逍遙散對慢性輕度不可預見應激抑郁大鼠中樞多巴胺及其受體的影響?
2021-12-24 09:41:32張歡潤岳廣欣楊婧雯吳望男聶文祎楚天云鞏子漢佘楷杰
中國中醫基礎醫學雜志 2021年11期
張歡潤, 岳廣欣, 梁 媛, 楊婧雯, 李 妍, 吳望男, 聶文祎, 楚天云, 鞏子漢, 佘楷杰
(中國中醫科學院中醫基礎理論研究所, 北京 100700)
抑郁癥以顯著而持久的心境低落為主要臨床特征,其高復發率、高自殺率對個人及社會造成了較大危害[1]。近年來,抑郁癥的發病率呈顯著上升趨勢,有學者預計,由該病所產生的相關支出將在2030年成為全球范圍內所有疾病負擔之首[2]。抑郁癥的發病機制有很多假說,單胺神經遞質學說受到廣泛認可。多巴胺(dopamine,DA)是獎賞功能最重要的神經遞質,與人的精神情緒活動密切相關,多項研究表明,抑郁癥的發病與DA的含量下降有關[3]。
目前臨床上有很多治療抑郁癥的藥物,如三環及四環類抗抑郁藥、單胺氧化酶抑制劑和選擇性 5-羥色胺再攝取抑制劑等,但僅對 1/3 的患者顯現出明顯改善作用,且藥物起效存在數周至數月的潛伏期[4]。中醫藥抗抑郁具有多靶點、療效好、不良反應少等獨特優勢。大量臨床與實驗文獻證實,肝郁脾虛證是抑郁癥最常見的證型,逍遙散是中醫治療肝郁脾虛證的經典方劑,對抑郁癥有著明確的治療效果[5,6]。課題組前期實驗發現,慢性輕度應激大鼠出現糖水攝入量減少、食欲與性欲下降等獎賞功能紊亂的癥狀表現,且伴有中樞和外周多巴胺(dopamine,DA)含量變化,逍遙散對此具有明顯的調節作用[7,8]。在前期研究基礎上,提出解郁名方逍遙散除了能夠通過調節神經-內分泌-免疫網絡、神經突觸可塑性以及腦腸軸等多方面機制發揮抗抑郁效應[5]之外,是否還能對獎賞功能產生影響這一科學性問題。因此,本研究從獎賞腦區DA及其受體與轉運體的基因與蛋白表達水平等方面,觀察逍遙散對應激抑郁大鼠的干預作用,進一步探究逍遙散治療抑郁癥的作用機制。本研究已通過中國中醫科學院中醫基礎理論研究所實驗動物倫理委員會批準,批準號2019-003。
1 材料與方法
1.1 動物
SPF級健康雄性SD大鼠60只,體質量180~220g,購于北京維通利華實驗動物技術有限公司,生產許可證號SCXK(京)2016-0006。飼養于中國中醫科學院中醫基礎理論研究所SPF級動物房,許可證號SYXK(京)2021-0017。室溫22±2 ℃,濕度34%~44%,明暗周期12 h,適應性喂養3 d后進行實驗。
1.2 中藥及其制備
逍遙散組成:柴胡15 g,當歸15 g,白芍15 g,白術15 g,茯苓15 g,生姜5 g,薄荷5 g,炙甘草7.5 g,組方選自《太平惠民和劑局方》。中藥飲片購自北京同仁堂(亳州)飲片有限責任公司,生產許可證號(皖)Y20170043,生產批號800000397。以上中藥分別加10倍、8倍、8倍水煎煮3次,過濾沉渣、濃縮、低溫真空干燥為浸膏,粉碎、低溫干燥備用。使用前稀釋為含生藥7.41 g/mL的中藥混懸液[9]。
1.3 主要試劑與儀器
DA、3,4-二羥基苯乙酸(3,4-dihydroxyphenylacelic acid,DOPAC)、高香草酸(homovanillic acid,HVA)標準品,96%(Extrasynthese,北京索萊寶生物科技有限公司);色譜柱為 XB-C18(50 mm×2.1 mm,1.8 μm,美國Welch公司);Trizol試劑(貨號15596-026,美國Invitrogen公司);所有引物均由寶生物(大連)工程有限公司合成;SYBR Green PCR熒光定量試劑盒(貨號4368708,美國Applied Biosystems公司);多巴胺受體1(Dopamine receptor 1,D1R,abcam,貨號ab20066),多巴胺轉運體(dopamine transporter,DAT,abcam,貨號ab18445), 多巴胺受體2(dopamine receptor 2,D2R,Proteintech,貨號55084-1-AP),以上一抗均購自北京永泰興成商貿有限公司。
3K15sICbIA高速泠凍離心機(上海珂淮儀器有限公司);Nexera UHPLC LC-30A高效液相色譜儀(Shimadzu),Mass Spectrometer QTRAP 4500 三重四極桿質譜儀(AB Sciex)。
1.4 分組與造模
根據課題組前期實驗方法進行應激易感大鼠的篩選,即施加2周應激因素后按糖水測試結果將應激大鼠分為應激易感大鼠(糖水消耗下降40%以上)與應激恢復大鼠(糖水消耗未減少),取應激易感大鼠納入下一步實驗,隨機分為模型組和逍遙散組[8],另設正常對照組5只/籠群養共10只。模型組與逍遙散組大鼠繼續施加4周的應激因素,連續造模時間共計6周[8]。應激與灌胃給藥過程中陸續有大鼠損耗,最后模型組10只,逍遙散組14只大鼠納入行為學實驗。
1.5 給藥方法
逍遙散組大鼠從第3周開始灌胃逍遙散混懸液,連續給藥4周。每天開始應激前30 min給藥,根據前期實驗制定最佳給藥劑量為2.224 g·kg-1·d-1,以生藥量計算為中劑量[10]。模型組和正常組大鼠灌胃相同體積純水。造模加灌胃4周后進行行為學檢測。
1.6 行為學檢測
1.6.1 曠場實驗 大鼠適應行為學室15 min后,放入曠場箱(120 mm×120 mm×45 cm)中心點,設中心點附近60 mm×60 cm區域為中央區,其余區域為周圍區。記錄各組大鼠5 min運動總距離(mm)、中央區進入次數(次)、中央區停留時間(s)、中央區運動距離(mm)、周圍區停留時間(s)和周圍區運動距離(mm)。
1.6.2 O迷宮 O迷宮是改良的高架十字迷宮,外徑95 cm、內徑85 cm,分為開臂和閉臂,閉臂高20 cm,O迷宮離地面高60 cm。實驗時將大鼠放入開臂上,頭向閉臂,觀察5 min內大鼠的開放臂停留時間(s)、開放臂運動距離(mm)、封閉臂停留時間(s)、封閉臂運動距離(mm)、5 min平均速度(mm/s)和5 min運動總距離(mm)。
1.7 液質聯用(high performance liquid chromato-graphy-mass Sp,HPLC-MS)方法檢測前額皮質與伏核DA、DOPAC、HVA濃度
行為學實驗完成后每組各取5只大鼠麻醉處死,冰上剝腦,用飛鷹牌保安刀片切出各層腦片,手術刀剝離出前額皮質與伏核。取前額皮質與伏核樣本50 mg~60 mg,加入0.1%甲酸-水混合物勻漿,離心、復溶后取上清液進樣。色譜條件:水相為0.1%甲酸水溶液(DA&DOPAC)/純(HVA),有機相為乙腈,流速0.3 mL/min,柱溫40 ℃,進樣量5.00 μL。用 Analyst(AB Sciex)軟件采集目標物的色譜圖并計算積分,以 1/X2為加權系數進行線性回歸。
1.8 熒光定量PCR檢測前額皮質與伏核D1R、D2R、DAT(SLC6A3 )mRNA表達
每組各取5只大鼠,Trizol法提取各組大鼠左側腦前額皮質與伏核的總RNA,使用Nanodrop 2000檢測RNA濃度及純度,將濃度過高的RNA進行適當比例稀釋,使其終濃度為100~500 ng/μL。采用逆轉錄試劑盒反轉錄合成cDNA,存入-20 ℃保存備用。使用SYBR Green PCR熒光定量試劑盒檢測D1R、D2R、DAT(SLC6A3)轉錄表達水平,以GAPDH作為內參。所有引物均由寶生物(大連)工程有限公司合成,2-ΔΔCt法進行相對定量分析。
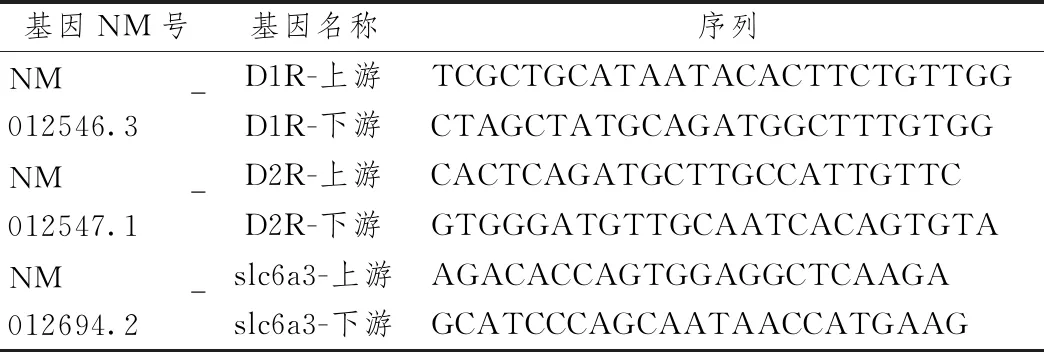
表1 引物序列表
1.9 Western blot檢測前額皮質與伏核腦區D1R、D2R、DAT蛋白表達
收集各組大鼠右側前額皮質與伏核到EP管中,提取組織總蛋白并對濃度進行測定。各組統一取蛋白樣品20 μg進行 12.5% SDS-PAGE電泳,蛋白分離并轉移至PVDF膜上。經5%脫脂牛奶37℃、封閉2 h、PBST洗膜后加入相應的一抗(D1R 1∶1000,D2R 1∶1000,DAT 1∶1000)4 ℃孵育過夜。內參為β-actin(1∶3000),PBST洗膜5×15 min,依次加入山羊抗兔的二抗(1∶3000)37 ℃ 孵育 1 h后,TBST 洗膜 5×15 min,在膜上滴加ECL顯色液后進行曝光、顯影和定影,采用PhotoShop和Alpha整理去色、分析目標光密度值。
1.10 統計學方法
2 結果
2.1 各組大鼠曠場實驗結果
表2示,慢性輕度不可預見應激模型大鼠5 min曠場實驗中央區進入次數與停留時間、中央區運動距離均有極其顯著減少(P<0.01),5 min運動總距離明顯下降(P<0.05),表明逍遙散能顯著上調中央區進入次數(P<0.05)。

表2 各組大鼠曠場實驗結果比較
2.2 各組大鼠O迷宮實驗結果
表3示,慢性輕度不可預見應激模型大鼠5 min平均速度、開放臂停留時間、開放臂運動距離、封閉臂運動距離、5 min運動總距離均有顯著下降(P<0.05,P<0.01),封閉臂停留時間明顯上升(P<0.05),除封閉臂運動距離外,逍遙散對其余指標均有顯著上調作用(P<0.05,P<0.01)。
表3 各組大鼠O迷宮實驗結果比較
2.3 各組大鼠前額皮質與伏核DA、DOPAC、HVA含量比較
圖1、2示,與正常組比較,模型組前額皮質與伏核中DA與DOPAC含量、伏核中HVA含量均明顯低于正常組(P<0.05,P<0.01),表明逍遙散能顯著上調模型組各腦區DA和HVA水平(P<0.05,P<0.01)。逍遙散組前額皮質中DOPAC含量也有顯著上升(P<0.01),伏核腦區中DOPAC水平雖有上調趨勢,但與模型組比較差異無統計學意義(P>0.05)。
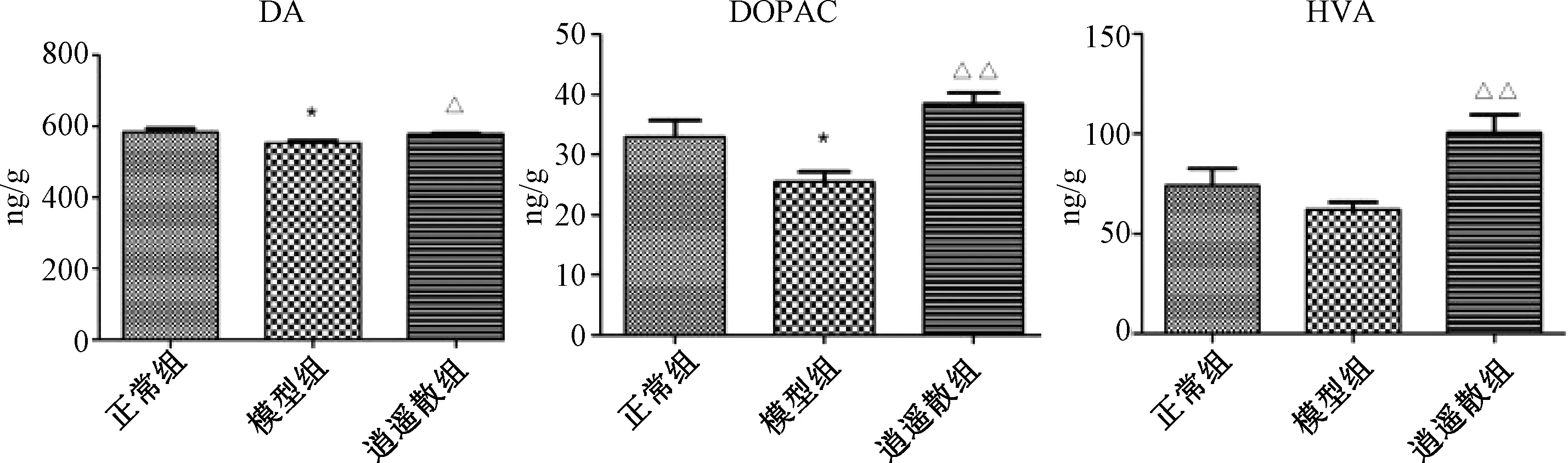
注:與正常組比較:*P<0.05;與模型組比較:△P<0.05,△△P<0.01
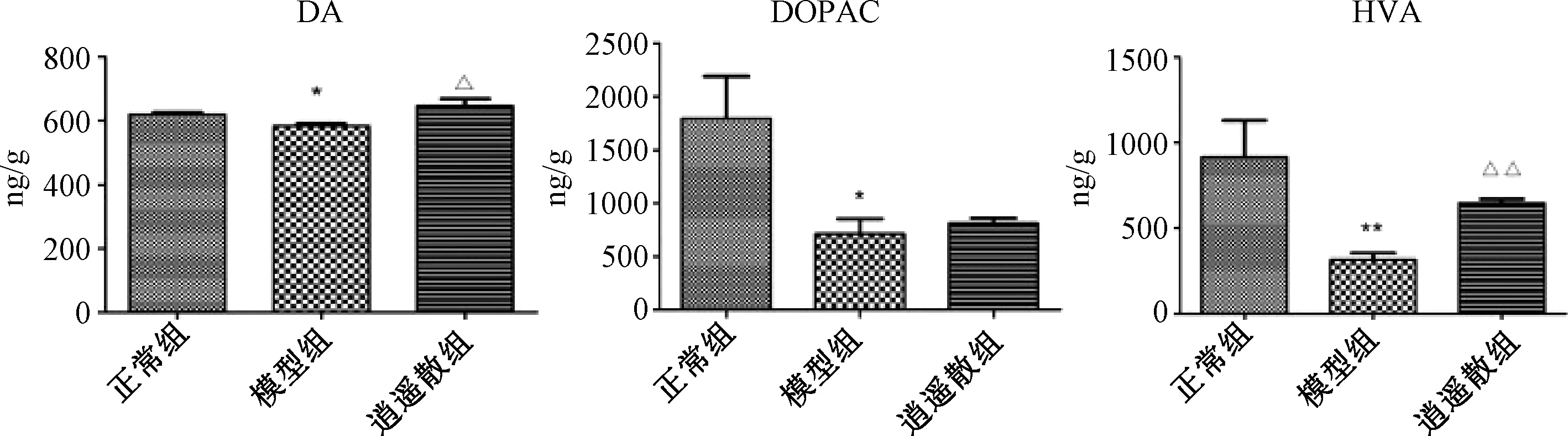
注:與正常組比較:*P<0.05,**P<0.01;與模型組比較:△P<0.05,△△P<0.01
2.4 各組大鼠前額皮質D1R、D2R、DAT的蛋白與mRNA表達
圖3示,前額皮質與正常組比較,模型組D1R的蛋白與mRNA表達均顯著上升(P<0.01),DAT的蛋白表達明顯增加(P<0.05),逍遙散對上述指標均有顯著下調作用(P<0.05,P<0.01)。模型組D2R的蛋白與mRNA表達較正常組均有明顯下降(P<0.05),逍遙散對其均表現出顯著的上調作用(P<0.05,P<0.01)。
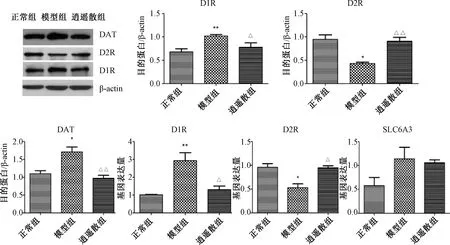
注:與正常組比較:*P<0.05,**P<0.01;與模型組比較:△P<0.05, △△P<0.01
2.5 各組大鼠伏核D1R、D2R、DAT的蛋白與mRNA表達
圖4示,伏核與正常組比較,模型組D1R蛋白與mRNA表達均顯著上升(P<0.01),DAT的蛋白與SLC6A3 mRNA表達也顯著上升(P<0.05),逍遙散對上述指標均有顯著下調作用(P<0.05,P<0.01)。模型組D2R的蛋白與mRNA表達較正常組均有明顯下降(P<0.05),逍遙散能夠顯著上調D2R蛋白表達(P<0.05)。
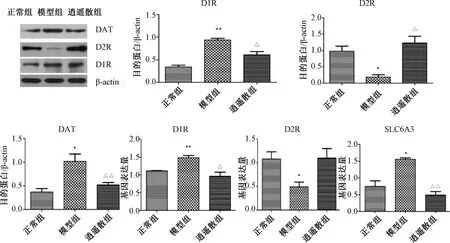
注:與正常組比較:*P<0.05,**P<0.01;與模型組比較:△P<0.05,△△P<0.01
3 討論
慢性不可預見應激模型在動物實驗中應用廣泛,可模擬人類抑郁癥發病機制[11]。曠場實驗、高架O迷宮是評價嚙齒類動物抑郁焦慮情緒的經典方法[12]。行為學結果顯示,慢性輕度不可預見應激模型大鼠在曠場實驗中活動性明顯降低,在中央區域的進入次數與停留時間也較正常對照組顯著下降。在O迷宮開放臂停留時間、進入次數及運動距離均有明顯降低,在封閉臂的停留時間明顯延長。分析結果可知,造模使大鼠對陌生環境的探索與適應能力下降、主動性與積極性降低,焦慮情緒增長,表現出明顯的抑郁樣癥狀。
單胺類神經遞質在中樞神經系統中起極其重要的作用[13]。DA是獎賞系統中主要神經遞質之一,DOPAC和HVA是DA的主要代謝產物,能夠反映中樞DA神經元的功能狀態[13]。本研究中,模型組大鼠在接受慢性應激刺激后,前額皮質、伏核腦區DA、DOPAC、HVA的含量均較正常組明顯下降。與國外1項研究發現,慢性應激能夠減少內側前額葉皮質中DA含量,與提高嚙齒類動物在社會擊敗中的易感性結果一致[14],說明慢性應激可能是通過下調腦內DA及其代謝產物水平,進而影響獎賞系統功能、導致快感減低等抑郁相關的癥狀表現。
神經受體假說認為,抑郁癥的發病與DA受體異常表達相關[15]。DA受體可分為D1樣和D2樣受體兩類。D1R屬于興奮型D1樣受體,主要定位于突觸后膜,激活后能夠誘發多個細胞內信號通路,增強神經元的興奮性,參與個體快感體驗能力的維持[16]。實驗證實,給小鼠注射D1R受體拮抗劑可以減少其運動動機[17]。D2R屬于D2樣受體,分布于DA能神經元突觸前膜,其所介導的信號通路作用與D1R正好相反,D2R和DA結合后可以通過激活G蛋白門控內向整流鉀離子通道,反過來使DA能神經元發生超極化,降低DA神經元興奮性,抑制DA的釋放[18]。伏核參與獎勵相關過程,激活伏核中阿片類受體和內源性大麻素受體,增強機體對食物獎勵的情感反應。但在快感缺失人群中,這種獎勵進程會被中斷[19]。前額皮質是除海馬以外最容易受到應激影響的腦區之一,慢性應激導致的前額皮質神經形態、突觸密度與功能的改變與多種情緒與認知功能失調有關[20]。本研究結果顯示,慢性輕度不可預見應激抑郁大鼠前額皮質和伏核腦區D1R mRNA與蛋白表達較正常組有所上升,D2R mRNA與蛋白表達下降。與抑郁癥自殺者尸檢發現,腦紋狀體組織中D1R含量增加結果一致[21],說明DR1和D2R均參與慢性輕度不可預見應激抑郁的發生過程,其相反的表達趨勢可能與二者在抑郁發病過程中所發揮的作用并不相同有關。本研究還發現,抑郁模型大鼠腦組織中DAT的基因與蛋白表達上升。DAT位于神經細胞突觸前膜,可調節細胞內DA存儲池的存儲數量,在維持多巴胺能神經傳遞的穩態方面有十分重要的作用。DAT受體增加會引發突觸間隙DA的耗竭,突觸后D1和D2受體表達代償性改變[25]。對轉基因小鼠的研究發現,DAT基因敲除小鼠細胞外DA濃度增加了5倍,而DAT基因加載小鼠紋狀體細胞外DA水平持續性降低,并顯示出突觸后D1R水平有顯著升高[26]。故推測本實驗中DAT和DA受體的變化都反映了應激后DA能系統處于非穩態狀態,進而導致個體對長期慢性應激表現出超敏反應,影響到動物的行為與情緒。
內科名方逍遙散出自《太平惠民和劑局方》,由柴胡、白芍、白術、茯苓、當歸、甘草、生姜、薄荷8味藥物組成,能疏肝解郁、養血健脾,是臨床常用調和肝脾的代表方,對抑郁癥有明確的治療作用[24-26],其抗抑郁的作用可由多種機制介導。本實驗中,逍遙散能顯著上調曠場中央區進入次數,上調O迷宮實驗中5 min平均速度、開放臂停留時間、開放臂運動距離、5 min運動總距離,減少封閉臂停留時間,使抑郁狀態改善。同時,逍遙散對模型大鼠各腦區的DA、DOPAC和HVA水平有不同程度上調作用,還能顯著降低模型大鼠前額皮質與伏核腦區中D1R mRNA及蛋白表達,升高前額皮質D2R mRNA及蛋白表達水平,對伏核腦區中DAT mRNA及蛋白表達也有下調作用。根據行為學及指標進行推測,逍遙散對慢性輕度不可預見應激抑郁大鼠的治療作用,可能與其調節模型大鼠前額皮質與伏核腦區DA及其代謝產物水平,DA受體D1R、D2R及DAT的表達,反饋性地調節DA受體平衡與穩定狀態相關,其干預機制可能包括逍遙散能夠通過阻斷突觸后DA受體,使DA能神經元反饋性興奮,釋放更多的DA神經遞質;逍遙散通過抑制突觸前膜DAT,降低對突觸間隙DA的重攝取與轉運,從而減少對DA的清除,使突觸間隙DA含量增加或DA消減時間延長,從而增強DA神經元介導的獎賞效應。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
光學精密工程(2016年6期)2016-11-07 09:07:19
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
核科學與工程(2015年4期)2015-09-26 11:59:03
