倒立水母Cassiopea andromeda的POU、Hox基因家族的多樣性及系統發育分析
2021-11-19 09:49:44陳依婧鄭森林彭勝藍付騰葦
應用海洋學學報 2021年4期
陳依婧, 鄭森林*, 彭勝藍, 林 茂, 付騰葦
(1.自然資源部第三海洋研究所, 福建 廈門 361005; 2.景德鎮陶瓷大學, 江西 景德鎮 333046)
刺胞動物門最早出現在前寒武紀[1-2],作為真后生動物中最原始的類型,刺胞動物門的多樣性是動物界中最古老的進化事件之一[3]。系統發育基因組學分析表明刺胞動物門(包括水母動物亞門和珊瑚動物亞門)是兩側對稱動物的姊妹分支[2, 4-5],水母動物亞門是研究發育生物學和真后生動物早期多細胞演化的理想材料。水母動物亞門由立方水母綱、缽水母綱、水螅蟲綱和十字水母綱組成[6]。目前已經對幾種刺胞動物的全基因組進行了測序,如珊瑚蟲綱的指形鹿角珊瑚(Acroporadigitifera)[7]、星狀海葵(Nematostellavectensis)[8]、水螅蟲綱的水螅(Hydravulgaris)[9]、半球美螅水母(Clytiahemisphaerica)[6]和缽水母綱的海月水母(Aureliaaurita)[10]。目前NCBI數據庫中水母亞門僅有6屬9種全基因組數據,其中缽水母綱僅有3屬全基因組數據,可見相關研究尚較匱乏。
絕大多數水母的生命周期都包含營固著生活的水螅體形態,它經過無性繁殖產生浮游形態的水母體[11-12],但是有一類水母在形成水母體之后仍然保持部分的附著及倒立的行為,這就是俗稱的“倒立水母”。安朵仙水母(Cassiopeaandromeda)是一種最為常見的倒立水母,常見于熱帶及亞熱帶淺海沿岸生態系統,如紅樹林、砂質泥灘和海草床[13],最常在紅樹林樹葉覆蓋的泥質基質上定居,因而也被稱為“紅樹林水母”。仙女水母屬(Cassiopea)隸屬于缽水母綱,與其他浮游缽水母行為顯著不同,其生命周期大部分為呈靜止狀態的水母體,形態特異的中凹扁平傘部附著于基質上,而其凸出的觸手和口唇朝上[14]。仙女水母這種特殊的倒立附著生物學行為,理論上應有其遺傳學基礎。但這種倒立水母的全基因組尚無數據,本研究對典型的倒立水母——安朵仙水母進行了全基因組測序,并同步測序了同屬缽水母綱的巴布亞硝水母(Mastigiaspapua)、海蜇(Rhopilemaesculentum)用于平行比較。
目前關于仙女水母特殊倒立行為相關基因的研究只見零星報道。仙女水母的特殊行為包括:倒立、附著、活動能力較低。生物倒立行為可能與感知重力的結構有關,刺胞動物門中僅有缽水母和立方水母的傘緣有感覺器(Rhopalia),其細微結構眼點(Ocelli)感知光,平衡石(Statoliths)感知重力,還能控制游泳肌肉收縮的速度,是水母網狀神經系統的組成部分[15]。已有研究認為水母感覺器是其祖先具有感覺功能的結構通過不同感覺細胞群中POU編碼基因的差異表達而演變出來的[16]。
仙女水母的“睡眠”類似行為值得關注,作為刺胞動物的代表,它是動物界能被觀察到該生物行為的最早的一支進化分支[17]。其“睡眠”特征為可快速逆轉的靜止狀態,及其在“睡眠”期間對感官刺激的延遲響應[13, 17]。因此,本研究對水母神經、感官發育的相關基因進行了分析,包括決定胚胎前后軸分化的Hox蛋白[18]和調節胚胎神經系統發育的POU蛋白[19]。
在附著行為方面,常見的水生生物粘附蛋白有神經肽和磷酸化蛋白等,但目前關于水母粘附的分子機制的研究信息很少[20],本研究著重比較分析安朵仙水母、巴布亞硝水母、海蜇與營附著生活的水螅的粘附相關蛋白的種類和數目差異。
1 材料與方法
1.1 樣本采集和基因組測序
用于基因組分析的安朵仙水母、海蜇、巴布亞硝水母的樣本由實驗室培養獲得,其母代采集地分別為中國海南海、中國海渤海和熱帶西太平洋島嶼。取水母的傘部組織在CTAB緩沖液中用20 mg/L的蛋白酶K(Sigma公司)消化過夜,然后使用酚氯仿試劑(Solarbio公司)提取基因組DNA,獲得的DNA樣在-20 ℃下保存。采用PE150建庫方案及HiSeq×10測序平臺,預估測序深度為水母基因組(參照海月水母基因組)的200×。
1.2 全基因組拼接和注釋
使用MEGAHIT軟件[21]將高質量的濾后數據組裝到不同的重疊群(Contig)。為了對這3種水母進行基于同源性的基因預測,我們使用TblastN(閾值1.0E-5)檢索了同源物種水螅的蛋白序列[22]。利用GenBlast[23]的程序GenBlastA與GenBlastG對外顯子區進行檢索及注釋。
1.3 同源基因比對和系統發育分析
將安朵仙水母、海蜇、巴布亞硝水母的所有注釋蛋白序列與刺胞動物門水螅、指形鹿角珊瑚、星狀海葵及海月水母的所有POU、Hox/ParaHox基因做BLASTP[22]最佳匹配(閾值1.0E-5),另外利用Orthofinder-V2.2.7[24]進行同源基因家族聚類,得到預測的POU、Hox/ParaHox基因序列集。使用MUSCLE[25]對POU、Hox/ParaHox基因進行多重序列比對。使用ModelGenerator[26]選擇最佳替代模型LG+G后,使用PhyML-V3.0[27]構建POU基因的最大似然法(Maximum Likelihood,ML)系統發育樹,自展值(Bootstrap)=1 000。使用FastTree-V2.1.11[28]、LG+CAT模型構建Hox/ParaHox基因的ML系統發育樹。輸出結果應用BioEdit[29]繪蛋白序列比對圖、Evolview V3[30]美化系統發育樹。
2 結果與討論
2.1 全基因組測序及注釋
我們測序并組裝了3種缽水母安朵仙水母、巴布亞硝水母和海蜇的首個全基因組(表1)。巴布亞硝水母樣本的原始測序數據去除雜質后得到78.78 G的數據,組裝后最終得到506 Mb的基因組;拼接獲得的重疊群一共506 243 595 bp,總共372 755條重疊群,N50值(將重疊群按長度從大到小排列累加長度,當總長達到總堿基數50%時的重疊群長度)為2 748 bp。重疊群最小長度為200 bp, 最大為68 090 bp, 平均1 358 bp。安朵仙水母樣本的原始測序數據去除雜質后得到67.05 G的數據,組裝后最終得到423 Mb的基因組;拼接獲得的重疊群一共423 279 109 bp,總共205 460條重疊群,N50 值為4 888 bp。重疊群最小長度為200 bp, 最大為76 879 bp, 平均2 060 bp。海蜇樣本的原始測序數據去除雜質后得到65.83 G的數據,組裝后最終得到384 Mb的基因組;拼接獲得的重疊群一共384 147 202 bp,總共337 206條重疊群,N50 值為2 348 bp。重疊群最小長度為200 bp, 最大212 791 bp, 平均1 139 bp。通過對全基因組應用同源基因預測方法,我們預測了39 815個巴布亞硝水母的蛋白質編碼基因,63 191個安朵仙水母的蛋白質編碼基因,66 907個海蜇的蛋白質編碼基因。可見3種缽水母中,海蜇基因組最小但非冗余蛋白編碼基因最多。
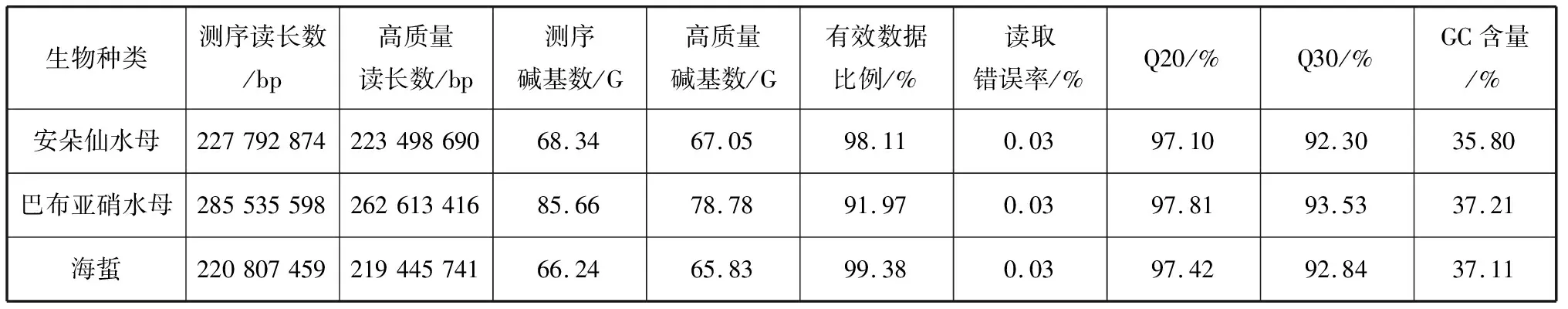
表1 測序產出數據質量
2.2 Hox蛋白序列的多樣性與系統發育分析
分別檢索到安朵仙水母的11個、海蜇的11個和巴布亞硝水母的9個Hox同源異型盒(Homeobox)蛋白序列(不計重復序列)。系統發育分析顯示這些序列與同屬缽水母綱的海月水母的親緣關系較近,且刺胞動物的Hox基因的相同亞型聚類在一起(圖1、2)。從類型上劃分,安朵仙水母的其中6個、海蜇的其中6個及巴布亞硝水母的其中4個屬于“后段Hox基因”即HoxE,HoxF(PosteriorHoxGene),這3種缽水母“后段Hox基因”完整。與其他刺胞動物門一樣,安朵仙水母、海蜇和巴布亞硝水母不存在“中段Hox基因”(CentralHoxGene)。安朵仙水母、海蜇和巴布亞硝水母的“前段Hox基因”(AnteriorHoxGene)僅有HoxA,但是HoxB,C,D缺失,這與海月水母一致。關于Hox基因的旁系同源(Paralogue)ParaHox基因,安朵仙水母、海蜇和海月水母都含有完整的GSX及XLOX/CDX基因,但是巴布亞硝水母和水螅、指形鹿角珊瑚一樣只含有GSX基因。
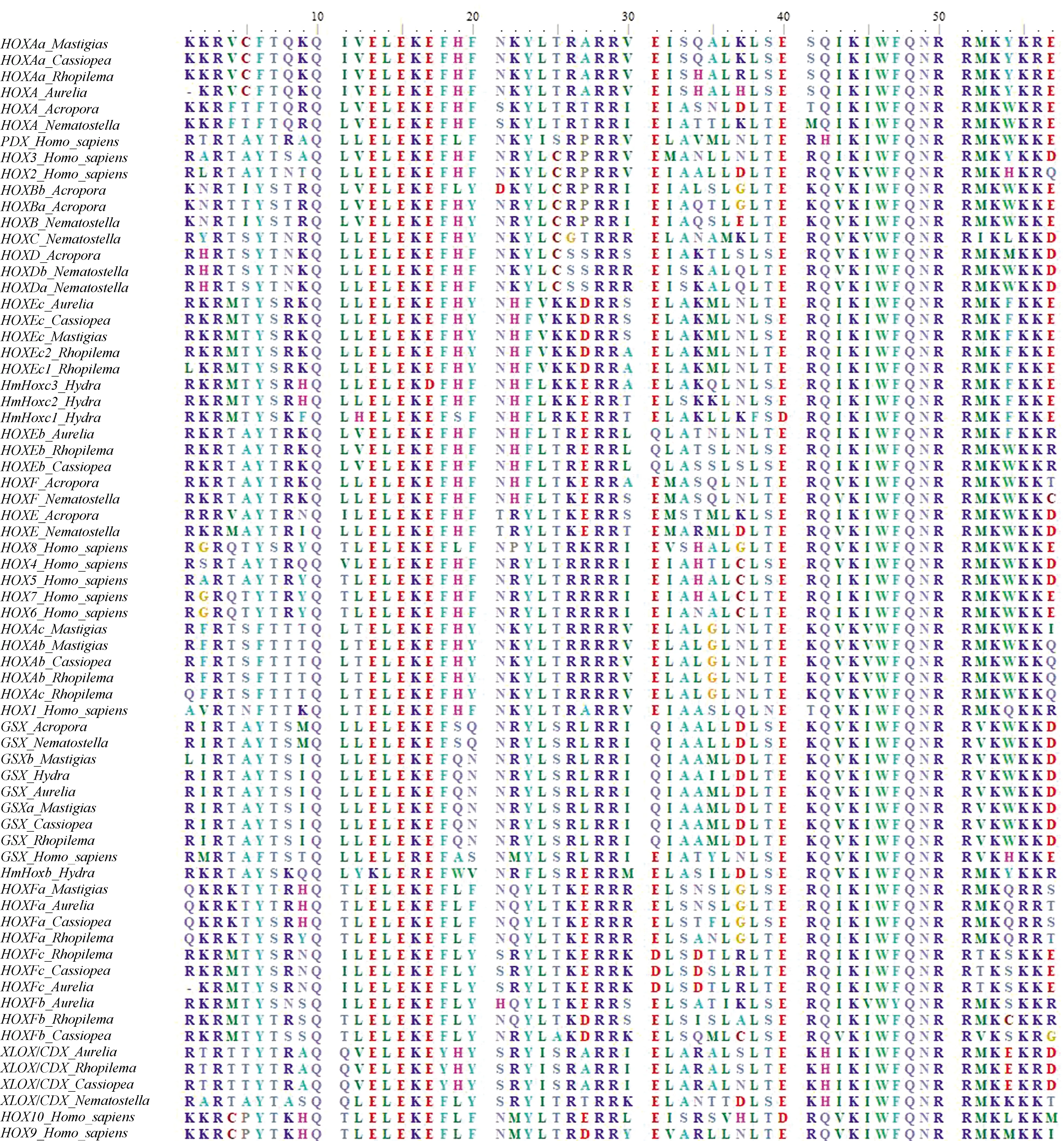
圖1 刺胞動物門及人類的Hox/ParaHox基因家族蛋白序列比對
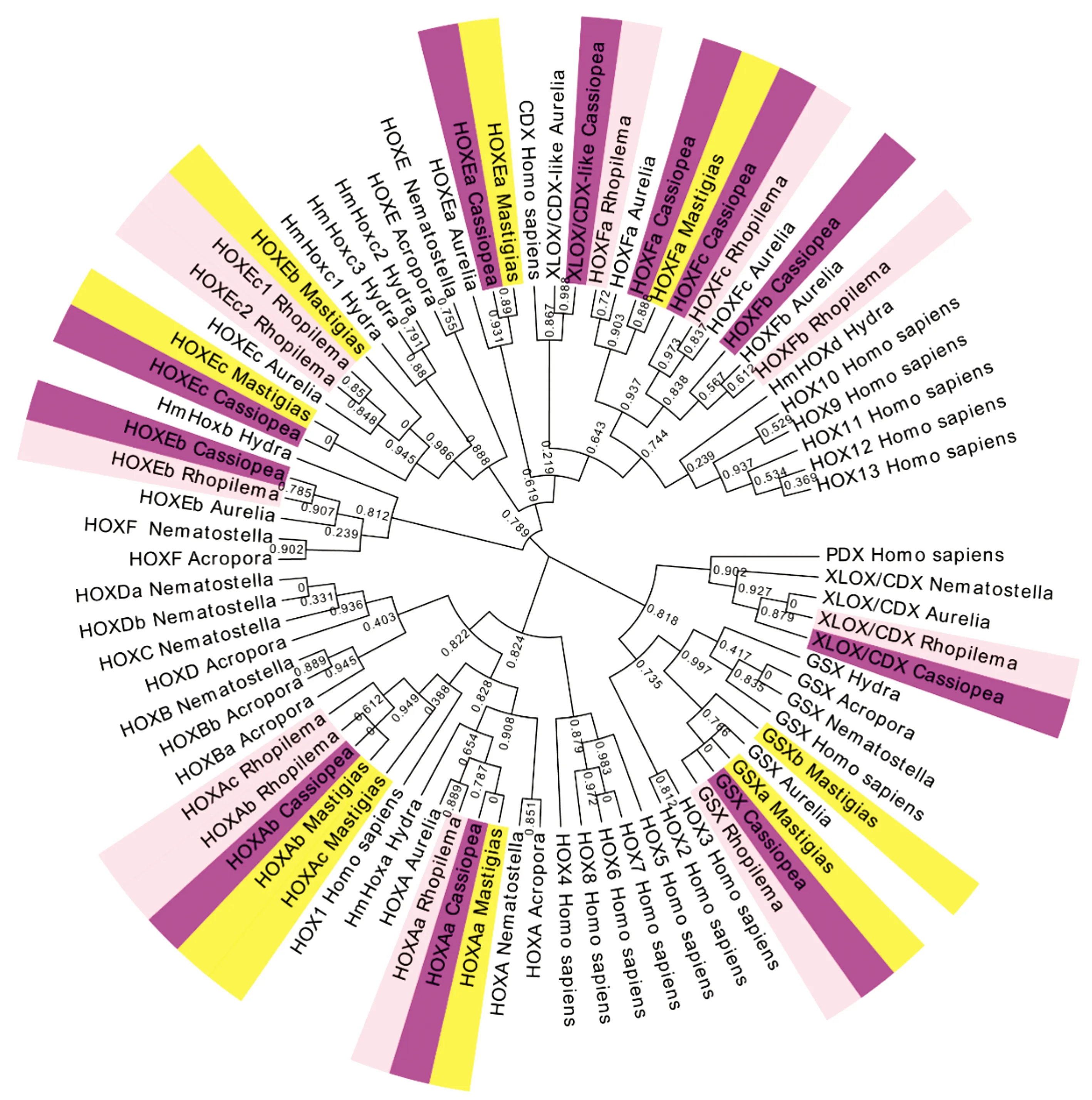
圖2 刺胞動物門及人類的Hox/ParaHox基因家族蛋白序列的ML系統發育樹
2.3 POU蛋白序列的多樣性與系統發育分析
Hox和POU基因家族都具有高度保守的結構域,而缽水母綱POU的類型和數目具有更大的種間差異性(圖3、4)。在根口水母目中,安朵仙水母和海蜇各有4種POU亞型,而巴布亞硝水母僅有2種。安朵仙水母和海蜇有POU-1,-2/3/5,-4,-6基因;巴布亞硝水母僅有POU-3,-6基因;旗口水母目的海月水母則有2種亞型POU-1和POU-4,缺少POU-2/3/5和POU-6亞型。可見POU蛋白有兩個結合域,位于N端的POU特異結構域(POU Specific Domain,1—73 位)與位于C端的POU同源異形域(POU Homeodomain,102—158 位)[31-32],各亞型的特異性氨基酸位點如111、131、158等處見圖3。系統發育分析顯示,刺胞動物POU蛋白的相同亞型分別聚類在一起;各亞型中,3種缽水母的POU蛋白序列與同屬缽水母綱的海月水母的親緣關系較近,與珊瑚蟲綱親緣關系較遠。在進化關系方面,POU-1,6是最原始的POU家族成員。

圖3 刺胞動物門及人類的POU基因家族蛋白序列比對

圖4 刺胞動物門POU基因家族蛋白序列的ML系統發育樹
2.4 與附著機制相關的粘附蛋白的比較
通過比較3種水母基因組注釋信息和已報道的水螅粘附蛋白[20],我們發現α-l-阿拉伯呋喃糖苷酶B(Alpha-l-arabinofuranosidase B,即AbfB)和Galaxin-like蛋白是安朵仙水母、巴布亞硝水母、海蜇共有的潛在粘附蛋白。安朵仙水母具有巴布亞硝水母、海蜇所不具有的獨特潛在粘附蛋白:鐵螯合物還原酶類似蛋白(Ferric-chelate Reductase)和具有半乳糖結合凝集素結構域的鼠李糖結合凝集素類似蛋白(Rhamnose-binding Lectin-like Protein with Galactose binding Lectin Domain)。安朵仙水母的粘附相關谷氨酰胺合成酶的編碼基因拷貝數比巴布亞硝水母多出8個。但我們沒有在這3種水母中發現水螅粘附蛋白中的其他蛋白質家族(包括過氧化物酶、FRG1家族類似蛋白、半胱氨酸蛋白酶抑制蛋白結構域、甲殼素結合環磷酰胺A蛋白結構域和多表皮生長因子類似蛋白11結構域)。
2.5 討論
安朵仙水母具有全面的POU基因亞型,由于POU是神經系統和感覺器官發育的重要調節因子,這很可能是該水母具有倒立附著行為特征的重要遺傳因素之一。在后生動物中,POU一共有6種亞型,其中2/3/5亞型從進化上聚為一簇,可歸為一個類型[33],即共4個亞型。本研究發現安朵仙水母、星狀海葵、海蜇具備所有這4種亞型基因,指形鹿角珊瑚有3種亞型,而水螅和缽水母綱的海月水母、巴布亞硝水母僅有2種亞型。在本研究分析的刺胞動物中,安朵仙水母的POU-1,-6亞型的核苷酸多態性最高,可能與其特殊的倒立附著行為有關。其中POU-2/3/5亞型在星狀海葵、指形鹿角珊瑚、安朵仙水母中序列變異較多,分別有3、2、2種不同序列,但在海蜇、巴布亞硝水母中僅有1種序列,且在水螅及海月水母中缺失。根據以上所述的種類和數目情況,營附著生活的安朵仙水母的POU基因家族可能與營固著生活的珊瑚、海葵更接近。其中安朵仙水母所具有的POU-3亞型可能是個潛在的擴增基因,它在兩側對稱動物中未被檢索到[10]。POU表達具有組織特異性,比如在索氏桃花水母(Craspedacustasowerbyi)中,POU-6多數在平衡囊(Statocyst)中表達,較少在生殖腺(Gonad)和胃腔(Gastric cavity)中表達;POU-4中csPOU-4f1在平衡囊中表達,csPOU-4f2在生殖腺和平衡囊中表達,僅csPOU-4f3未被觀察到在平衡囊中表達;POU-4,-6都幾乎未被觀察到在卵母細胞組織(Oocyte)中表達[32],安朵仙水母POU-4與其中csPOU-4f3序列相似性最高。已知POU-4參與控制視覺器官的發育,如調控脊椎動物中視網膜神經節細胞的發育[34],但是該蛋白在水母眼點處的表達情況仍未知。已有研究認為POU-1和POU-4指引了水母不同感覺細胞群類群的發育,如在海月水母中POU-4被認為參與位于頂端細胞表面、具有微絨毛的機械感覺細胞發育過程,而POU-1參與位于感覺器的外細胞層的感覺細胞發育過程[16]。可見POU基因的多樣性對水母的感覺器官功能有重要影響,很可能與安朵仙水母“睡眠”態中對感官刺激延遲響應的特征有關,這反映了盡管簡單生物水母未發育出發達的眼結構,但其相對復雜的感知系統已具備深入研究的價值。由于安朵仙水母具有完整的POU亞型,這些基因可能起到不同功能,共同參與了其倒立、睡眠態行為的復雜神經調節過程。但目前對POU的研究主要集中在高等動物,如POU促進哺乳動物傷口愈合、細胞凋亡、神經誘導、神經元定位等[19, 35],在低等的刺胞動物中的功能尚不明確,其與倒立水母的具體功能關系需進一步研究。
Hox基因家族是決定兩側對稱動物的胚胎前后軸的分化及其他發育基因的重要調節因子,它在非兩側對稱動物中僅存在于刺胞動物門中,所以在刺胞動物門中的研究對Hox基因的早期演化有重要意義[36]。Hox蛋白在刺胞動物門中表現出高度相似的結構域,缽水母屬中不同物種的相同Hox亞型都聚類在一起,都說明其在進化上具有一定的保守性。本研究從全基因組角度發現,在安朵仙水母、海蜇和巴布亞硝水母中未找到“中段Hox基因”,這與先前的研究認為“中段Hox基因”僅在兩側對稱動物中出現、在刺胞動物不存在“中段Hox基因”假說是一致的[36]。本研究發現的另一個現象為安朵仙水母、巴布亞硝水母、海蜇的“前段Hox基因”僅有HoxA(這與海月水母一致),而珊瑚蟲綱的星狀海葵則同時存在Hox-A,B,C,D所有“前段Hox基因”亞型,指形鹿角珊瑚也具有Hox-B,C,D(圖1),這一現象說明缽水母及水螅在進化過程中“前段Hox基因”產生了部分缺失。先前研究認為星狀海葵只具有一支“前段Hox基因”[37-38],但本研究發現其同樣具有“后段Hox基因”HoxE,F。近年以星狀海葵作為模式生物深入研究了Hox基因在刺胞動物的胚胎發育過程中的作用機理[39],但是對于水母中Hox基因的具體功能還了解甚少[40]。
粘附相關蛋白的編碼基因在安朵仙水母和巴布亞硝水母、海蜇間具有明顯差異,推測是安朵仙水母倒立行為的另一個特征因素。水螅基盤細胞分泌的粘接劑的主要成分是蛋白質和聚糖[41],多糖的大量表達是滿足可逆粘附(Reversible Adhesion)的一個條件,但聚糖在粘附蛋白中的根本作用機制仍有待明確[20, 41]。谷氨酰胺合成酶(Glutamine Synthetase)是一種粘附關鍵酶,它控制細胞內氮的使用,從而生成富含氮的蛋白質[42],安朵仙水母的該酶編碼基因拷貝數比巴布亞硝水母多出8個,說明其表達量可能較多。鑒于已知AbfB具有催化O-糖基化合物進行水解反應的功能,推測安朵仙水母和巴布亞硝水母、海蜇的主要粘附相關蛋白可能是O-糖基化的。本研究發現安朵仙水母具有巴布亞硝水母、海蜇所不具備的粘附相關的半乳糖結合凝集素蛋白,而凝集素結構域可為分泌到體外的粘附物質提供額外的比如具有抗細菌降解功能的物質[43],許多附著物種粘附蛋白具有半乳糖結構域,如海膽Nec-1、Nec-2蛋白[44]、海星Sfp1蛋白[45]。鐵螯合物還原酶類似蛋白為安朵仙水母特有的,但巴布亞硝水母、海蜇并不具備,推測這種抗氧化活性物質對水母粘附起作用,可能替代了水螅中過氧化物酶催化粘附蛋白聚合[20, 41]的作用。
盡管水母的細胞類型和數目相對高等動物較少,但仍然呈現了一個復雜的調節網絡:倒立水母“睡眠”態行為可能與網狀神經系統調節運動和感覺器官功能有關,倒立水母附著行為除了受控于基盤處肌肉運動的物理因素[41],還可能與凝集素、蛋白酶、多糖等多種分泌因子相關。
3 結論
通過比較安朵仙水母、海蜇、巴布亞硝水母及其他刺胞動物的重要轉錄調節因子多樣性與系統發育關系發現,安朵仙水母具有更多POU編碼基因和復雜POU亞型,以及具有粘附相關凝集素及還原酶的編碼基因,可能是與安朵仙水母倒立附著生活方式相關的關鍵遺傳信息。首先,安朵仙水母具有全面的POU亞型,其POU亞型的核苷酸多態性較高,非冗余蛋白序列最多。安朵仙水母的POU基因家族的種類數目情況與營固著生活的珊瑚、海葵更接近。其次,安朵仙水母具有巴布亞硝水母、海蜇所不具有的粘附相關鼠李糖結合凝集素和一類抗氧化活性物質鐵螯合物還原酶。3種缽水母共有ABfB和Galaxin-like潛在粘附蛋白。另外,本研究發現缽水母“前段Hox基因”存在部分亞型缺失;安朵仙水母和海蜇的Hox旁系同源ParaHox基因種類完整;研究結果支持刺胞動物不存在“中段Hox基因”的假說。
