為4.0 mmol/L。秋、冬和春季各斷面水體電導率均無明顯差異,在239.0~263.2 μs/cm小幅變化,僅夏季各斷面水體電導率明顯較低,全年平均電導率為243.4 μs/cm。春、夏和秋季各季節內各斷面COD均無明顯差異,僅冬季各斷面COD有較大差異;從季節上COD則均表現為自春季逐漸升高至冬季,而后轉為降低的循環;全年COD為1.3~10.6 mg/L,平均值為5.1 mg/L。
官地庫區秋、冬和春季各季節內各斷面總氮含量無明顯差異,僅夏季為自上而下快速升高;從季節上均表現為自夏季逐漸升高至冬季,而后再下降的循環;全年總氮為0.10~0.94 mg/L,平均值為0.54 mg/L。總磷從空間上無明顯變化趨勢,季節上總體表現為自春季逐漸升高至秋季,而后再下降的循環;全面總磷含量為0.006~0.085 mg/L,平均值為0.042 mg/L。氮磷比僅在冬季表現為自上而下逐漸降低,其它季節均為自上而下逐漸升高的趨勢;從季節上僅A斷面表現為自冬季逐漸降低值秋季,而后再升高的循環趨勢,其它斷面均為自春季逐漸降低至秋季而后再升高的循環趨勢;全年為2.3~91.6,平均值為29.4。
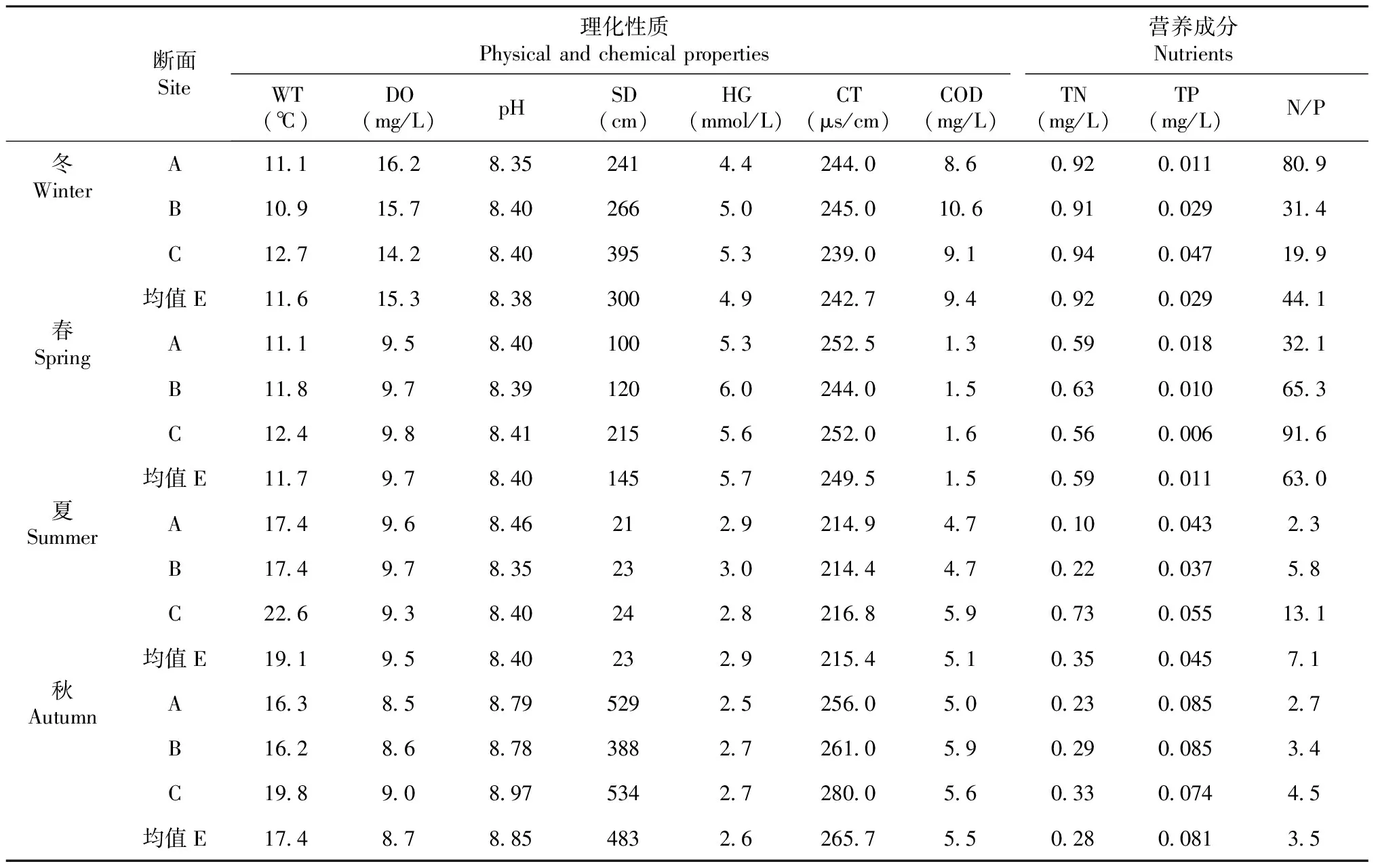
表2 官地庫區水體理化性質及主要營養成分
2.1.2 水體環境因子間Pearson相關性 環境因子中除pH和總磷外均與水溫呈負相關關系,其中溶氧、總磷、氮磷比與水溫相關性顯著,而硬度與之達極顯著相關;化學需氧量、總氮與溶氧均呈極顯著正相關關系;pH與透明度、硬度、電導率、總磷的相關性均達顯著,其中與電導率、總磷為極顯著正相關關系;透明度與pH、電導率、總磷均呈顯著正相關關系,且前兩項更達極顯著程度;硬度與水溫、pH、總磷均呈負相關關系,而與總氮、氮磷比均呈正相關關系,其中與水溫、總氮相關性達極顯著;電導率與pH、透明度均呈正相關相關,且后者相關性達極顯著;總氮與溶氧、硬度均呈正相關關系,且后者相關性達極顯著;總磷與水溫、pH、透明度均呈負相關關系,與硬度、氮磷比均呈正相關關系,其中與pH、硬度、氮磷比相關性均達極顯著;氮磷比與水溫、硬度、總磷分別呈顯著負相關、極顯著正相關、極顯著負相關關系。
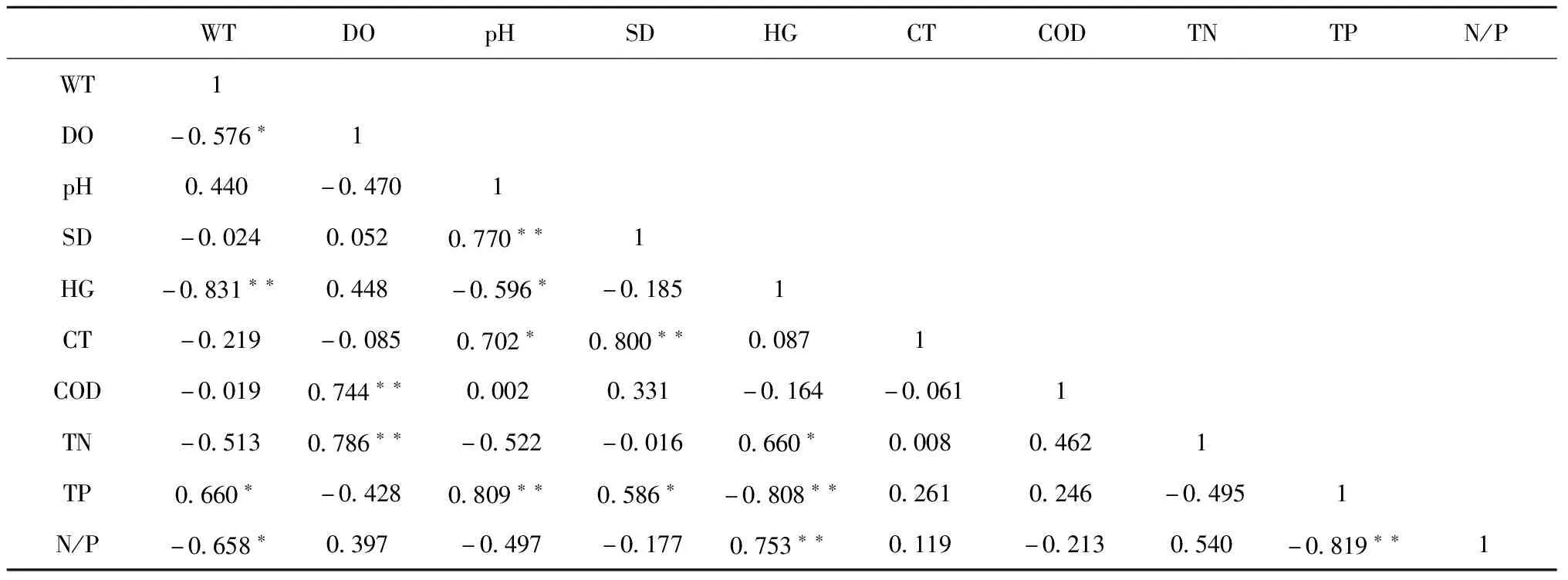
表3 水體環境因子間Pearson相關性
2.2 浮游植物群落結構
2.2.1 浮游植物群落多樣性指數 官地水庫浮游植物群落香濃威納指數均高于1(圖2),其中僅B斷面的冬、春季低于2,且明顯低于同季節的其它斷面,而冬季A和C、夏季A和B斷面高于3;全年香農維納指數變幅為1.01~3.80,平均值為2.50。均勻度指數空間上均表現為B斷面最低,C斷面最高,僅夏季完全相反;季節上A斷面表現為自春季逐步升高至冬季后轉而降低的循環,B斷面則表現為自春季升高至夏季后轉而逐步降低的循環,C斷面則為春夏季向秋冬季升高的趨勢;全面均勻度指數變幅為0.27~0.78,平均值為0.59。辛普森指數空間上變化趨勢與均勻度指數相同;季節上A和B斷面均為夏季逐步降低至春季后轉為升高的循環,而C斷面春季顯著高于其它季節;全年辛普森指數變幅為0.25~0.88,平均值為0.67。綜合比較,C斷面冬春季多樣性指數均顯著低于其它斷面和季節,秋季各斷面上各類多樣性指數均無顯著差異。
2.2.2 浮游植物群落組成及藻類密度 官地庫區全年鑒定出6門97種(屬)藻類(圖3),其中硅藻門40種(屬),綠藻門33種(屬),裸藻門10種(屬),藍藻門7種(屬),甲藻門4種(屬),隱藻門3種(屬);冬季共有6門42種(屬),春季共有6門45種(屬),夏季共有6門56種(屬),秋季3門16種(屬);總體來看,硅藻門和綠藻門種(屬)數在全年和各季度均為最多,而秋季藻類門類數和種(屬)數均顯著低于其它季節。全年藻類平均密度為139.8×104cell/L(圖4),空間上藻類密度自上游而下逐漸升高,A、B、C斷面藻類密度分別為98.6×104、134.7×104、186.2×104cell/L;冬至秋季藻類密度分別為25.3×104、194.83×104、244.53×104、94.8×104cell/L,表現為自夏季降低至冬季后升高的循環。
2.2.3 浮游植物群落優勢門類 從全年視角來看(圖5-a),官地庫區綠藻數量占比達42.5%,遠高于其它類群,是絕對優勢門類;藍藻和硅藻數量占比分別達24.2%和19.3%,是亞優勢門類。從季節視角來看(圖5-b),綠藻數量除春季占比低以外,其它季節占比均為最高,是絕對優勢門類;而藍藻僅在春季占比最高,為絕對優勢門類;硅藻僅在夏季占比略低于10%,其它季節占比在25%上下小范圍波動,為亞優勢門類。從斷面視角來看(圖5-c),冬季A、C斷面優勢門類均為綠藻和硅藻,B斷面藍藻為絕對優勢門類;春季A、B斷面絕對優勢門類分別為硅藻和藍藻,C斷面優勢門類為硅藻和藍藻;夏季A斷面隱藻和綠藻為優勢門類,B、C斷面綠藻均為絕對優勢門類;秋季各斷面均以綠藻為絕對優勢門類;總體上綠藻為優勢門類的斷面和季度最多,硅藻次之。
2.2.4 浮游植物群落優勢種群 從全年視角(表4)來看,優勢種群有26種(屬),其中硅藻和綠藻門各有8種(屬),藍藻門有4種(屬),裸藻和隱藻門各有2種(屬),甲藻門僅1種(屬);而優勢度大于0.1的僅有硅藻門美麗星桿藻,綠藻門湖生卵囊藻、卵形衣藻、小球衣藻、小球藻,藍藻門不定微囊藻、水華微囊藻,隱藻門卵形隱藻,其中小球衣藻優勢度最高,達0.713。
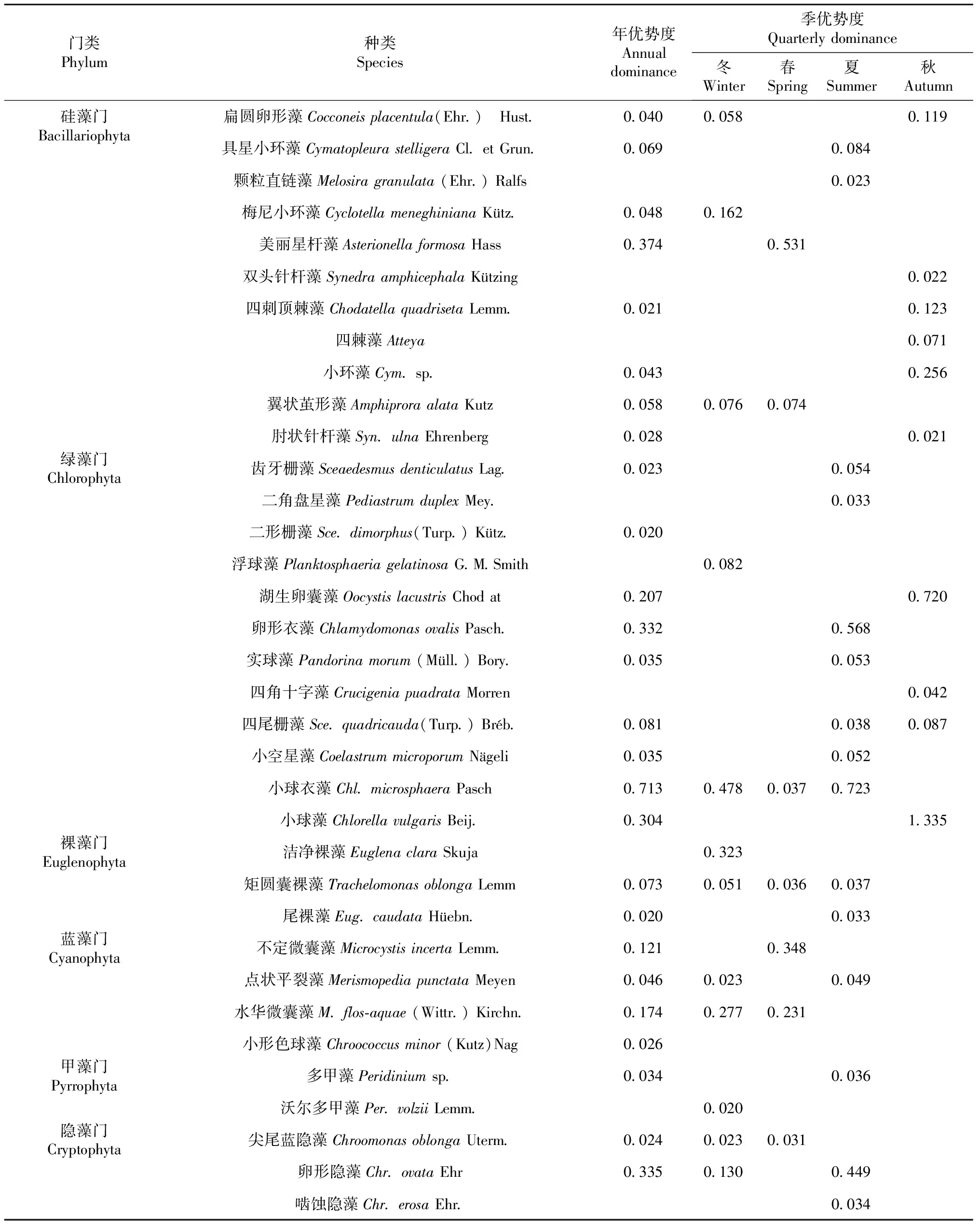
表4 不同時間尺度下各藻類種群優勢度
從季節視角來看,冬季優勢種群有12種(屬),各門類分布較為均勻;春季優勢種群有7種(屬);夏季優勢種群有15種(屬),主要集中在綠藻門;秋季優勢種群有10種(屬),僅分布在硅藻和綠藻門。優勢度大于0.1,冬季有硅藻門梅尼小環藻、綠藻門小球衣藻、裸藻門潔凈裸藻、藍藻門水華微囊藻、隱藻門卵形隱藻;春季有硅藻門美麗星桿藻、藍藻門不定微囊藻和水華微囊藻;夏季有綠藻門卵形衣藻和小球衣藻、隱藻門卵形隱藻,秋季有硅藻門扁圓卵形藻、四刺頂棘藻、小環藻以及綠藻門湖生卵囊藻、小球藻。總體上看,綠藻門小球衣藻在冬春夏季均為優勢種群,而綠藻門小球藻在秋季優勢度為全年最高。
從斷面視角(表5)來看,冬季A斷面綠藻門小球衣藻為絕對優勢種群,B斷面藍藻門水華微囊藻為絕對優勢種群,C斷面硅藻門翼狀繭形藻、綠藻門浮球藻和小球衣藻、裸藻門潔凈裸藻數量占比均超過10%;春季A、B、C斷面絕對優勢種群分別為硅藻門美麗星桿藻、藍藻門不定微囊藻、藍藻門水華微囊藻;夏季A斷面優勢種群為隱藻門卵形隱藻,B斷面優勢種群為藍藻門點狀平裂藻,C斷面優勢種群為藍藻門卵形衣藻和小球衣藻;秋季綠藻門小球藻和湖生卵囊藻為各斷面優勢種群,其中小球藻為B斷面絕對優勢種群。不同季節各斷面中,占比超過5%且出現次數超過3次的藻類分別有硅藻門美麗星桿藻和綠藻門湖生卵囊藻、小球衣藻、小球藻,其中小球衣藻高達5次。

表5 各斷面不同季節占比大于5%的藻類種群
2.3 浮游植物群落結構與環境因子相關性分析
2.3.1 浮游植物群落多樣性指數與環境因子冗余分析 第一軸和第二軸特征值分別為0.897和0.089(圖6,Lengths of gradient=0.184),兩軸對浮游植物群落多樣性的解釋量為98.6%。香農威納指數、均勻度指數和辛普森指數均與化學需氧量、透明度、電導率、溶解氧呈顯著正相關性,與水溫呈顯著負相關性;種群數量與氮磷比、硬度、總氮、溶解氧均呈顯著正相關性,與總磷、水溫、pH均呈極顯著負相關性。硬度、水溫、化學需氧量和總磷是驅動多樣性指數變化的主要環境因子。
2.3.2 浮游植物門類與環境因子冗余分析 第一軸和第二軸特征值分別為0.403和0.262(圖7,Lengths of gradient=1.209),兩軸對浮游植物群落門類的解釋量為66.5%。硅藻門和綠藻門與透明度、pH、總磷、電導率均呈正相關性,與溶氧、總氮、硬度、氮磷比均呈負相關性,而裸藻門與硅藻門和綠藻門幾乎相反;藍藻門與水溫、化學需氧量均呈顯著負相關性,與氮磷比呈顯著正相關性,而甲藻門幾乎相反;隱藻門與化學需氧量、溶氧均呈顯著正相關性,與電導率呈顯著負相關性。
2.3.3 浮游植物優勢種群與環境因子冗余分析 第一軸和第二軸特征值分別為0.356和0.160(圖8,Lengths of gradient=3.762),兩軸對浮游植物群落多樣性的解釋量為51.6%。美麗星桿藻與除電導率、透明度外的環境因子相關性均達到顯著,而湖生卵囊藻與之相似;卵形衣藻僅與pH、化學需氧量、透明度、電導率等均呈顯著負相關性,與其它環境因子相關性很弱;小球藻僅與總磷、pH、化學需氧量、透明度、電導率等均呈顯著負相關性;小球衣藻與總磷、pH、化學需氧量外的環境因子相關性均達顯著;不定微囊藻、點狀平裂藻和卵形隱藻均與電導率、透明度、化學需氧量、pH呈顯著負相關性,水華微囊藻與之幾乎相反。按圖8可將占比大于5%的藻類種群大致分為III類,第I類僅與pH、化學需氧量、透明度、電導率相關性較強,與其它環境因子關聯較弱;第II類與第I類幾乎完全相反,第III類又幾乎與第I類相同。由sol文件可知,pH、透明度、電導率和總氮是驅動占比大于5%藻類種群變化的主要環境因子。
3 討 論
3.1 水體環境因子
根據《地表水環境質量標準》(GB 3838—2002),官地水庫水體溶解氧、pH、化學需氧量等指標全年均處于I類水質標準范圍內。冬春季水體總氮含量變幅為0.56~0.94 mg/L,處于III類水質標準范圍內;夏秋季絕大多數斷面介于0.2~0.5 mg/L,且平均值分別為0.35和0.28 mg/L,可視作處于II類水質標準范圍內。由于官地水庫屬于高山峽谷河道型庫區,水體交換量大[16],為日調節水庫[11],庫區有一定流速,可采用一般地表水總磷標準來衡量其水質類別。冬春季水體總磷含量很低,絕大多數斷面低于0.02 mg/L,且兩季平均值為0.020 mg/L,可視作處于I類水質標準范圍內;夏秋季水質總磷變幅為0.37~0.85 mg/L,處于II類水質標準范圍內。總體看來,官地水庫水體溶解氧高、呈弱堿性、營養物質含量低,屬于寡營養水體,水質處于I~III類之間。
官地水庫水溫夏秋季升高,而夏秋季也是雅礱江流域雨季來臨的時節,雨水增多入庫流量擴大帶來的磷元素促使總磷升高、氮磷比下降,但水量猛增也稀釋了原本水體中的氮、鈣、鎂等營養元素,使總氮和硬度迅速降低。夏季由于雨水過多造成洪流,致使該季度水體透明度驟降;秋季上游更清澈的水體持續匯入庫區,促使水體透明度迅速加深。一般水體硬度與電導率呈顯著正相關性[17],但官地庫區水體硬度和電導率變化趨勢完全不同,相關性很弱,這可能是入庫水體硬度和電導率差異較大,致使匯合時兩者“中和”造成硬度與電導率變化不一致。總體看來,官地水庫屬于日調節型,水體交換量大,水質受入庫水體影響較深,呈現較為明顯的季節變化趨勢。
3.2 浮游植物群落多樣性
官地水庫浮游植物群落香農威納指數全年平均值為2.50,水質總體處于β-中污帶,夏季屬于清潔水質;均勻度指數絕大多數大于0.50,全年平均值為0.59,水質總體均勻度良好;辛普森指數絕大多數大于0.50,全年平均值為0.67,表明水體總體良好。研究表明[18-20],水溫升高有助于浮游藻類快速生長和繁殖,影響浮游植物群落豐度的主要環境因子,與本研究中藻類密度與水溫呈極顯著正相關性結論一致。官地水庫常年水溫較低,冬春夏季水溫升高促使部分種類迅速生長,雖致使整體群落密度和種群數量增加,但部分藻類密度比例增長速度高于種群數量增加的效果,從而造成群落香農威納指數、均勻度指數和辛普森指數均與水溫呈負相關性;而且秋季總氮和氮磷比持續降低造成種間競爭激烈[3],使得群落密度和種群數量快速下降,而種群數量降低的效果又相對高于密度,從而形成前文和種群數量與水溫呈顯著負相關性的結果。總體上,水溫為浮游植物群落種群數量和藻類密度的限制因子;調查前期總磷也為限制因子,后期則總氮也為限制因子[21]。
3.3 浮游植物群落優勢類群
官地水庫冬春季以硅藻門、綠藻門和藍藻門藻類為主,夏秋季以硅藻門和綠藻門藻類為主,硅藻為全年主要類群之一預示著水質良好[22];總體上為綠-硅藻水相,與國內大多數水庫、湖泊類似[23-25]。通常大型水庫和湖泊冬春季水溫低,更有利于硅藻生長,而更喜高溫的藍藻則在水溫較高的夏秋季成為優勢類群[26-28]。由于官地水庫夏秋季為雨季,雨水增多,入庫增大,水體流速加快,此時有較硬硅殼、體積和重量均較大的硅藻可像河中泥沙一樣借助一定的縱向流速與流速梯度等產生表面壓力所形成的浮力來懸浮于水中[29],從而占據生長優勢成為優勢類群;而冬春季水體流速低,體積和重量較小且具備氣囊的藍藻占據生長優勢成為優勢類群[13, 30]。所以官地水庫浮游藻類藻相主要隨季節發生變化,而水體理化因子和營養含量則為所適應的藻類種群助力。