粉防己堿誘導雌激素受體陽性乳腺癌細胞自噬與凋亡的機制研究
2021-11-01 02:40:02趙立娜李以良祝東升
解放軍醫藥雜志 2021年10期
關鍵詞:乳腺癌
趙立娜,翟 臻,梁 晨,李以良,李 巍,祝東升
雌激素受體(ER)陽性乳腺癌是最常見的乳腺癌亞型之一[1],其主要治療方法為內分泌治療,但長期治療可能會導致多種不良反應及內分泌抵抗,從而降低了有效性[2]。因此,針對ER陽性乳腺癌的治療方法仍舊面臨重重困難,迫切需要開發新的治療策略。
粉防己堿(tetrandrine,TET)是從粉防己(StephaniatetrandraS.Moore)根部提取的一種雙芐基異喹啉生物堿[3]。多項研究已經證明TET通過誘導細胞凋亡從而發揮抗腫瘤活性[3-4]。內質網是細胞質內參與細胞穩態、凋亡和蛋白質折疊等多種生理過程的細胞器[5],內質網應激則是細胞自噬和(或)凋亡的重要途徑[6]。基于上述理論研究,誘導內質網應激從而促進腫瘤細胞凋亡在抗癌治療中的臨床價值得到了越來越多的關注;但TET是否促進ER陽性乳腺癌細胞凋亡、是否通過內質網應激發揮促凋亡作用尚不清楚。本研究以BT474細胞為實驗模型,揭示內質網應激對TET誘導ER陽性乳腺癌細胞自噬與凋亡的作用。
1 材料與方法
1.1細胞系 ER陽性乳腺癌細胞系BT474購自美國模式培養物研究所(ATCC)。
1.2主要試劑及儀器 TET(Sigma)、RPMI 1640(Gibco)、胎牛血清(FBA,Gibco)、胰酶(Gibco)、MTT(Sigma)、BCA蛋白定量試劑盒(Thermo)、Annexin V和PI凋亡試劑盒(索萊寶)。兔源抗LC3-Ⅱ單克隆抗體、兔源抗Beclin1單克隆抗體、兔源抗GRP78抗體、兔源抗p-PERK抗體、兔源抗p-eIF2α抗體、兔源抗CHOP抗體、兔源抗ATF6抗體、兔源抗β-actin抗體(Cell Signaling Technology,CST)、HRP標記羊抗兔IgG二抗(Santa Cruz Biotechnology)。酶標儀(Bio-Rad)、流式細胞儀(BD)、蛋白電泳系統(Bio-Rad)、凝膠成像系統(Bio-Rad)、細胞培養箱(Thermo)。
1.3細胞培養 乳腺癌細胞系BT474使用RPMI 1640 培養液+10% FBS,于37 ℃,5% CO2培養箱中培養。
1.4MTT檢測 對數期BT474細胞經0.25%胰酶消化后,接種于96孔板中,每孔細胞數3×103個。培養24 h后,加入0(對照組)、5、15、30 μmol/L TET,于5% CO237 ℃培養箱中繼續培養72 h。在培養終止前4 h,每孔沿孔壁加入5 mg/ml MTT溶液10 μl。培養結束后,小心吸棄培養液終止培養,每孔沿孔壁加入150 μl DMSO,輕柔震蕩15 min。將96孔板放入酶標儀,檢測490 nm處各孔吸光度。
1.5細胞凋亡檢測 細胞以1×106個/孔的密度接種于6孔板中,用0、5、15、30 μmol/L TET處理72 h后收獲細胞;PBS充分漂洗2次,沿孔壁加入200 μl緩沖液,輕彈6孔板底部以重懸細胞,分別吸取100 μl細胞懸浮液加入1.5 ml EP管中;根據細胞凋亡檢測試劑盒(Thermo Fisher Scientific)說明書,取10 μl 1∶1混合的Annexin-V 和PI 預混液加入細胞懸浮液中,避光孵育15 min,加入150 μl結合緩沖液,采用FACScanⅡ流式細胞儀進行檢測。計算凋亡細胞比例。
1.6蛋白免疫印跡法檢測蛋白表達 收集0、5、15、30 μmol/L TET處理72 h后的BT474細胞,經RIPA裂解液裂解后,BCA蛋白定量檢測蛋白含量,取40 μg蛋白上樣檢測Beclin1和LC3-Ⅱ蛋白含量。LC3-Ⅱ抗體(1∶2000)、Beclin1抗體(1∶2500)、GRP78抗體(1∶2000)、p-PERK抗體(1∶2500)、p-eIF2α抗體(1∶2500)、CHOP抗體(1∶5000)、ATF6抗體(1∶2000)、β-actin抗體(1∶2500),4 ℃孵育過夜;清洗后加入HRP標記羊抗兔IgG二抗,室溫孵育1 h(1∶5000);清洗后參照BCL顯色液說明書配置顯色液,加入顯色液至膜上;凝膠成像系統掃描成像,目標蛋白灰度值/內參灰度值代表相對蛋白表達量。
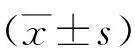
2 結果
2.1TET降低BT474細胞活力 TET 5、15、30 μmol/L組細胞活力分別下降10.5%、31.3%、57.2%。與對照組比較,TET 5、15、30 μmol/L組細胞活力顯著降低,且呈濃度依賴性,差異有統計學意義(P<0.05,P<0.01)。見圖1。
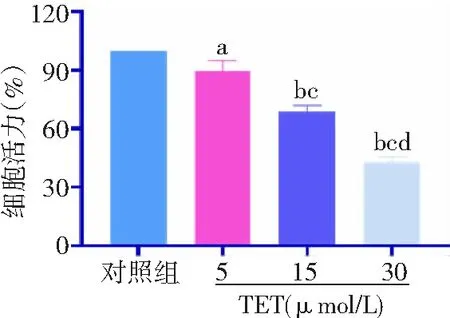
圖1 不同濃度TET處理后BT474細胞活力比較對照組不加TET;TET為粉防己堿;與對照組比較,aP<0.05,bP<0.01;與5 μmol/L組比較,cP<0.01;與15 μmol/L組比較,dP<0.01
2.2TET促進BT474細胞凋亡 細胞凋亡率對照組為(10.35±0.78)%,TET 5、15和30 μmol/L組分別為(16.25±0.62)%、(33.49±0.82)%、(37.01±0.73)%。與對照組比較,TET 5、15和30 μmol/L組細胞凋亡率顯著增高,且呈濃度依賴性,差異有統計學意義(P<0.05,P<0.01)。見圖2。
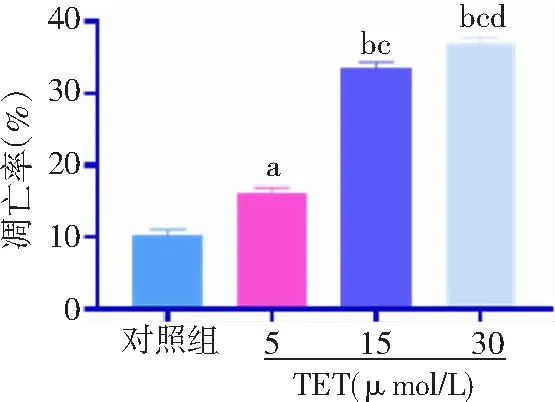
圖2 不同濃度TET處理后BT474細胞凋亡水平比較對照組不加TET;TET為粉防己堿;與對照組比較,aP<0.05,bP<0.01;與5 μmol/L組比較,cP<0.01;與15 μmol/L組比較,dP<0.01
2.3TET促進BT474細胞自噬 與對照組比較,TET 5、15、30 μmol/L組細胞自噬相關蛋白Beclin1和LC3-Ⅱ表達水平顯著升高,且15、30 μmol/L組高于5 μmol/L 組,30 μmol/L組高于15 μmol/L組,差異有統計學意義(P<0.05,P<0.01)。圖3、4。
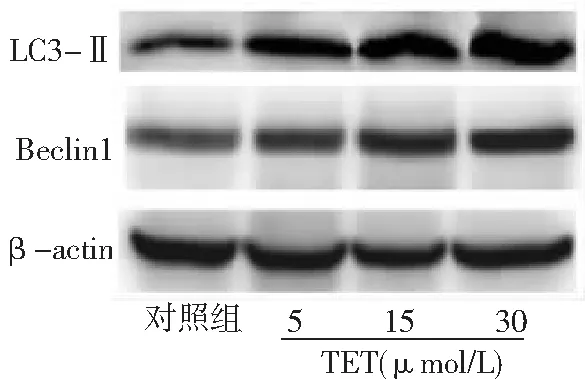
圖3 不同濃度TET處理后BT474細胞自噬相關蛋白Beclin1和LC3-Ⅱ表達蛋白免疫印跡圖對照組不加TET;TET為粉防己堿
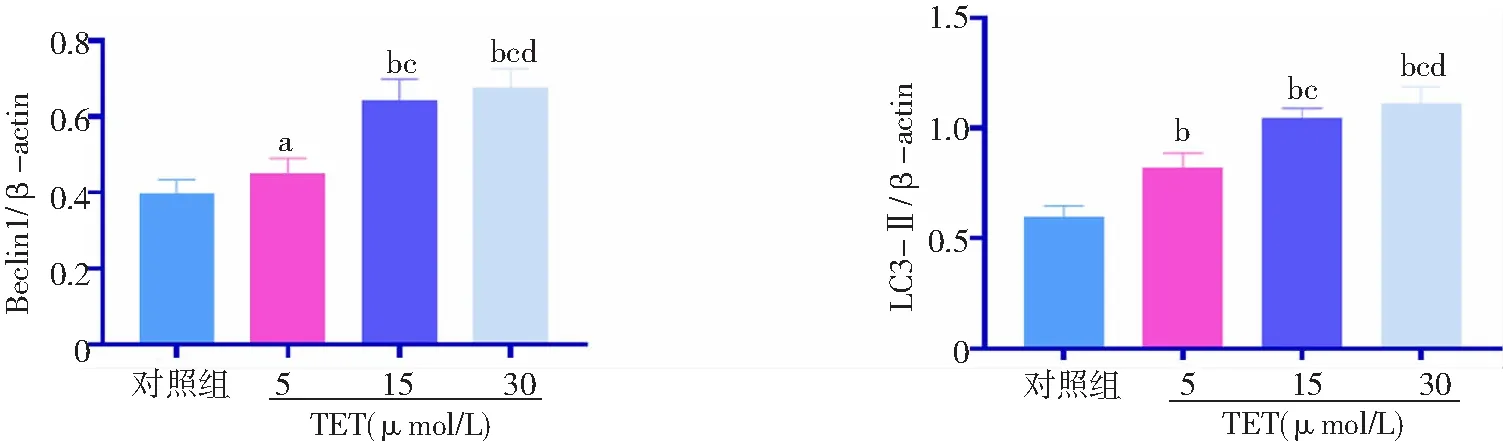
圖4 不同濃度TET處理后BT474細胞自噬相關蛋白Beclin1和LC3-Ⅱ相對表達水平比較對照組不加TET;TET為粉防己堿;與對照組比較,aP<0.05,bP<0.01;與5 μmol/L組比較,cP<0.01;與15 μmol/L組比較,dP<0.01
2.4TET激活BT474細胞內質網應激 與對照組比較,TET 5、15、30 μmol/L組內質網應激通路相關蛋白萄萄糖調節蛋白78(GRP78)、p-PERK、p-eIF2α、CHOP表達水平升高,且15、30 μmol/L組高于5 μmol/L組,30 μmol/L組高于15 μmol/L組,差異有統計學意義(P<0.05,P<0.01)。對照組與TET 5 μmol/L組ATF6表達水平比較差異無統計學意義(P>0.05)。與對照組和5 μmol/L組比較,TET 15、30 μmol/L組ATF6表達水平升高,且30 μmol/L組高于15 μmol/L組(P<0.01)。見圖5、6。
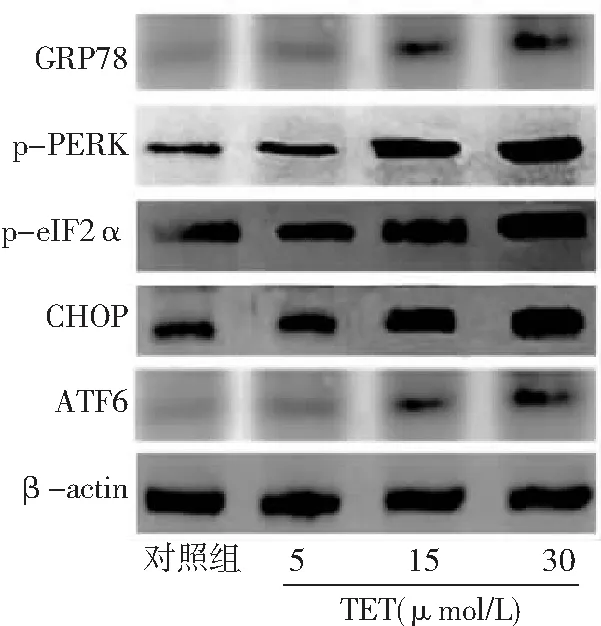
圖5 不同濃度TET處理后BT474細胞內質網應激通路相關蛋白表達免疫印跡圖對照組不加TET;TET為粉防己堿,GRP78為萄萄糖調節蛋白78
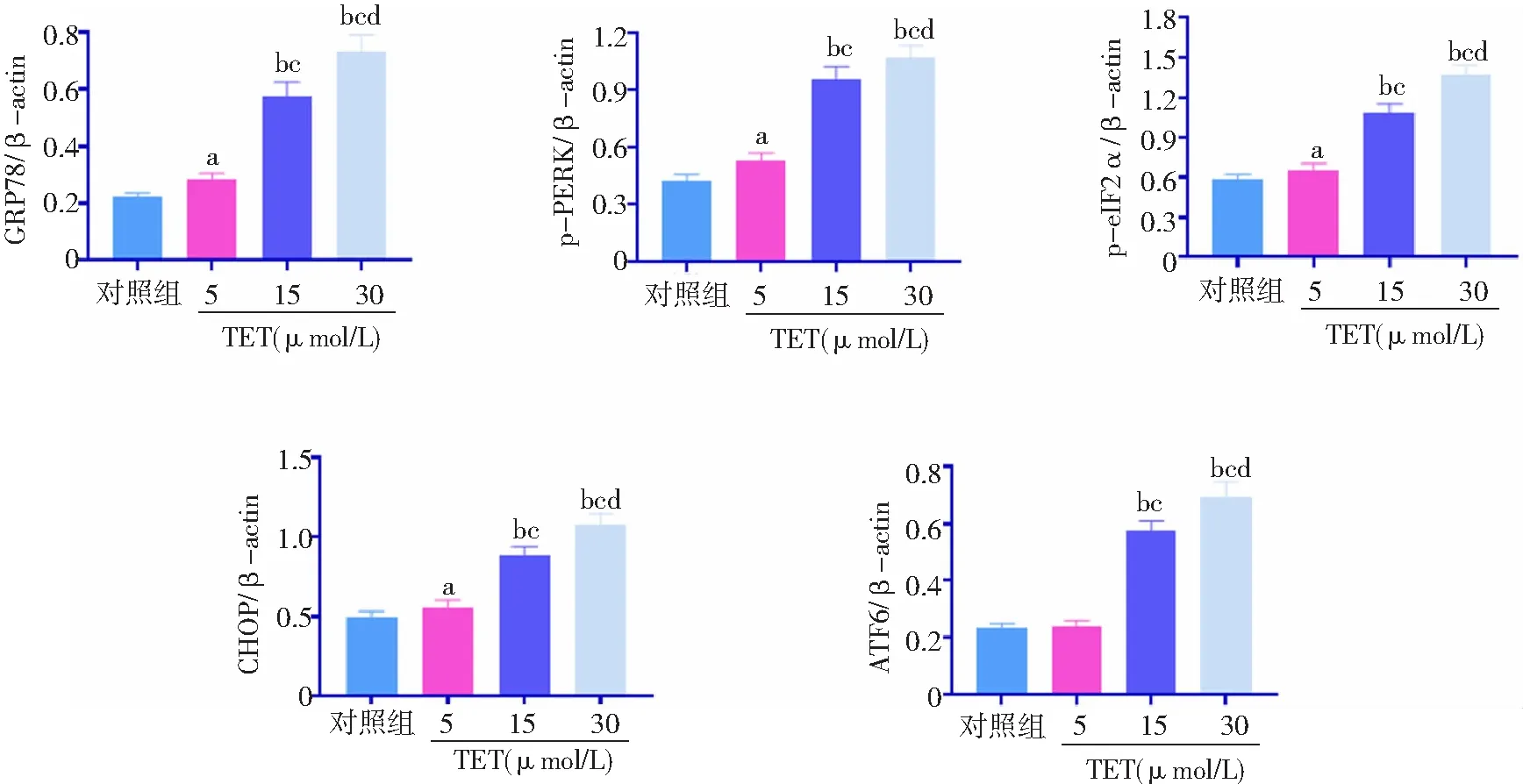
圖6 不同濃度TET處理后BT474細胞內質網應激通路相關蛋白相對表達水平比較對照組不加TET;TET為粉防己堿,GRP78為萄萄糖調節蛋白78;與對照組比較,aP<0.05,bP<0.01;與5 μmol/L組比較,cP<0.01;與15 μmol/L組比較,dP<0.01
3 討論
目前臨床針對ER陽性乳腺癌治療包括芳香化酶抑制劑和抗雌激素化合物,雖然內分泌治療已經大幅降低了乳腺癌的復發率和病死率,但臨床不良反應和獲得性耐藥仍然是該治療方案的最大挑戰[2]。因此,亟須探究和開發新的有效治療手段。
TET對多種受體狀態的乳腺癌細胞有抗增殖和促凋亡作用,增加放化療的敏感性[6-7]。既往研究證實TET抑制PI3K/AKT/mTOR信號通路,從而促進三陰性乳腺癌細胞系MDA-MB-231的自噬和凋亡[3]。但TET是否促進ER陽性乳腺癌細胞自噬和凋亡及其相關作用機制研究較少。
自噬是溶酶體介導的細胞降解過程,該生理過程在腫瘤進展中起重要作用[8],也是癌癥治療的潛在靶點,是增強放療和(或)化療敏感性的策略之一[9]。Beclin1蛋白是自噬體形成的重要因子,介導自噬相關蛋白的亞細胞定位,從而調控自噬泡的形成及隨后成熟[10]。LC3-Ⅱ附著于自噬體膜上,是自噬體的結構蛋白[11]。文獻報道,TET誘導胃癌細胞自噬與凋亡,其作用機制與下調抗凋亡蛋白Bcl-2及促進細胞自噬標志物蛋白Beclin1表達有關[12]。本研究揭示了TET促進ER陽性乳腺癌細胞系BT474的自噬和凋亡,與既往研究一致,TET處理可提高BT474細胞中Beclin1和LC3-Ⅱ表達,證明TET能夠激活自噬信號通路,從而抑制細胞增殖,促進細胞凋亡,且該效應具有明顯的濃度依賴性。
內質網應激可通過激活細胞自噬信號通路導致細胞凋亡[13]。內質網應激是真核生物中重要的應激-防御機制之一,其通過未折疊蛋白反應(UPR)及其他相關分子信號調控自噬與細胞凋亡[13]。UPR信號通路由以下通路組成:PERK通路[14]、IRE1通路[15]和ATF6通路[16]。內質網分子伴侶GRP78是評估內質網應激水平的分子標志物[17]。Tiwari等[18]報道γ-生育三烯酚可提高乳腺癌細胞內質網應激和細胞自噬水平,促進癌細胞凋亡;同樣,法卡地醇可通過內質網應激途徑誘導癌細胞凋亡[19]。本研究發現TET可激活ER陽性乳腺癌細胞內質網應激信號通路相關蛋白GRP78、p-PERK、p-eIF2α、CHOP、ATF6表達,激活經典p-PERK、CHOP和ATF6通路。證實TET可能通過內質網應激激活細胞自噬細胞通路和細胞凋亡,從而抑制ER陽性乳腺癌細胞系BT474的增殖。與既往文獻報道一致。本研究為TET治療ER陽性乳腺癌提供了理論依據,但不足的是本研究使用的細胞系有限,后續可擴充乳腺癌細胞系類型以驗證本結論。
綜上所述,TET通過內質網應激通路激活細胞自噬,促進細胞凋亡,抑制ER陽性乳腺癌細胞BT474增殖。本研究豐富了TET在ER陽性乳腺癌中的作用機制,為臨床的研究和應用奠定了基礎。
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
現代臨床醫學(2022年1期)2022-02-12 02:04:58
甘肅科技(2020年20期)2020-04-13 00:30:42
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
幸福家庭(2019年14期)2019-01-06 09:15:38
祝您健康(2018年5期)2018-05-16 17:10:16
癌癥進展(2016年9期)2016-08-22 11:33:20
中國組織化學與細胞化學雜志(2016年4期)2016-02-27 11:16:08
