煙草白粉病抗性連鎖分子標(biāo)記開(kāi)發(fā)及育種利用
2021-11-22 09:17:40張孝廉張吉順賈蒙驁張莉莉曹領(lǐng)改
中國(guó)煙草學(xué)報(bào) 2021年5期
關(guān)鍵詞:煙草
張孝廉,張吉順*,賈蒙驁,張莉莉,曹領(lǐng)改
1 貴州省煙草科學(xué)研究院,煙草行業(yè)分子遺傳重點(diǎn)實(shí)驗(yàn)室,貴陽(yáng)市觀山湖區(qū)龍灘壩路29號(hào) 550081;
2 貴州大學(xué)生命科學(xué)學(xué)院,貴州省農(nóng)業(yè)生物工程重點(diǎn)實(shí)驗(yàn)室,山地植物資源保護(hù)與種質(zhì)創(chuàng)新省部共建教育部重點(diǎn)實(shí)驗(yàn)室,貴陽(yáng) 550025
煙草白粉病俗稱“上灰”、“下霜”、“上硝”,感病煙葉烘烤后薄如紙,色暗銹褐色,喪失經(jīng)濟(jì)價(jià)值,散葉堆放和鮮煙裝炕階段白粉病菌均可侵染煙葉[1]。目前,煙草白粉病的防治主要以化學(xué)防治為主,極易造成農(nóng)藥殘留,是當(dāng)前煙葉農(nóng)殘超標(biāo)的主要因素之一。培育抗病品種是防治煙草白粉病最經(jīng)濟(jì)、安全和有效的方法,因此,白粉病抗性相關(guān)研究得到了廣泛的關(guān)注,并取得了重要進(jìn)展。小麥中已經(jīng)發(fā)現(xiàn)了50 多個(gè)Pm(Powdery mildew)抗性位點(diǎn)[2-5],擬南芥中也分離到許多涉及不同途徑的白粉病抗性基因[6-8]。其中,mlo(Mildew resistance locus O)基因最先在大麥中[9]被鑒定,由于mlo介導(dǎo)的白粉病抗性具有廣譜、持久的優(yōu)點(diǎn),在小麥等作物中開(kāi)展了廣泛的研究和利用,并取得顯著效果。在番茄[10],辣椒[11],煙草[12]等茄科植物中也陸續(xù)報(bào)道了與白粉病抗性相關(guān)的MLO同源基因。通過(guò)對(duì)茄科植物中白粉病相關(guān)MLO基因鑒定和功能驗(yàn)證,明確了煙草NtMLO1基因可以互補(bǔ)番茄mlo突變體的感病性[12]。NtMLO1和NtMLO2基因突變導(dǎo)致了日本晾曬煙品種Kokubu 的白粉病抗性[13-14]。
煙草白粉病抗病育種中最廣泛而成功利用的是Kokubu 抗性,而國(guó)內(nèi)品種抗煙草白粉病育種也取得了一定的進(jìn)展,孟坤等[15]用不同的白粉病菌對(duì)21 個(gè)煙草品種進(jìn)行接種鑒定,發(fā)現(xiàn)對(duì)兩個(gè)菌種均表現(xiàn)免疫的是塘蓬,表現(xiàn)高抗的為NC89,高感品種為中煙90 和CF203。李華麗等[16]以臺(tái)煙7 號(hào)和白肋21 作為雜交親本,后代自交衍生的127 個(gè)F2 和F2:3 家系為材料,檢測(cè)到1 個(gè)煙草白粉病加性QTL,貢獻(xiàn)率為11.63%。牟建英等[17]利用2317 對(duì)SSR 引物對(duì)煙草白粉病抗性基因進(jìn)行QTL 定位,檢測(cè)到1 個(gè)白粉病抗性位點(diǎn),與SSR 引物52725 緊密連鎖,遺傳距離為1.01cM,其加性效應(yīng)為-14.38,貢獻(xiàn)率為21.02%。
本文通過(guò)對(duì)抗感分離群體進(jìn)行白粉病抗病鑒定,開(kāi)發(fā)與白粉病抗性緊密連鎖的分子標(biāo)記,旨在為國(guó)內(nèi)煙草白粉病抗源的高效利用及抗病育種提供參考。
1 材料與方法
1.1 材料種植和接種鑒定
本研究中使用的煙草材料見(jiàn)表1,由貴州省煙草科學(xué)研究院種質(zhì)資源庫(kù)保存。統(tǒng)一采用漂浮育苗法育苗,置于光照培養(yǎng)箱中,設(shè)置8 h/16 h 光照/黑暗,25℃/23℃,80%濕度。于6 ~8 葉期進(jìn)行白粉病接種實(shí)驗(yàn),取新鮮的感白粉病煙草葉片,將葉片上新鮮的白粉病孢子均勻抖落在待接種煙苗上,每個(gè)材料接種煙苗30 株,置于黑暗中暗培養(yǎng)2 d 后恢復(fù)8 h/16 h光照/黑暗光周期,25℃/23℃,80%濕度,接種后10 d 調(diào)查發(fā)病情況。按照國(guó)家標(biāo)準(zhǔn)《GB/T 23222—2008 煙草病蟲(chóng)害分級(jí)及調(diào)查方法》中對(duì)白粉病發(fā)病等級(jí)的劃分,鑒定煙草材料對(duì)白粉病的抗性。
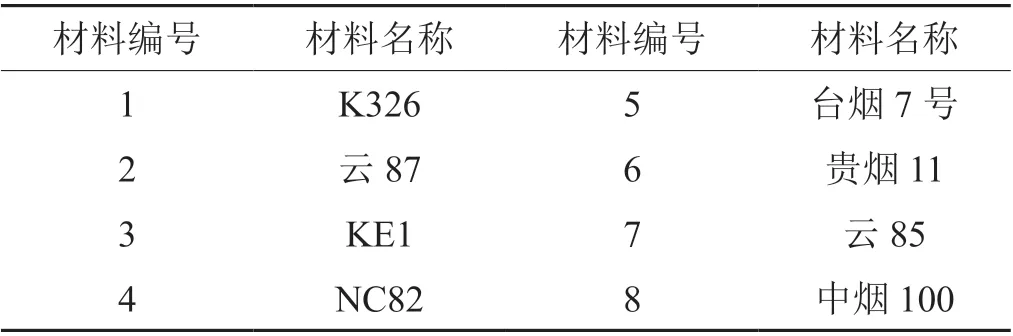
表1 供試煙草品種Tab. 1 Tobacco germplasm resource used in this research
1.2 DNA 提取與NtMLO 基因突變區(qū)的克隆
根據(jù)已報(bào)道NtMLO1和NtMLO2的基因序列[14]設(shè)計(jì)引物MLO1-F/MLO1-R,MLO2-F/MLO2-R(表2)擴(kuò)增突變區(qū)序列。以表1 中材料的DNA 為模板進(jìn)行PCR 擴(kuò)增,擴(kuò)增產(chǎn)物進(jìn)行膠回收送到上海生工公司進(jìn)行基因測(cè)序。
1.3 分子標(biāo)記設(shè)計(jì)與篩選
根據(jù)抗病和感病材料間NtMLO1和NtMLO2的基因序列差異,在差異位點(diǎn)設(shè)計(jì)引物(表2)。PCR 反應(yīng)體系(20 μL):50 ng/μL DNA 模板1 μL,2×Go Taq Mix 10 μL,100 μmol/L 正 反 向 引 物 各1 μL,ddH2O 7 μL。PCR 反應(yīng)程序:94 ℃預(yù)變性3 min,94℃變性30 s,58℃退火30 s,72℃延伸30 s,共30個(gè)循環(huán),72℃延伸10 min,4℃保存。擴(kuò)增產(chǎn)物使用2%的瓊脂糖凝膠進(jìn)行電泳檢測(cè)。

表2 引物序列信息Tab. 2 The sequence information of primers
2 結(jié)果分析
2.1 不同資源的抗性鑒定
選取8 份材料進(jìn)行抗性鑒定,按照國(guó)標(biāo)進(jìn)行調(diào)查和統(tǒng)計(jì),發(fā)現(xiàn)臺(tái)煙7 號(hào)和KE1 表現(xiàn)出高抗白粉病,抗病等級(jí)為1 級(jí),NC82 等材料表現(xiàn)為高感白粉病。
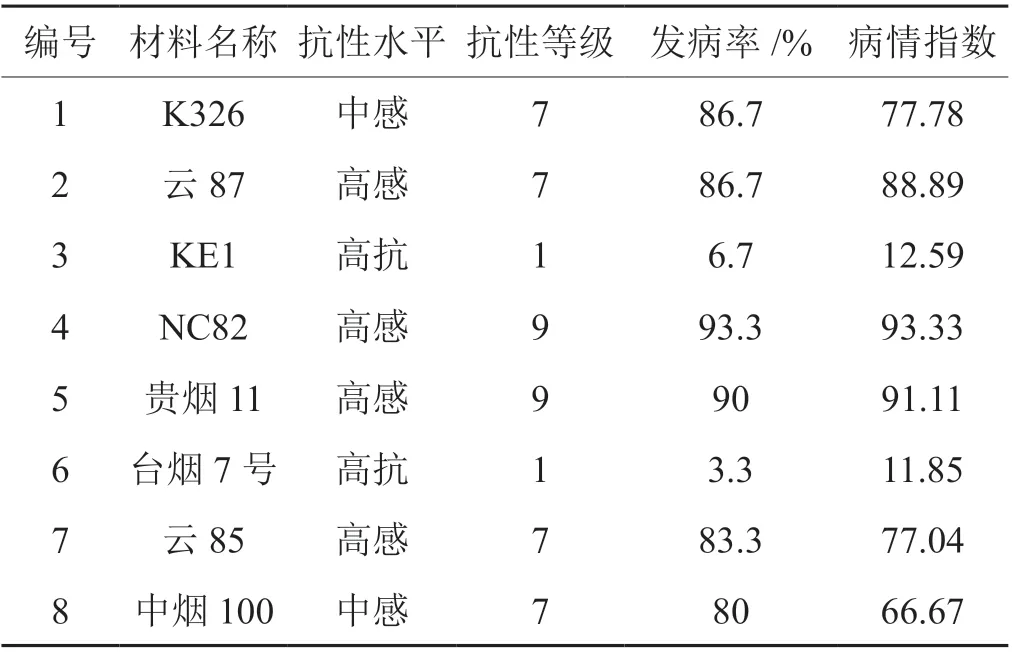
表3 不同煙草品種對(duì)白粉病抗性統(tǒng)計(jì)Tab. 3 The powdery mildew resistance of different tobacco varieties

圖1 不同煙草品種對(duì)白粉病抗性的鑒定Fig. 1 Identification of powdery mildew resistance for different tobacco varieties
2.2 分離群體抗性鑒定及遺傳分析
對(duì)抗病材料臺(tái)煙7 號(hào)和感病材料K326 構(gòu)建的960 個(gè)F2 單株及642 個(gè)BC1F1 單株開(kāi)展了苗期白粉病抗性鑒定,結(jié)果表明(表4)F1 全部表現(xiàn)為感病,F(xiàn)2 群體的感抗分離比符合15:1,BC1F1 群體的抗感分離比為1:3,表明臺(tái)煙7 號(hào)的抗性符合雙基因隱性遺傳規(guī)律。
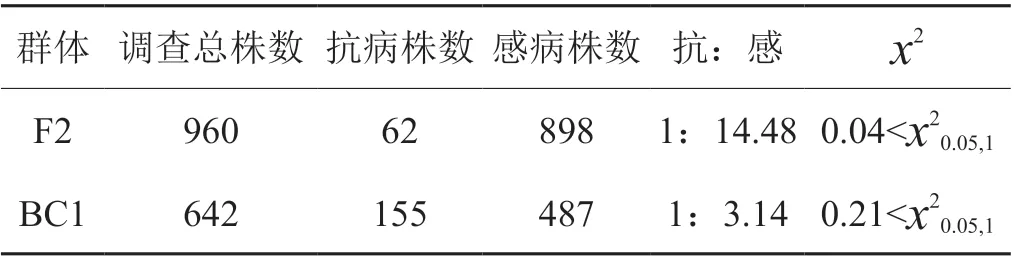
表4 分離群體抗病/感病表型調(diào)查T(mén)ab. 4 Investigation of resistacne/susceptibility phenotype in the segregation population
2.3 不同抗性材料中NtMLO 基因克隆及標(biāo)記開(kāi)發(fā)
以8 份供試材料的DNA 為模板進(jìn)行PCR 擴(kuò)增,結(jié)果在不同材料中都擴(kuò)增到了預(yù)期大小的片段,測(cè)序結(jié)果如圖2 所示,感病材料中NtMLO1和NtMLO2基因的序列與參考序列相同,而臺(tái)煙7 號(hào)、KE1 中的NtMLO1基因序列在2962 ~2963 bp 存在2 個(gè)堿基的差異,從AG 突變?yōu)門(mén)A,NtMLO2基因序列缺失了第2969 ~2970 位的2 個(gè)堿基。
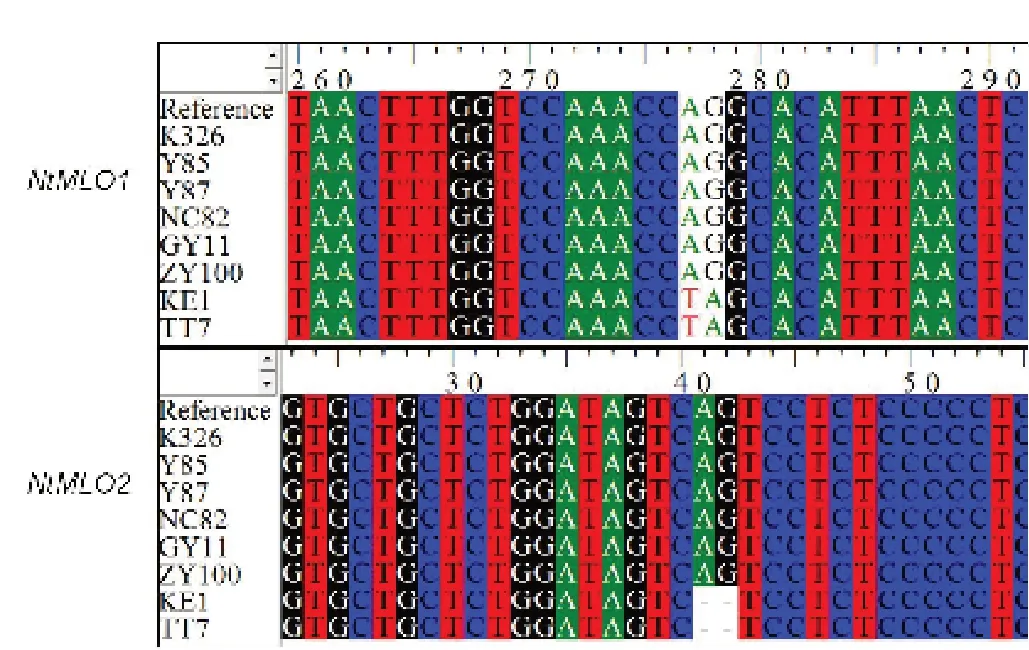
圖2 8 份供試材料中NtMLO1 和NtMLO2 基因的差異Fig. 2 The sequence differences of NtMLO1 and NtMLO2 in eight tested materials
利用抗、感材料中的基因序列差異,開(kāi)發(fā)分子標(biāo)記,標(biāo)記序列見(jiàn)表2,將差異位點(diǎn)設(shè)計(jì)到引物的3’端,同時(shí)引入1 個(gè)堿基差異,檢測(cè)不同堿基替換對(duì)擴(kuò)增效率和特異性的影響。MLO1-R 與A-F 配對(duì)擴(kuò)增野生型材料中的NtMLO1基因,MLO1-R 與a-F 配對(duì)擴(kuò)增突變型材料中的Ntmlo1基因,MLO2-R 與B-F 配對(duì)擴(kuò)增野生型材料中的NtMLO2基因,MLO2-R 與b-F 配對(duì)擴(kuò)增突變型材料中的Ntmlo2基因。結(jié)果如圖3所示,A-F5 和A-F7 均能特異的擴(kuò)增出野生型中的NtMLO1基因,而在突變型材料中無(wú)擴(kuò)增。a-F 的7 個(gè)引物都可以特異的擴(kuò)增出突變型材料中的Ntmlo1基因,而在野生型材料中無(wú)擴(kuò)增,但是a-F4 和a-F5 的擴(kuò)增效果不好。因此選用引物對(duì)A-F7/MLO1-R 和a-F7/MLO1-R 作為NtMLO1/Ntmlo1基因的分子標(biāo)記。B-F和b-F 的引物擴(kuò)增效率和特異性都很好,選擇B-F1/MLO2-R 和b-F1/MLO2-R 作 為NtMLO2/Ntmlo2基 因的分子標(biāo)記。
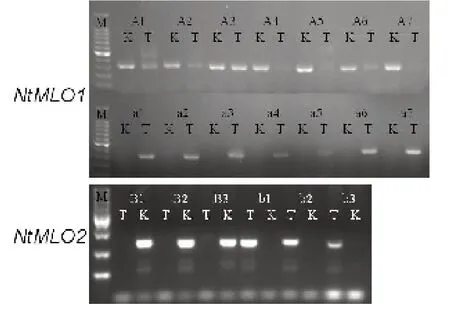
圖3 NtMLO 基因分子標(biāo)記的篩選Fig. 3 Screening of NtMLO molecular markers
2.4 分子標(biāo)記在分離群體中的應(yīng)用
使用抗病材料臺(tái)煙7 號(hào)和感病材料K326 構(gòu)建分離群體,在F2 代群體中驗(yàn)證分子標(biāo)記與抗性的關(guān)系。設(shè)置NtMLO1在感病品種中的基因型為A,在抗病品種中的基因型為a,NtMLO2在感病品種中的基因型為B,在抗病品種中的基因型為b。接種鑒定的結(jié)果發(fā)現(xiàn)只有基因型為aabb 的后代表現(xiàn)為抗病,而基因型為AABB、AABb、AAbb、AaBB、AaBb、Aabb、aaBB、aaBb 的單株均表現(xiàn)為感病。利用這四對(duì)分子標(biāo)記可以明確分離后代的基因型,在雜交育種過(guò)程中可以根據(jù)基因型來(lái)選擇后代單株(圖4)。

圖4 分子標(biāo)記在F2 分離群體中的應(yīng)用Fig. 4 Application of molecular markers in the F2 segregation population
2.5 分子標(biāo)記在回交改良中的應(yīng)用
在回交改良的過(guò)程中,如果是隱性基因控制的性狀,每一代需要通過(guò)自交,鑒定后代的表型(圖5a)。利用本研究開(kāi)發(fā)的兩個(gè)抗病分子標(biāo)記(a 和b)對(duì)回交后代的基因型進(jìn)行鑒定,不需要通過(guò)自交鑒定表型,即可篩選到攜帶抗性位點(diǎn)(a和b)的后代單株(圖5b)。
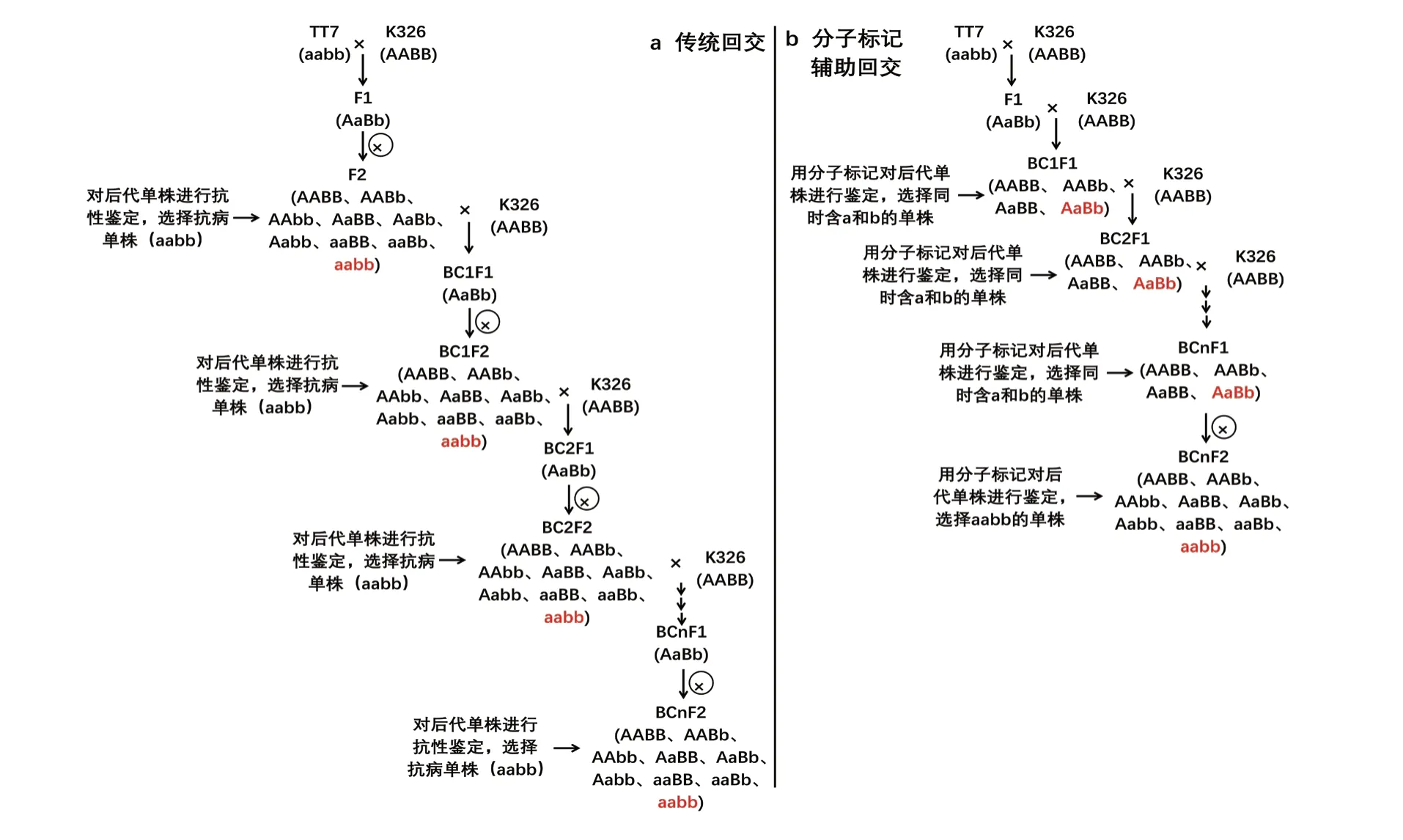
圖5 分子標(biāo)記輔助回交改良與傳統(tǒng)回交改良進(jìn)程比較Fig. 5 Comparison of molecular marker assisted backcross improvement process and traditional backcross improvement process
以回交改良K326 的BC3F1 代為例,其后代存在AABB、AABb、AaBB、AaBb 四種基因型,通過(guò)a-F7/MLO1-R 和b-F1/MLO2-R 分子標(biāo)記,對(duì)后代基因型進(jìn)行鑒定(圖6),選擇兩對(duì)引物均有擴(kuò)增,即同時(shí)攜帶a 和b 抗性位點(diǎn)(基因型為AaBb)的單株BC3F1-5、7、9 進(jìn)行套袋收種。
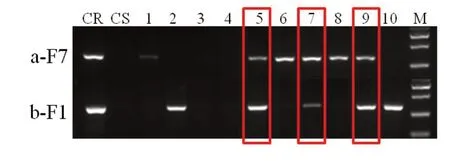
圖6 分子標(biāo)記在早代回交改良中的應(yīng)用Fig. 6 Application of molecular markers in the early backcross improvement process
高代回交材料開(kāi)展自交后,開(kāi)展抗性純合位點(diǎn)篩選結(jié)合遺傳背景鑒定,可獲得背景回復(fù)率高且攜帶抗性位點(diǎn)的純合株系。為獲得盡可能多的抗性單株供遺傳背景鑒定,需進(jìn)行較大規(guī)模的單株標(biāo)記鑒定,此時(shí),采用逐步淘汰法可顯著減輕工作量。以BC6F2 為例,先使用A-F7/MLO1-R 標(biāo)記進(jìn)行篩選(圖7A),去除掉攜帶A 的單株(約占3/4),使用B-F1/MLO2-R標(biāo)記進(jìn)行篩選(圖7B),去除掉攜帶B 的單株(約占3/4),篩選出來(lái)的單株BC6F1-4 和27,使用a-F7/MLO1-R 和b-F1/MLO2-R 標(biāo)記進(jìn)行驗(yàn)證(圖7C),BC6F2-4 和27 兩個(gè)單株均有擴(kuò)增,基因型為aabb。
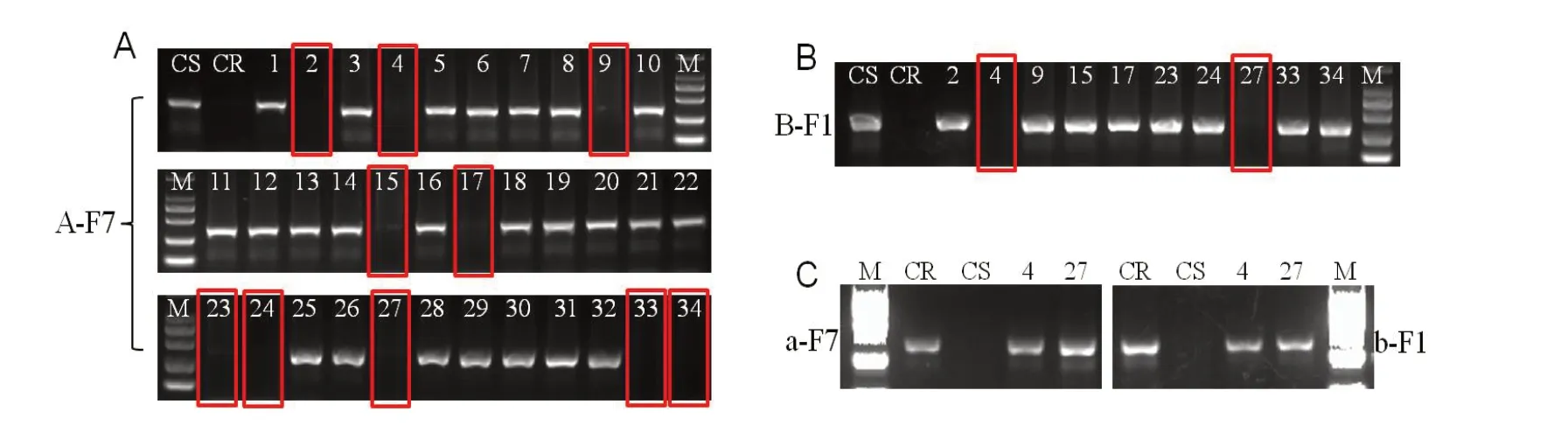
圖7 分子標(biāo)記在回交改良純合世代中的應(yīng)用Fig. 7 Application of molecular markers in the last backcross improvement process
3 討論
煙草白粉病是由二孢白粉菌(Erysiphe cichoracearumDC.)引起的嚴(yán)重病害,該病對(duì)煙草危害很大,發(fā)病嚴(yán)重時(shí),白粉覆蓋葉片,使煙葉喪失經(jīng)濟(jì)價(jià)值。煙草白粉病在我國(guó)各主要產(chǎn)煙省(自治區(qū))均有發(fā)生,而且近年來(lái)發(fā)生嚴(yán)重和區(qū)域擴(kuò)大的趨勢(shì),已經(jīng)由煙草生產(chǎn)上的次要病害發(fā)展成為影響煙草生產(chǎn)的主要病害之一。因此開(kāi)展煙草白粉病相關(guān)基因的研究及抗白粉病材料的創(chuàng)制對(duì)于該病的控制和提高煙葉質(zhì)量具有重要意義。
研究發(fā)現(xiàn),MLO基因的突變會(huì)使植物獲得廣譜持久的白粉病抗性[18],創(chuàng)制和利用MLO基因的突變來(lái)改良植物的白粉病抗性具有重要價(jià)值。目前,在大麥[9]、小麥[19-20]、番茄[10]、茄子[21]、蘋(píng)果[22]等植物中,都獲得了MLO基因自然突變、誘變或基因編輯的抗白粉病材料。煙草中NtMLO1和NtMLO2兩個(gè)感病基因在白粉病抗性中發(fā)揮了重要作用[12,14],本文通過(guò)對(duì)8 份煙草資源進(jìn)行了白粉病接種和抗性鑒定,獲得了臺(tái)煙7 號(hào)和KE1 兩份抗病材料,進(jìn)一步分析發(fā)現(xiàn)2份材料的白粉病抗性也是由于NtMLO1和NtMLO2基因同時(shí)發(fā)生了突變而獲得的。
開(kāi)展白粉病抗源材料的高效利用和抗性轉(zhuǎn)移對(duì)煙草白粉病抗性育種具有重要價(jià)值,但由于栽培煙草是異源四倍體,NtMLO1和NtMLO2存在基因功能冗余的現(xiàn)象[14],必須同時(shí)導(dǎo)入這兩個(gè)突變位點(diǎn)才能使煙草獲得白粉病抗性。通過(guò)常規(guī)育種手段開(kāi)展白粉病抗性遺傳改良,存在抗性鑒定困難、抗性易丟失、育種周期長(zhǎng)等問(wèn)題,開(kāi)發(fā)白粉病抗性連鎖分子標(biāo)記開(kāi)展白粉病輔助選擇是提高育種效率的重要手段。本研究利用抗、感材料中NtMLO基因的序列差異開(kāi)發(fā)分子標(biāo)記,利用開(kāi)發(fā)的分子標(biāo)記開(kāi)展了主栽品種的抗性回交改良,獲得了白粉病抗性顯著提高的新品系。
本文開(kāi)發(fā)的分子標(biāo)記在煙草苗期即可確定雜交后代的抗感情況,有效解決了煙草白粉病抗性鑒定易受環(huán)境影響,抗性易丟失等問(wèn)題,在煙草白粉病抗病育種中具有重要作用。該分子標(biāo)記具備以下特點(diǎn):(1)成本較低,只需進(jìn)行普通PCR 擴(kuò)增結(jié)合瓊脂糖凝膠電泳,在短時(shí)間內(nèi)即可確定抗性位點(diǎn)的基因型。(2)4 對(duì)分子標(biāo)記配合使用具有共顯性特點(diǎn),可有效區(qū)分雜交后代的雜合/純合基因型。(3)品種定向改良過(guò)程中,只需利用本文的a 和b 兩個(gè)分子標(biāo)記可以在回交過(guò)程中對(duì)攜帶雙突變基因型的后代單株進(jìn)行篩選,無(wú)需自交一代,無(wú)需接種鑒定,可縮短約一倍育種進(jìn)程,大規(guī)模單株篩選采用逐步淘汰法可顯著減少標(biāo)記鑒定的工作量。
4 結(jié)論
獲得了煙草白粉病抗源材料2 份,其中臺(tái)煙7 號(hào)的白粉病抗性符合雙基因隱性遺傳規(guī)律,開(kāi)發(fā)了與白粉病抗性共分離的分子標(biāo)記,并應(yīng)用于主栽品種的抗性回交改良,可顯著提高煙草白粉病抗病育種效率。
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國(guó)煙草學(xué)報(bào)(2019年5期)2019-11-14 07:54:12
首都公共衛(wèi)生(2019年5期)2019-05-21 01:08:34
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
自動(dòng)化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學(xué)林(2013年2期)2013-11-12 12:58:54
中國(guó)煙草學(xué)報(bào)(2012年5期)2012-04-12 06:21:18
