糞腸球菌內源性質粒的序列分析及其穿梭載體的構建
2021-12-16 02:21:20郭建軍魏國汶
食品工業科技 2021年23期
袁 林,曾 靜,郭建軍,魏國汶
(江西省科學院微生物研究所, 江西南昌 330096)
乳酸菌是一群革蘭氏陽性、能夠發酵碳水化合物、以乳酸為主要代謝產物的各類細菌統稱,是人與大多數動物腸道內的常見優勢菌群[1?5]。乳酸菌被廣泛地應用于食品工業以及飼料工業等,是全世界公認的安全級(Generally Recognized as Safe, GRAS)食品微生物,并逐漸成為微生物學研究的模式生物[6]。現階段研究與開發乳酸菌食品級載體-受體系統,探索重要基因在乳酸菌中的克隆表達,已成為乳酸菌分子生物學研究的熱點[7?11]。
質粒是細菌細胞中染色體之外,能夠自主復制的共價閉合環狀雙螺旋的DNA分子或遺傳因子。已有研究表明,乳酸菌可攜帶數個質粒,大小不一,其中多數質粒為隱蔽性質粒,少數質粒則具有某些特殊功能[12]。乳酸菌的一些特性與其所攜帶的質粒相關,并且這些特性可以通過質粒穩定轉移至其他乳酸菌或其他細菌中[13]。目前已有許多研究對乳酸菌內源性質粒進行序列測定和功能分析,并利用乳酸菌內源性質粒上重要元件構建乳酸菌載體,其中獲得乳酸菌內源性質粒的復制子是構建乳酸菌載體的關鍵[14?16]。例如,Fang 等[17]對來源于植物乳桿菌Lactobacillus plantarumLP3的質粒pLP3進行分離及測序分析,并基于質粒pLP3的復制子成功構建乳酸菌表達載體;Zuo等[18]對來源于糞腸球菌Enterococcus faecalisML21的高拷貝數質粒pML21進行分離及測序分析,然后采用分子生物學技術將質粒pML21的復制子與大腸桿菌-雙歧桿菌穿梭載體pAM1上包含紅霉素抗性基因Em和大腸桿菌復制子的DNA片段相融合,獲得了大腸桿菌-糞腸球菌穿梭載體。
本研究從本實驗室前期分離純化的具有優良益生特性的糞腸球菌Enterococcus faecalisEXW27中分離得到了內源性質粒pXW,擬對其進行DNA序列的測定及分析,確定質粒pXW的最小復制子,然后利用質粒pXW的復制子構建大腸桿菌-乳酸菌穿梭載體,并研究大腸桿菌-乳酸菌穿梭載體的宿主范圍、轉化效率和穩定性。本研究所涉及的糞腸球菌E.faecalisEXW27的內源性質粒pXW可以為乳酸菌基因操作提供新工具,并可以為構建具有自主知識產權的乳酸菌表達載體提供基礎元件。
1 材料與方法
1.1 材料與儀器
大腸桿菌Escherichia coliDH5α、糞腸球菌E.faecalisEXW27、乳脂乳球菌LactococcuscremorisMG1363、乳酸乳球菌乳酸亞種Lactococcus lactissubsp.lactisLM0230、嗜酸乳桿菌Lactobacillus acidophilusXW118、植物乳桿菌Lactiplantibacillus plantarumXW113、鼠李糖乳桿菌Lacticaseibacillus rhamnosusDB、副干酪乳桿菌Lacticaseibacillus paracaseiLG1、乳酸片球菌Pediococcus acidilacticiXW28 均由本實驗室保存;KOD-Plus-neo DNA聚合酶 日本Toyobo公司;DNA限制性內切酶、T4 DNA連接酶、DNA Marker、蛋白質Marker、質粒DNA小量純化試劑盒、DNA膠回收試劑盒 日本Takara公司;MRS肉湯培養基、M17肉湯培養基 杭州百思生物技術有限公司;實驗所用試劑 均為分析純。
PCR 儀 (Mastercycler gradient) 美國 Eppendorf公司;QuantStudio 實時熒光定量 PCR 儀 美國Thermo Fisher Scientific 公司;TY04S-3C 凝膠成像系統 北京君意東方電泳設備有限公司;SP-752PC 紫外可見分光光度計 上海光譜儀器有限公司。
1.2 實驗方法
1.2.1 引物設計與合成 大腸桿菌-乳酸菌穿梭質粒構建過程中以及糞腸球菌內源性質粒拷貝數測定過程中涉及的引物及其序列見表1,引物由生工生物工程(上海)股份有限公司合成。

表1 引物信息Table 1 Primer sequences
1.2.2 質粒的提取與序列測定 挑取MRS平板上單菌落,接種于100 mL新鮮的 MRS 液體培養基中,37 ℃靜置過夜培養。轉接至100 mL新鮮的MRS 液體培養基中,37 ℃靜置培養24 h。8000 g離心10 min收集菌體,首先用10 mmol/L Tris-HCl、5 mmol/L EDTA,pH8.0 緩沖液洗滌菌體沉淀,然后用含有30 mg/mL溶菌酶溶液(10 mmol/L Tris-HCl、5 mmol/L EDTA,pH8.0)重新懸浮菌體,并于 37 ℃處理1 h,最后采用 Takara 質粒 DNA 小量純化試劑盒提取質粒。利用瓊脂糖凝膠電泳確認所提取質粒的大小,并將分離純化的質粒送至生工生物工程(上海)股份有限公司進行序列測定。
1.2.3 質粒的序列分析及注釋 將測序完成的DNA 序列用 SnapGene Viewer(version 2.3.2)軟件進行人工拼接,繪制質粒圖譜,標注重要酶切位點并分析質粒DNA序列中的G+C含量。采用NCBI網站的 ORF finder (https://www.ncbi.nlm.nih.gov/orffinder/)查找質粒的開放閱讀框(open reading frame,ORF),結合質粒啟動子預測網站Promoter Pridition(http://www.fruitfly.org/seq_tools/promoter.html)確定各開放閱讀框的啟動子序列。采用NCBI網站的 BLAST(https://blast.ncbi.nlm.nih.ov/Blast.cgi)對各開放閱讀框所編碼的蛋白質序列進行比對分析,預測蛋白質的功能。
1.2.4 質粒拷貝數的測定
1.2.4.1 糞腸球菌生長曲線的測定 挑取MRS平板上單菌落,接種于100 mL新鮮的MRS液體培養基中,37 ℃靜置過夜培養。轉接至100 mL新鮮的MRS液體培養基中,37 ℃靜置培養24 h。每間隔2 h取1 mL培養液,測定培養液于600 nm的吸光值,測定糞腸球菌的生長曲線。
1.2.4.2 糞腸球菌總DNA的提取 挑取MRS平板上單菌落,接種于100 mL新鮮的MRS液體培養基中,37 ℃靜置過夜培養。轉接至100 mL新鮮的MRS液體培養基中,37 ℃靜置培養24 h。8000 g離心10 min收集菌體,首先用10 mmol/L Tris-HCl、5 mmol/L EDTA,pH8.0緩沖液洗滌菌體沉淀,參照《分子克隆實驗指南》[19]中細菌總DNA提取方法提取菌體的總DNA。
1.2.4.3 質粒拷貝數的測定 采用相對定量 PCR法[20]測定內源性質粒在糞腸球菌EXW27中的拷貝數。選取糞腸球菌基因組上單拷貝基因gyrB作為內參基因,內源性質粒上單拷貝基因repB作為目的基因。針對基因gyrB和repB分別設計熒光定量PCR引物:gyrB-F、gyrB-R、repB-F、repB-R。引物對的設計要求是引物對的擴增效率均高于95%,并且兩者的數值相差低于1%。采用所設計引物對以細菌總DNA為模板進行熒光定量PCR,并采用瓊脂糖凝膠電泳檢測熒光定量PCR反應的產物。對糞腸球菌總DNA進行系列梯度稀釋,取已稀釋的DNA樣品作為模板,測得各樣品的CT值,將CT值與log10C0(C0為稀釋度)做標準曲線。將所得到的引物擴增標準曲線的斜率代入引物擴增效率E的計算公式(公式1)中,分別計算得到各引物對的擴增效率。取不同生長階段的糞腸球菌菌體,并按照如上方法提取總DNA,將總DNA等比稀釋至0.1~1 ng/μL,作為熒光定量PCR的模板。采用所設計引物進行熒光定量PCR,讀取熒光定量PCR反應的CT值,并代入公式2計算質粒的拷貝數。

式中,k為擴增擴增標準曲線的斜率。

式中,E為引物的擴增效率,△CT為用目的基因相關的引物和內參基因相關的引物進行熒光定量PCR擴增得到的CT值的差值。
1.2.5 質粒最小復制子的鑒定
1.2.5.1 大腸桿菌-糞腸球菌穿梭載體的構建 通過分析質粒的開放閱讀框,初步確定質粒上的復制子信息。設計不同引物對(如表1所示),采用PCR技術擴增不同長度的含復制子的DNA片段。擴增F1片段所用引物對為F1-F、F1-R;擴增F2片段所用引物對為F2-F、F1-R;擴增F3片段所用引物對為F3-F、F1-R;擴增F4片段所用引物對為F2-F、F4-R;擴增F5片段所用引物對為F2-F、F5-R;擴增F6片段所用引物對為F2-F、F6-R。以大腸桿菌-糞腸球菌穿梭載體pXWM1的構建為例,采用引物對F2-F、F5-R擴增F1片段,采用引物對pAM1-F、pAM1-R擴增大腸桿菌-雙歧桿菌穿梭載體pAM1上包含紅霉素抗性基因和大腸桿菌復制子的DNA片段。將得到的兩種DNA片段均用BamHI和XhoI雙酶切處理,酶切產物回收后酶連轉化大腸桿菌DH5α感受態細胞,挑取轉化子酶切驗證,獲得重組質粒pXWM1。
1.2.5.2 糞腸球菌感受態制備及電擊轉化 糞腸球菌感受態制備及電擊轉化參照文獻[21?23]進行。挑取 MRS 平板上單菌落,接種于100 mL新鮮的 GM17液體培養基 [0.5%(g/100 mL)葡萄糖、M17液體培養基]中,30 ℃靜置培養過夜。將過夜培養物以1%的接種量接種于100 mL新鮮的 SGM17 液體培養基 [0.5 mol/L蔗糖、0.5%(g/100 mL)葡萄糖、1%(g/100 mL)甘氨酸、M17 液體培養基 ]中,30 ℃靜置培養5~6 h。離心收集菌體沉淀,用冰浴的電擊緩沖液(0.5 mol/L蔗糖、10%甘油)洗滌兩次后,重懸于100 μL電擊緩沖液中,放置在冰上待用。向100 μL 感受態細胞中加入100 ng 重組質粒,輕輕混勻,置于冰上備用。將含有重組質粒的糞腸球菌感受態細胞混合液小心加入冰預冷的電轉杯中,然后進行電擊轉化。電擊轉化參數為1.8 kV、200 Ω、25 μF,脈沖時間為4.0 ms。電擊完成后,立即向電轉杯中加入900 μL新鮮的 SGM17 液體培養基,30 ℃靜置復蘇2 h。取復蘇后的菌液涂布于含2.5 μg/mL 紅霉素的 MRS 固體平板,30 ℃培養24~48 h。
1.2.6 穿梭載體的宿主范圍、轉化效率和穩定性測定
1.2.6.1 穿梭載體的宿主范圍測定 參照1.2.5.2中方法制備多種乳酸菌感受態細胞,將驗證過的能夠有效復制的穿梭載體轉化乳酸菌感受態細胞,轉化子涂布于含2.5 μg/mL紅霉素的MRS固體平板,30 ℃培養48~96 h。觀察轉化平板上是否有穿梭質粒的轉化子菌落,確定穿梭質粒的宿主范圍。
1.2.6.2 穿梭載體的轉化效率測定 分別取 20、50、100 和 500 ng 重組質粒與乳酸菌感受態細胞混勻,電擊轉化乳酸菌感受態細胞,每個濃度做3個平行。轉化子于 30 ℃ 靜置活化 3 h 后,將菌懸液稀釋至合適的濃度,涂布于含 2.5 μg/mL 紅霉素的 MRS固體平板,30 ℃ 培養 48 h 至菌落長出。對平板菌落進行計數,并計算轉化效率,最終以這四個梯度中效率最高者作為穿梭載體的轉化效率。
1.2.6.3 穿梭載體的穩定性測定 挑取轉化平板上單菌落接種至含25 μg/mL的MRS液體培養基中,37 ℃過夜培養。再以1%接種量接種至不含紅霉素的MRS液體培養基中,37 ℃培養12 h。再次按1%接種量接種至不含紅霉素的MRS液體培養基中,37 ℃培養12 h,依次傳代培養直至60代。稀釋培養好的菌液分別涂布于含紅霉素的MRS固體平板和不含紅霉素的MRS固體平板。將兩種固體平板上的菌落數代入公式3計算質粒丟失率,并以此數據來表征質粒的穩定性。

1.3 數據圖像的處理
以Primer premier 5.0的引物設計軟件進行相關引物的設計,以Tanon 1600全自動數碼凝膠成像系統對DNA瓊脂糖電泳凝膠進行采集及分析。
2 結果與分析
2.1 質粒的分離與序列測定
從糞腸球菌EXW27中分離得到質粒,并對其進行瓊脂糖凝膠電泳檢測,結果如圖1所示。瓊脂糖凝膠電泳結果表明從糞腸球菌EXW27提取得到的質粒為單一條帶,大小約為8000 bp。將此質粒命名為質粒pXW,并將分離純化的質粒樣品送至生工生物工程(上海)股份有限公司進行序列測定。

圖1 質粒pXW的瓊脂糖凝膠電泳檢測圖Fig.1 Agarose gel electrophoresis analysis of plasmid pXW
2.2 質粒的注釋
將測序完成的DNA序列用SnapGene Viewer軟件進行人工拼接,繪制質粒圖譜,如圖2A所示。質粒pXW是雙鏈環狀DNA分子,共8617 bp,GC含量為33.29%。經BLAST分析顯示:質粒pXW的核苷酸序列與來源于屎腸球菌E.faeciumFS86的質粒(GeneBank登錄號:MT501398.1)的核苷酸序列具有最高的相似性,兩者的相似率為88.37%。并且兩種質粒的核苷酸序列比對結果顯示:除了E.faeciumFS86來源質粒的質粒圖譜(圖2B)上紅色標記出的四個額外片段(片段 1、2、3、4)外,兩種質粒的核苷酸序列完全一致。其中片段1在E.faeciumFS86來源質粒上的位置為2579~2794,片段2在E.faeciumFS86來源質粒上的位置為4477~4647,片段3在E.faeciumFS86來源質粒上的位置為 6996~7323,片段 4在E.faeciumFS86來源質粒上的位置為9080~9498。以上結果表明,糞腸球菌和屎腸球菌之間可能通過水平基因轉移來獲得質粒片段或質粒。目前,有關E.faeciumFS86來源質粒的序列分析未見相關研究報道。本研究對自主分離得到的質粒pXW進行序列分析。

圖2 質粒圖譜Fig.2 Physical map
對質粒pXW進行開放閱讀框(ORF)搜索,結果顯示質粒pXW含有8個開放閱讀框。在NCBI數據庫中對這些開放閱讀框進行序列信息的比對,預測所有ORF的功能,各ORF編碼的蛋白質信息如表2所示。ORF1編碼一個含159個氨基酸殘基的蛋白質,為遷移蛋白質 MobC(Mobilization protein),該MobC蛋白的氨基酸序列與來源于屎腸球菌E.faecium24-10的MobC蛋白的氨基酸序列的相似性高達99%。ORF2編碼一個含304個氨基酸殘基的蛋白質,為遷移蛋白質MobA(Mobilization protein),該MobA蛋白的氨基酸序列與來源于屎腸球菌E.faeciumAcr4的MobA蛋白的氨基酸序列的相似性高達96%。MobC蛋白和MobA蛋白可能與質粒的遷移作用相關[24]。ORF4編碼一個含245個氨基酸殘基的蛋白質,為復制起始蛋白RepB(Replication initiation protein),該RepB蛋白的氨基酸序列與來源于E.faeciumC68的RepB蛋白的氨基酸序列的相似性高達99%。RepB蛋白的BLAST分析結果顯示,RepB蛋白屬于Rep_3超家族(pfam01051),預測與質粒的θ型復制相關[24?26]。ORF5編碼一個含178個氨基酸殘基的蛋白質,為DUF536結構域包含蛋白質(DUF536 domain-containing protein),該DUF536結構域包含蛋白質的氨基酸序列與來源于E.faecium的DUF536結構域包含蛋白質的氨基酸序列的相似性高達99%。DUF536結構域包含蛋白質屬于DUF536超家族,為未知功能蛋白質,預測其與質粒的 θ型復制相關[24?26]。ORF7編碼一個含403個氨基酸殘基的蛋白質,為轉座酶(transposase),該轉座酶的氨基酸序列與來源于E.faecium64/3 xUW2774的轉座酶的氨基酸序列的相似性高達100%。ORF8編碼一個含567個氨基酸殘基的蛋白質,為ABC轉運器ATP-結合蛋白(ABC transporter ATP-binding protein),該 ABC轉運器 ATP-結合蛋白的氨基酸序列與來源于E.faecium的ABC轉運器ATP-結合蛋白的氨基酸序列的相似性高達99%。ORF3和ORF6在NCBI數據庫中沒有搜索到與其相關的蛋白質。質粒pXW上除具有與質粒復制相關的蛋白質(復制起始蛋白RepB和DUF536結構域包含蛋白質)外,還包含有與質粒遷移作用相關的蛋白質(遷移蛋白質MobC和MobA),以及轉座酶和ABC轉運器ATP-結合蛋白,因此質粒pXW被歸為顯性質粒。質粒pXW在糞腸球菌EXW27中所起的作用有待進一步研究確證。

表2 質粒pXW的開放閱讀框信息Table 2 Open reading frame information for plasmid pXW
2.3 質粒拷貝數的測定
本研究采用相對定量PCR法測定內源性質粒pXW在糞腸球菌EXW27中的拷貝數。選取糞腸球菌基因組上單拷貝基因gyrB作為內參基因,內源性質粒上單拷貝基因repB作為目的基因。針對基因gyrB和repB分別設計熒光定量PCR引物:gyrBF、gyrB-R、repB-F、repB-R。采用所設計引物以糞腸球菌EXW27總DNA為模板進行熒光定量PCR,并用瓊脂糖凝膠電泳檢測熒光定量PCR的產物,結果如圖3所示。gyrB和repB的PCR產物大小分別為145 bp和150 bp。對糞腸球菌EXW27的總DNA進行系列梯度稀釋,取已稀釋的DNA樣品作為模板,分別采用引物對gyrB-F/gyrB-R或repB-F/repBR進行熒光定量PCR反應,測得各樣品的CT值,將CT值與log10C0(C0為樣品的稀釋度)做標準曲線,如圖4所示。引物對gyrB-F/gyrB-R的擴增標準曲線為 y=?3.4327x+2.3219,R2=0.9995;引物對repB-F/repB-R的擴增標準曲線為y=?3.4358x+5.4843,R2=0.9999。將所得的擴增標準曲線斜率代入引物擴增效率E的計算公式(公式1),計算得到引物對gyrB-F/gyrB-R和repB-F/repB-R的擴增效率分別為95.57%和95.48%。引物對的擴增效率均高于95%,而且兩者的數值相差僅為0.09%,低于1%。因此引物對gyrB-F/gyrB-R和repB-F/repB-R是可以作為相對定量PCR引物使用的。

圖3 PCR產物的瓊脂糖凝膠電泳檢測Fig.3 Agarose gel electrophoresis analysis of PCR products

圖4 引物擴增效率的標準曲線Fig.4 Standard curve of amplification efficiency of primers
取不同生長階段的糞腸球菌EXW27菌體,并提取其總DNA,將總DNA等比稀釋至0.1~1 ng/μL,作為熒光定量PCR的模板。采用所設計引物進行熒光定量PCR,讀取熒光定量PCR反應的CT值,并代入公式2計算在糞腸球菌EXW27于不同生長時期的質粒pXW拷貝數。質粒pXW的拷貝數隨糞腸球菌EXW27生長的變化趨勢(圖5)為:糞腸球菌EXW27的對數生長早中期時質粒pXW的拷貝數最高,且基本保持穩定,最高拷貝數為32.09±0.93;其對數生長后期至穩定期時質粒pXW的拷貝數逐漸降低,到生長穩定期時,質粒拷貝數穩定為12.75±0.79。因此糞腸球菌EXW27的內源性質粒pXW屬于高拷貝質粒[27]。在細菌對數生長后期,質粒拷貝數發生下降屬于普遍現象,其中的原因可能是細菌生長后期時細胞生理代謝發生變化,細胞的生長趨于停滯,胞內DAN合成減慢,內源性質粒復制所依賴的DNA聚合酶合成量也隨之下降。雖然質粒自身編碼的復制相關蛋白可以識別復制起始位點,但是缺乏必要的酶系,導致復制復合體無法形成,質粒復制受到阻礙,質粒的拷貝數下降。

圖5 質粒pXW的拷貝數與糞腸球菌EXW27生長曲線之間的關系Fig.5 Relationship between the copy number of plasmid pXW and the growth curve of E.faecium EXW27
2.4 質粒的復制方式推定以及最小復制子的鑒定
通過對質粒pXW上的開放閱讀框進行BLAST比對分析,確定了質粒上與其復制相關的復制蛋白,一個是復制起始蛋白RepB,一個是復制起始蛋白RepB下游的DUF536結構域包含蛋白質。本研究進一步對復制起始蛋白RepB的上游DNA序列進行詳細分析(如圖6所示),發現復制起始蛋白RepB的上游存在類似起始質粒復制的反向重復序列和正向重復序列,推測復制起始蛋白RepB所在區域可能為復制子區域。根據以上分析,推測質粒pXW含有1個復制子,包括復制起始子、復制起始蛋白RepB以及下游的DUF536結構域包含蛋白質。復制起始子含有1對13 bp的反向重復序列IR(Invert repeats,IR)、2個 8 bp的正向重復序列DR1(Direct repeats,DR)、3.5個 12 bp的正向重復序列DR2以及4.5個22 bp的正向重復序列DR3。復制起始蛋白RepB屬于Rep_3超家族(pfam01051)。Rep_3超家族中RepB蛋白均與質粒的θ型復制相關,例如糞腸球菌E.faecium226的質粒pMBB1為θ型復制質粒,其RepB蛋白屬于Rep_3超家族[26]。緊接其下游的DUF536結構域包含蛋白質為未知功能蛋白質, BLAST分析結果顯示其可能與質粒的θ型復制相關。此外,質粒pXW的復制子結構屬于典型的θ型質粒的復制子結構[28?29]。因此推定質粒pXW的復制類型為θ型復制。

圖6 質粒pXW的復制區域分析Fig.6 Genetic context of the replication region of plasmid pXW
為了確定質粒pXW的最小復制子,本研究構建了含有不同長度復制子片段的大腸桿菌-糞腸球菌穿梭載體,并將不同的大腸桿菌-糞腸桿菌穿梭載體分別轉化糞腸球菌感受態,驗證質粒pXW的最小復制子。如圖7所示,復制子片段F1包含一部分ORF3、完整的反向重復序列IR、完整的正向重復序列DR、ORF4、ORF5以及ORF5的下游區域;復制子片段F2包含完整的反向重復序列IR、完整的正向重復序列DR、ORF4、ORF5以及ORF5的下游區域;復制子片段F3包含完整的正向重復序列DR、ORF4、ORF5以及ORF5的下游區域;復制子片段F4包含完整的反向重復序列IR、完整的正向重復序列DR、ORF4、ORF5以及部分ORF5的下游區域;復制子片段F5包含完整的反向重復序列IR、完整的正向重復序列DR、ORF4以及ORF5;復制子片段F5包含完整的反向重復序列IR、完整的正向重復序列DR、ORF4以及部分ORF5。穿梭載體的轉化結果顯示,含有復制子片段F1、F2、F4或F5的大腸桿菌-糞腸球菌穿梭載體電擊轉化糞腸球菌感受態后有菌落長出,含有復制子片段F3或F6的大腸桿菌-糞腸球菌穿梭載體電擊轉化糞腸球菌感受態后無菌落長出。因此,對于質粒pXW的復制子,完整的反向重復序列IR、完整的正向重復序列DR、ORF4以及ORF5是必需的。質粒pXW的最小復制子包括完整的反向重復序列IR、完整的正向重復序列DR、復制起始蛋白RepB以及θ型復制相關蛋白DUF536結構域包含蛋白。

圖7 質粒pXW最小復制子的鑒定Fig.7 Minimal replicon identification of plasmid pXW
2.5 質粒的宿主范圍、轉化效率及穩定性測定
將確定最小復制子過程中構建的含有復制片段F5的穿梭載體命名為pXWM1,其質粒圖譜如圖8所示。為確定穿梭質粒pXWM1的宿主范圍,將穿梭質粒pXWM1電擊轉化至各種乳酸菌感受態細胞中,利用紅霉素標記進行篩選,以確定穿梭質粒pXWM1的宿主范圍,并計算其在各種乳酸菌中的轉化效率和穿梭質粒的穩定性。結果顯示,穿梭質粒pXWM1均可在表3所示的乳酸菌中成功電轉,并獲得對應的轉化子。這表明,質粒pXW屬于寬宿主范圍質粒。前面研究結果顯示質粒pXW被推斷為θ型復制質粒,質粒較穩定,并且質粒pXW含有質粒遷移蛋白MobC和MobA,這些因素與該質粒屬于寬宿主質粒是相符合的。
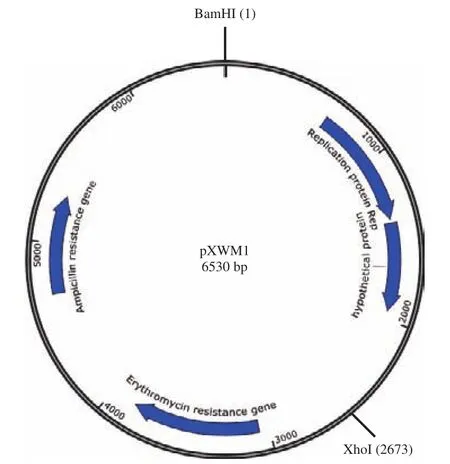
圖8 pXWM1的質粒圖譜Fig.8 Physical map of pXWM1
穿梭質粒pXWM1在表3所示的乳酸菌中的電擊轉化效率介于 1.96×102~8.96×104CFU/μg (質粒DNA)之間,比普通乳酸菌質粒轉化效率(10~105CFU/μg質粒DNA)略低。穿梭質粒比普通乳酸菌質粒轉化效率略低的可能原因是[30]:乳酸菌宿主菌中存在限制性修飾系統,排斥未甲基化的外源質粒;乳酸菌宿主菌內可能存在不親和群的質粒;不同乳酸菌宿主菌的DNA甲基化水平;以及其他限制轉化效率的因素如電擊電壓、電擊緩沖液、感受態細胞的狀態等。穿梭載體pXWM1在表3所示的乳酸菌中經過60代連續傳代培養后,穿梭質粒的丟失率介于28.54%~54.17%之間。穿梭載體pXWM1在不同乳酸菌感受態細胞中電轉化效率和質粒穩定性的差異,可能與質粒和宿主菌的相容性相關[14]。

表3 穿梭載體pXWM1在不同乳酸菌宿主中的轉化效率和穩定性Table 3 Transformation efficiency and stability of shuttle vector plasmid pXWM1 in different hosts
3 結論
從具有優良益生特性的糞腸球菌E.faecalisEXW27中分離得到了內源性質粒pXW,該質粒大小為8617 bp,GC含量為33.29%。針對該質粒DNA序列的BLAST分析結果顯示質粒pXW為新質粒。進一步通過生物信息學分析發現,該質粒編碼8個ORF,其中包括質粒遷移作用蛋白MobC(ORF1)和 MobA(ORF2)、質粒復制起始蛋白 RepB(ORF4)、θ型復制相關蛋白DUF536結構域包含蛋白(ORF5)、轉座酶(ORF7)、ABC 轉運器 ATP-結合蛋白(ORF8)以及假定蛋白質(ORF3、ORF6)。根據復制起始蛋白RepB、緊接其下游的DUF536結構域包含蛋白以及復制子的序列分析,推測該質粒的復制方式為θ型復制。質粒pXW在糞腸球菌EXW27中拷貝數最高可達32.09±0.93,表明質粒pXW屬于高拷貝數質粒。本研究確定了質粒pXW的最小復制子,并利用該復制子成功構建大腸桿菌-糞腸球菌穿梭載體pXWM1。該穿梭載體的宿主范圍寬,實現了在多種乳酸菌中的成功轉化,并且轉化效率高,質粒丟失率低。質粒pXW的這些優良特性有利于將其改造為具有自主知識產權的乳酸菌工程載體。
