遺傳改造乳酸鏈球菌合成Nisin的研究進(jìn)展
2021-11-25 12:42:26羅林根夏立秋
湖南師范大學(xué)自然科學(xué)學(xué)報(bào) 2021年5期
羅林根,穰 杰,夏立秋*
(1.湖南師范大學(xué)生命科學(xué)學(xué)院,中國 長沙 410081;2.臨湘市第一中學(xué),中國 岳陽 414000)
乳酸鏈球菌素( Nisin)是由乳酸鏈球菌產(chǎn)生的一類小分子肽,具有廣譜抑菌作用,不僅對(duì)致病性病原體如李斯特氏菌、金黃色葡萄球菌、芽胞桿菌等革蘭氏陽性菌具有抗菌活性,還能緩解由部分微生物引起的食品腐敗變質(zhì)問題。由于Nisin具有抑菌效果好、熱穩(wěn)定性高以及對(duì)人安全無毒等優(yōu)點(diǎn),早在1969年就被世界衛(wèi)生組織(WHO) 批準(zhǔn)用作食品防腐劑,隨后于1988年被美國食品藥品監(jiān)管局(Food Drug Administration,FDA)列入GRAS (一般認(rèn)為安全) 名單。目前,Nisin作為一種國際公認(rèn)的安全無毒的天然防腐劑,已被商業(yè)化廣泛應(yīng)用[1]。
Nisin的分子結(jié)構(gòu)已于1971年得到解析,其分子式為C143H228N42O37S7,相對(duì)分子質(zhì)量為3 500,一般以二聚體(7 000)或四聚體(14 000)結(jié)構(gòu)存在。Nisin主體結(jié)構(gòu)由34個(gè)氨基酸殘基構(gòu)成,并通過硫醚鍵連接成五環(huán)結(jié)構(gòu),共涉及11種常見氨基酸和4種稀有氨基酸。在此基礎(chǔ)上,通過氨基酸修飾可產(chǎn)生不同Nisin變體。目前,已有8種天然Nisin變體(Nisin A,Nisin Z,Nisin F,Nisin Q, Nisin H,Nisin P,Nisin U和Nisin U2)被報(bào)道,其中Nisin A和Nisin Z研究最為廣泛[2]。
Nisin作為一種無毒的天然防腐劑,需求量一直持續(xù)增長,然而由乳酸鏈球菌生產(chǎn)的Nisin存在發(fā)酵活力、產(chǎn)量較低以及生產(chǎn)成本高等問題,嚴(yán)重限制它的商業(yè)化推廣應(yīng)用。發(fā)酵培養(yǎng)基優(yōu)化是促進(jìn)乳酸鏈球菌合成Nisin的重要手段。吳疆等根據(jù)代謝發(fā)酵控制原理,采用二次回歸正交旋轉(zhuǎn)組合設(shè)計(jì),研究了乳酸鏈球菌搖瓶發(fā)酵培養(yǎng)基中蔗糖、蛋白胨、酵母浸出物以及KH2PO3等4種主要成分對(duì)Nisin產(chǎn)量的影響,通過DPS軟件優(yōu)化其添加比例,顯著提高Nisin的產(chǎn)量[3]。通過培養(yǎng)基優(yōu)化雖然能促進(jìn)Nisin的合成,但其產(chǎn)量遠(yuǎn)沒有達(dá)到工業(yè)化生產(chǎn)要求。目前,對(duì)參與目標(biāo)天然產(chǎn)物生物合成的相關(guān)基因進(jìn)行定向遺傳改造或代謝途徑優(yōu)化是促進(jìn)其生物合成的主要方法,并在許多重要天然產(chǎn)物生產(chǎn)菌中廣泛應(yīng)用。
1 乳酸鏈球菌基因組特征
近年來,利用全基因組測(cè)序與生物信息學(xué)方法對(duì)乳酸鏈球菌進(jìn)行遺傳背景的解析為乳酸鏈球菌的進(jìn)一步開發(fā)提供了廣闊的研究空間。自Bolotin等[4]在2001年發(fā)布了第一株名為乳酸鏈球菌乳酸亞種IL1403菌株的全基因組序列以來,總共有235株乳酸鏈球菌及其亞種的全基因組完成圖或草圖得到公布,基因組大小在1.71~2.96 Mb,GC含量在34.5%~38.6%[5-7](表1)。以乳酸鏈球菌乳酸亞種IL1403為例,該菌株基因
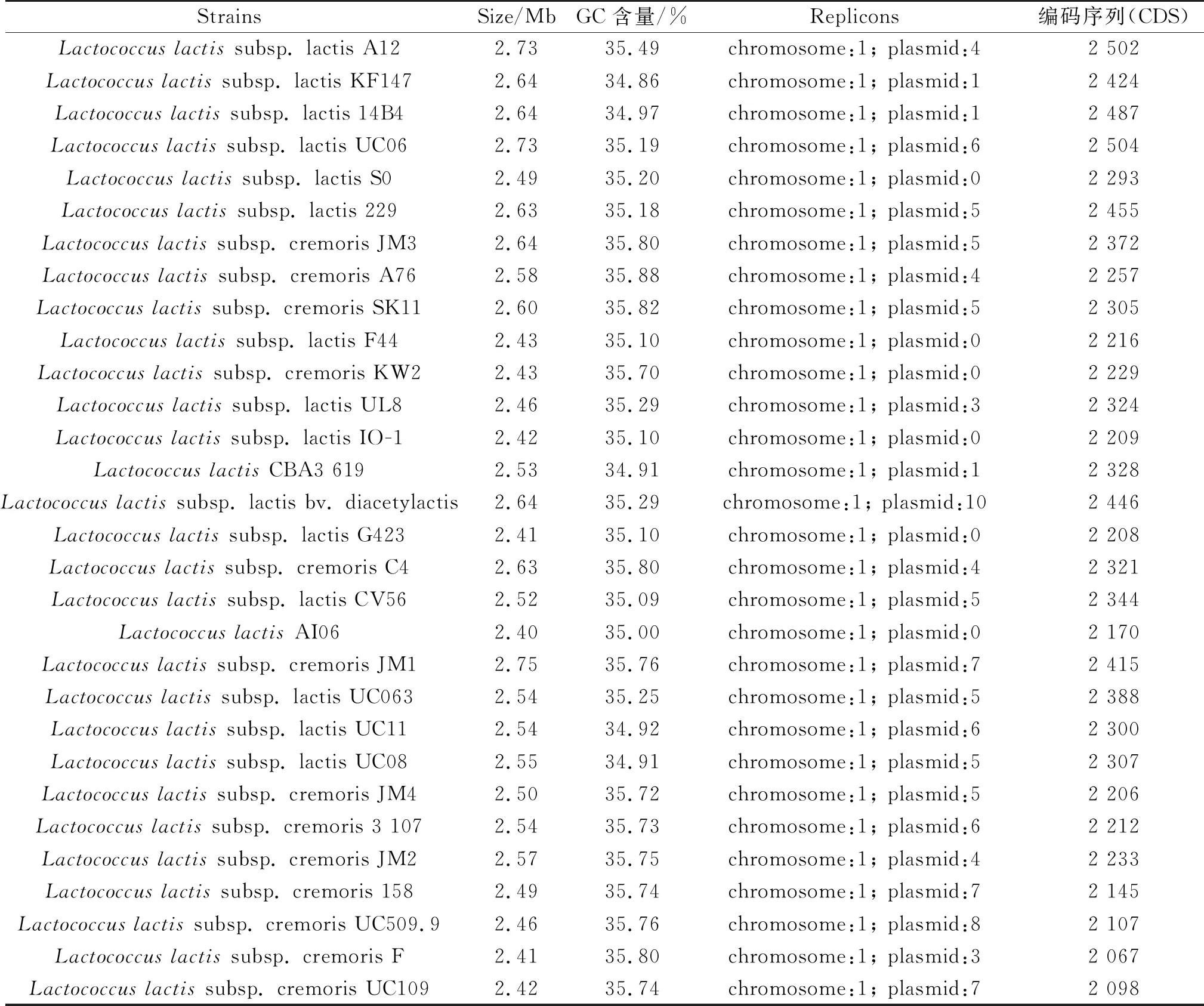
表1 部分已完成測(cè)序的乳酸鏈球菌基因組信息比較
組大小為2.37 Mb,GC含量為35.3%,共編碼2 219個(gè)蛋白質(zhì),19個(gè)rRNA,63個(gè)tRNA以及其他4個(gè)非編碼RNA。對(duì)所編碼蛋白質(zhì)的生物學(xué)功能進(jìn)行分析,發(fā)現(xiàn)該菌株含有全部20種氨基酸的生物合成途徑[4]。在對(duì)已測(cè)序的乳酸鏈球菌基因組進(jìn)一步分析發(fā)現(xiàn),部分菌株還存在多個(gè)質(zhì)粒,如乳酸鏈球菌乳酸亞種KF147的基因組由一條染色體DNA和1個(gè)質(zhì)粒組成[8],而乳酸鏈球菌乳酸亞種SD96的質(zhì)粒數(shù)目達(dá)到了10個(gè)[9],從而賦予不同乳酸鏈球菌亞種獨(dú)特的生物學(xué)功能。此外在乳酸鏈球菌基因組中還存在多個(gè)前噬菌體序列和插入序列,如乳酸鏈球菌乳酸亞種MG1363的基因組存在多達(dá)134 kb前噬菌體序列和92個(gè)插入序列[10]。這些序列的存在可引起高頻率的基因水平轉(zhuǎn)移,這可能是乳酸鏈球菌在適應(yīng)復(fù)雜的營養(yǎng)環(huán)境過程中,導(dǎo)致染色體大小發(fā)生變化的重要原因。
2 Nisin生物合成基因簇及合成途徑
全基因組測(cè)序結(jié)果顯示,在乳酸鏈球菌基因組中存在一個(gè)大小為14 kb的基因簇nisABTCIPRKFEG,并已證明這11個(gè)基因都參與了Nisin的合成[11](圖1A)。其中nisA編碼了含有57個(gè)氨基酸殘基的Nisin前體肽,隨后Nisin前體肽中的絲氨酸和蘇氨酸在脫水酶NisB和環(huán)化酶NisC的催化下脫水生成Dha和Dhb殘基并與半胱氨酸偶聯(lián),形成甲基羊毛硫氨酸環(huán)[12](圖1B)。完全修飾的Nisin前體在轉(zhuǎn)運(yùn)蛋白NisT(屬于ABC轉(zhuǎn)運(yùn)蛋白家族)的作用下可以跨膜運(yùn)輸?shù)郊?xì)胞外,同時(shí)在胞外蛋白酶NisP的作用下對(duì)Nisin前體進(jìn)行前導(dǎo)肽的切割最終形成成熟且有活性的抗菌肽[13]。由于Nisin具有強(qiáng)烈的抑菌性,在長期進(jìn)化過程中乳酸鏈球菌產(chǎn)生了一種自我保護(hù)的機(jī)制,如在脂蛋白NisI和ABC轉(zhuǎn)運(yùn)蛋白NisFEG的協(xié)同作用下,一方面通過直接結(jié)合Nisin,防止其與脂質(zhì)體Ⅱ分子結(jié)合在細(xì)胞膜上形成小孔,另一方面,ABC 轉(zhuǎn)運(yùn)蛋白 NisFEG能夠把入侵細(xì)胞膜的Nisin分子排出到細(xì)胞外,減少進(jìn)入細(xì)胞膜的Nisin分子數(shù)量,從而防止Nisin分子局部濃度過高形成孔洞等,避免自身受Nisin的侵害[14]。另外,該基因簇中還存在調(diào)控Nisin合成的蛋白,如NisR和NisK。當(dāng)細(xì)胞外出現(xiàn)成熟的Nisin分子時(shí),它會(huì)與NisK結(jié)合,引發(fā)該蛋白組氨酸磷酸化,并將磷酸基團(tuán)轉(zhuǎn)移至NisR上使其磷酸化,最終激活nis基因簇上相應(yīng)基因的轉(zhuǎn)錄調(diào)控[15]。

圖1 Nisin生物合成基因簇及其編碼產(chǎn)物(A.nis基因簇結(jié)構(gòu)及其啟動(dòng)子位置示意圖;B. Nisin結(jié)構(gòu))
3 代謝工程改造促進(jìn)乳酸鏈球菌Nisin的合成
與培養(yǎng)基優(yōu)化相比,代謝工程定向改造在提高菌株次生代謝產(chǎn)物合成能力上具有更強(qiáng)的目的性。在改造策略上可以選擇對(duì)目標(biāo)次生代謝產(chǎn)物進(jìn)行基因簇多拷貝、正調(diào)控基因過表達(dá)、負(fù)調(diào)控基因阻斷以及優(yōu)勢(shì)基因異源表達(dá)等。
3.1 增加Nisin基因簇中關(guān)鍵基因的拷貝數(shù)
研究發(fā)現(xiàn)增加直接參與Nisin生物合成基因的拷貝數(shù)可促進(jìn)Nisin的產(chǎn)量,增加參與Nisin生物合成調(diào)控與抗性基因的拷貝數(shù)(nisRK和nisFEG),也可達(dá)到相同的效果[16]。雖然Nisin在其自身的生物合成中是作為誘導(dǎo)劑發(fā)揮作用,但乳酸鏈球菌的正常生長卻受到了Nisin的不利影響。特別是在發(fā)酵的后期,生長培養(yǎng)基中大量Nisin的積累會(huì)對(duì)乳酸鏈球菌產(chǎn)生反饋抑制作用。由于乳酸鏈球菌可通過產(chǎn)生脂蛋白NisI和轉(zhuǎn)運(yùn)蛋白NisFEG來保護(hù)自己,因此,通過增加乳酸鏈球菌中Nisin耐藥基因(nisI和nisFEG)的表達(dá),可使細(xì)胞對(duì)Nisin的耐藥性增強(qiáng),從而避免自身受Nisin的影響[17]。Stein等人將含有nisI的重組載體導(dǎo)入野生型乳酸鏈球菌后,使得Nisin的產(chǎn)量提高20%,同時(shí),nisI拷貝數(shù)的增加還促使nis基因簇中其它基因的表達(dá)量增強(qiáng)[14]。有研究人員在對(duì)乳酸鏈球菌乳酸亞種164菌株進(jìn)行遺傳操作時(shí),通過對(duì)結(jié)構(gòu)基因nisZ、調(diào)控基因nisRK和抗性基因nisFEG進(jìn)行共表達(dá),使得Nisin Z的活性由16 000 AU·mL-1增強(qiáng)到25 000 AU·mL-1[18]。在其他的研究中發(fā)現(xiàn),同時(shí)提高nisRK和nisFEG的拷貝數(shù),可使Nisin的產(chǎn)量提高45%[19]。
3.2 提高乳酸鏈球菌對(duì)低pH值發(fā)酵液的耐受性
由于乳酸鏈球菌在發(fā)酵過程中會(huì)產(chǎn)生乳酸,導(dǎo)致發(fā)酵液pH值會(huì)逐漸降低,使得細(xì)胞出現(xiàn)自溶或釋放蛋白酶,進(jìn)而抑制Nisin產(chǎn)量,因此提高菌株對(duì)乳酸的耐受性對(duì)提高其產(chǎn)量有促進(jìn)作用[20]。解決這個(gè)問題的關(guān)鍵是克隆和表達(dá)丙酮酸脫羧酶和醇脫氫酶基因,以促進(jìn)乳酸鏈球菌將碳水化合物代謝生成乙醇。Wardani等人的研究結(jié)果顯示,通過相應(yīng)修飾可使Nisin的產(chǎn)量提高1.7倍[21]。同時(shí)有研究發(fā)現(xiàn)通過增加天冬酰胺合成酶基因的拷貝數(shù)來提高乳酸鏈球菌細(xì)胞壁組成中D-Asp酰胺化的比例,可使重組菌株對(duì)環(huán)境中酸性條件的耐受性明顯提高,從而提高Nisin的產(chǎn)量。在分批發(fā)酵和補(bǔ)料分批發(fā)酵中,重組菌株F44A的Nisin產(chǎn)量分別提高到3 405 IU·mL-1和5 346 IU·mL-1[22]。此外,Zhang等將17個(gè)與耐酸相關(guān)的基因在乳酸鏈球菌F44中進(jìn)行異源表達(dá)或過表達(dá)來維持細(xì)胞內(nèi)最佳pH值。其中,過表達(dá)hdeAB,ldh和murG基因的工程菌株在發(fā)酵過程中,菌株的耐酸能力顯著提高,細(xì)胞內(nèi)pH值更穩(wěn)定且Nisin產(chǎn)量提高到5 560 IU·mL-1[23]。
3.3 提高乳酸鏈球菌整體代謝能力
一般而言,Nisin的產(chǎn)量取決于發(fā)酵過程中乳酸鏈球菌的生物量,高濃度、高活性生產(chǎn)細(xì)胞的存在可顯著促進(jìn)目標(biāo)代謝產(chǎn)物的合成。因此,有許多研究旨在增加發(fā)酵條件下的乳酸鏈球菌基礎(chǔ)代謝能力,從而獲得生產(chǎn)性能高的工程菌。如Papagianni等人[24]將黑曲霉中與能量代謝相關(guān)的aox1基因在乳酸鏈球菌中進(jìn)行異源表達(dá),使細(xì)胞產(chǎn)生更多的能量,加速細(xì)胞分裂和代謝,最終使菌株生物量由3.2 g·L-1提高到5.8 g·L-1,Nisin的產(chǎn)量由5 900 IU·mL-1增加到7 900 IU·mL-1。他們又將pfk13-pkaC-aox1基因同時(shí)在乳酸鏈球菌中進(jìn)行表達(dá),通過提高菌株對(duì)發(fā)酵液中碳源的利用和能量生成,使得菌株生物量達(dá)到7.5 g·L-1,Nisin活性達(dá)到14 000 IU·mL-1。除上述研究來促進(jìn)Nisin的合成外,有研究人員還借助基因組重排技術(shù)來提高Nisin產(chǎn)量,所獲得的突變株F44能耐受高濃度的葡萄糖(8%~15%)和Nisin (5 000~14 000 IU·mL-1),最終使突變株Nisin的產(chǎn)量提高了2.4倍[25]。
4 合成生物學(xué)技術(shù)在改造乳酸鏈球菌合成Nisin中的策略
單個(gè)基因修飾在一定程度上能夠促進(jìn)目標(biāo)次生代謝產(chǎn)物生物合成,但次生代謝產(chǎn)物合成途徑通常相對(duì)復(fù)雜,受到多個(gè)代謝途徑和轉(zhuǎn)錄因子共同控制。通過對(duì)單個(gè)基因進(jìn)行修飾可能只會(huì)增加部分酶的表達(dá)量,對(duì)整個(gè)代謝途徑的影響不明顯,甚至中間產(chǎn)物的過度積累有時(shí)可能會(huì)對(duì)Nisin生物合成起抑制作用,導(dǎo)致難以按照確定的目標(biāo)進(jìn)行設(shè)計(jì)或設(shè)計(jì)的系統(tǒng)不穩(wěn)定,因此,亟需采用新的遺傳改造策略來提高Nisin的產(chǎn)量。近年來,合成生物學(xué)的興起和發(fā)展使得對(duì)微生物進(jìn)行遺傳改造進(jìn)入到一個(gè)全新的階段,研究主流也從單一基因的設(shè)計(jì)擴(kuò)展到對(duì)多個(gè)代謝途徑進(jìn)行改造。通過深度挖掘高效功能元件,重構(gòu)代謝網(wǎng)絡(luò),優(yōu)化元件與底盤的適配性,并對(duì)代謝網(wǎng)絡(luò)流量進(jìn)行精細(xì)調(diào)控,從而構(gòu)建基于人工基因線路的定制化細(xì)胞來實(shí)現(xiàn)藥物與功能材料或目的化合物的大規(guī)模生產(chǎn)及應(yīng)用[26]。
4.1 優(yōu)勢(shì)底盤細(xì)胞構(gòu)建
在特定環(huán)境中,維持細(xì)菌正常代謝所需的必需基因構(gòu)成了該菌株的最小基因組。采用最小基因組作為底盤微生物進(jìn)行目標(biāo)產(chǎn)物的合成,可減少本底其他不必要的代謝路徑對(duì)底物、能量和還原力的消耗。已公布的微生物全基因組測(cè)序結(jié)果顯示,基因組中存在大量的編碼非目標(biāo)產(chǎn)物的基因簇、前噬菌體序列、轉(zhuǎn)座酶和插入序列等,這些基因元件的存在很可能干擾目標(biāo)產(chǎn)物的合成與檢測(cè),同時(shí)導(dǎo)致菌株遺傳物質(zhì)不穩(wěn)定[27]。因此優(yōu)勢(shì)底盤細(xì)胞的開發(fā)最重要的策略就是對(duì)目標(biāo)菌株進(jìn)行基因組精簡(jiǎn)和優(yōu)化,并在很多微生物合成生物學(xué)改造中得到了廣泛應(yīng)用。如枯草芽胞桿菌PG10與野生型枯草芽胞桿菌相比,其基因組已縮減36%,但該突變株克服了與分泌過程和分泌產(chǎn)物不穩(wěn)定性有關(guān)的蛋白質(zhì)生產(chǎn)的幾個(gè)瓶頸,顯著提高細(xì)菌表達(dá)宿主分泌蛋白的產(chǎn)量[28]。研究發(fā)現(xiàn)慢病毒在大腸桿菌中進(jìn)行克隆時(shí)往往表現(xiàn)出極大的不穩(wěn)定性,導(dǎo)致難以獲得有效的克隆,Chakiath等人[29]對(duì)大腸桿菌基因組進(jìn)行縮減后,有效地促進(jìn)了慢病毒表達(dá)載體在細(xì)胞內(nèi)的重組穩(wěn)定性。Zhou等人[30]敲除了天藍(lán)色鏈霉菌體內(nèi)所有的10個(gè)PKS,NRPS基因簇以及900 kb的端粒序列,并在該底盤內(nèi)實(shí)現(xiàn)了放線紫紅素的過表達(dá)。已公布的乳酸鏈球菌全基因組測(cè)序結(jié)果顯示,其基因組不僅存在一定的可塑性(1.71~2.96 Mb),還包括多個(gè)非必須基因區(qū)域、前噬菌體序列和插入序列等,對(duì)其進(jìn)行精減,可極大減少本底非必須代謝途徑對(duì)底物、能量和還原力的消耗以及維持菌株的遺傳穩(wěn)定,從而顯著促進(jìn)Nisin的合成。
4.2 前體合成途徑優(yōu)化
目標(biāo)產(chǎn)物在宿主細(xì)胞中合成不足,很有可能是由于宿主內(nèi)某些底物或前體供應(yīng)不足。可以通過組合生物合成的方法對(duì)宿主菌代謝途徑進(jìn)行改造和優(yōu)化,以滿足目標(biāo)產(chǎn)物合成所需底物和前體的供應(yīng)。如果通過基因組精簡(jiǎn)來敲除和沉默原有基因簇的手段是“節(jié)流”,那么通過改造和優(yōu)化代謝途徑來增加底物和前體含量的方法可稱之為“開源”,這種方法需要對(duì)宿主菌的整個(gè)代謝網(wǎng)絡(luò)有比較深入的了解,才能夠進(jìn)行合理的改造。孫偉康等人[31]基于基因組功能注釋和比較基因組學(xué)構(gòu)建了乳酸鏈球菌的首個(gè)基因組規(guī)模代謝網(wǎng)絡(luò)模型,該模型共涉及557個(gè)基因,668個(gè)代謝物,840個(gè)反應(yīng),并進(jìn)一步在定性和定量?jī)蓚€(gè)層次驗(yàn)證了模型的準(zhǔn)確性,較為全面地揭示了乳酸鏈球菌的代謝特點(diǎn),為乳酸鏈球菌的前體合成途徑改造和優(yōu)化提供了良好的工具。基于Nisin的結(jié)構(gòu)解析可知,其生物合成至少涉及14種常見氨基酸和4種稀有氨基酸。結(jié)合基因組規(guī)模代謝網(wǎng)絡(luò)模型,可將涉及上述氨酸酸合成的代謝通路增強(qiáng)、競(jìng)爭(zhēng)性代謝通路阻斷等來優(yōu)化前體合成途徑,從而提高Nisin的合成能力。
5 總結(jié)與展望
Nisin是唯一被批準(zhǔn)用于食品工業(yè)的細(xì)菌素,其主要用于肉制品、乳制品、生鮮果蔬的保鮮。現(xiàn)階段Nisin主要通過微生物發(fā)酵生產(chǎn),然而野生型菌株Nisin的合成量遠(yuǎn)不能滿足工業(yè)化生產(chǎn)需求。發(fā)酵工藝優(yōu)化以及對(duì)關(guān)鍵功能基因進(jìn)行過表達(dá)是促進(jìn)乳酸鏈球菌合成Nisin的重要措施,但并沒有顯著提高Nisin的產(chǎn)量。近年來,合成生物學(xué)技術(shù)在提高目標(biāo)產(chǎn)物產(chǎn)量上有著發(fā)酵工藝和代謝工程改造所無法具備的優(yōu)點(diǎn),也是研究者們不斷關(guān)注的熱點(diǎn)。多株乳酸鏈球菌全基因組測(cè)序數(shù)據(jù)的公布以及基于乳酸鏈球菌基因組規(guī)模代謝網(wǎng)絡(luò)模型的構(gòu)建,使得研究人員能夠?qū)⑷樗徭溓蚓鳛橐粋€(gè)整體來研究,從而獲得與Nisin生物合成密切相關(guān)的合成途徑與代謝調(diào)控網(wǎng)絡(luò),為挖掘高效功能元件、確認(rèn)合成途徑中關(guān)鍵功能基因和代謝途徑奠定重要基礎(chǔ)。多種基因組編輯技術(shù)的出現(xiàn),如CRISPR/Cas9技術(shù)、Red/ET同源重組技術(shù)、Cre/loxP位點(diǎn)特異性重組技術(shù)等,可為乳酸鏈球菌進(jìn)行合成生物學(xué)改造提供重要編輯工具,為促進(jìn)乳酸鏈球菌Nisin的生物合成帶來全新的機(jī)遇。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中國果業(yè)信息(2019年10期)2019-11-13 01:21:34
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國衛(wèi)生(2016年3期)2016-11-12 13:23:26
