腸內外免疫營養對重癥急性胰腺炎大鼠機體免疫功能的影響
2021-12-30 09:22:32韓智君穆扎帕依米提迪米拉阿里根帕爾哈提阿不都熱衣木
中華災害救援醫學 2021年12期
韓智君,穆扎帕·依米提,迪米拉·阿里根,帕爾哈提·阿不都熱衣木
重癥急性胰腺炎(Severe Acute Pancreatitis,SAP)是指胰酶在胰腺內被激活后引起胰腺組織自身消化的急性化學性炎癥,具有起病迅急、病情重、并發癥多、病死率高等特點[1]。近年來營養支持作為一種非手術治療方法臨床應用較多[2],常見的營養支持方法包括腸內營養支持[3](Enteral Mutrition,EN)和腸外營養支持(Parenteral Nutrition,PN)。腸外營養是臨床常用的方法,刺激性小且符合SAP高代謝要求;腸內營養可刺激SAP患者腸黏膜細胞生長,促進腸道功能恢復,保持腸黏膜結構完整[4]。免疫營養是在普通營養基礎上添加免疫營養素后可以針對炎性反應、營養不良、代謝異常、免疫失衡等方面改善患者的營養、代謝和免疫狀態[5]。由此,本研究構建大鼠SAP模型,并給予適當的腸內、腸外免疫營養配方,對比腸內、腸外免疫營養治療對SAP大鼠腸屏障的作用,并初步解析其機制。
1 材料與方法
1.1 材料
1.1.1 實驗動物及分組 選取清潔級SD雄性大鼠,共48只,8~12 W,體重200~240 g,購自廣東省醫學實驗動物中心,合格證號:SCXK(粵)2013-0002。將大鼠隨機分為假手術組、模型組、普通腸內營養組、腸內免疫營養組、普通腸外營養組和腸外免疫營養組,每組8只。
1.1.2 實驗主要試劑與儀器 內毒素試劑盒、FITCConjugated Mouse anti-Rat CD4 monoclonal antibody、PEConjugated Mouse anti-Rat CD8 monoclonal antibody、大鼠分泌型免疫球蛋白(SIgA)、白細胞介素10(IL-10)、白細胞介素6(IL-6)、ELISA檢測試劑盒、流式細胞儀BD FACSCantoTMⅡ(Becton-Dickinson)、倒置熒光顯微鏡、低速離心機、實時熒光定量PCR儀、酶標儀。
1.2 建立動物模型、分組 根據參考文獻[6]建立大鼠SAP模型。假手術組:切開腹腔后,不注射牛磺膽酸鈉,輕翻胰腺后縫合切口。將建立模型的大鼠隨機分為模型組、普通腸內營養組、腸內免疫營養組、普通腸外營養組和腸外免疫營養組。模型組大鼠腹腔注射生理鹽水。普通腸內營養組:建模24 h內,經空腸勻速輸入平衡液,建模24 h后,給予瑞素。腸內免疫營養組:在瑞素中加入谷氨酰胺(24.15 g/kg)、精氨酸(11.63 g/kg)和ω-3脂肪酸(3 g/kg)。普通腸外營養組:建模24 h內,經頸內靜脈勻速輸入平衡液,建模24 h后,給予每100 ml含7%凡命、20% 英脫利匹特、50%葡萄糖溶液、7 ml力肽、0.4 ml注射用水溶性維生素(Soluvit)、0.4 ml多種微量元素注射液(Ⅱ)(Addamel)和0.4 ml脂溶性維生素注射液(Ⅱ)(Vitalipid)以及22 ml平衡液。腸外免疫營養組:在普通腸外營養的基礎上按1.5 ml/kg加入L-丙氨酰-L-谷氨酰胺(L-alanyl-L-glutamine, LALG)后立即輸入。連續給藥3 d。
1.3 酶聯免疫吸附測定(Enzyme Linked Immunosorbent Assay,ELISA)檢測各組大鼠小腸黏液SIgA含量及血漿IL-10、IL-6表達水平 處理結束后麻醉處死大鼠并采用腹主動脈取血5 ml,將血液置于肝素抗凝管中,2 500 r/min離心15 min,取血漿,采用ELISA檢測各組大鼠血漿IL-10、IL-6的表達水平。剪取空回腸縱行剖開,用玻片刮取大鼠腸黏液,加入l ml濃度為0.01 mol/L的PBS緩沖液,30 000 r/min離心10 min,取上清,采用ELISA檢測大鼠腸黏液SIgA含量。
1.4 HE染色檢查胰腺組織病理學改變 麻醉處死大鼠取胰腺,用生理鹽水沖洗,15%中性甲醛溶液固定、石蠟包埋、切片(4 μm),蘇木素染色液染色3~10 min,光鏡下觀察胰腺組織的病理改變。
1.5 各組大鼠血漿內毒素檢測 采用無熱原肝素鈉采血管采集腹主動脈血5 ml,2 500 r/min離心15 min,取血漿,按照內毒素試劑盒說明書進行。
1.6 流式細胞術檢測外周血CD4+T、CD8+T、CD4+T/CD8+T淋巴細胞 在專用試管中加入大鼠外周血標本100 μl,分別加入異硫氰酸熒光素(Fluorescein Isothiocyanate,FITC)標記的CD4單克隆抗體和FITC標記的CD8單克隆抗體5 μl,然后充分混勻,室溫下避光孵育15 min,然后每管加入紅細胞裂解液2 ml充分混勻,室溫避光下溶血10 min,1 500 r/min離心10 min,棄上清,PBS洗滌后采用流式細胞術進行檢測。
1.7 實時熒光定量PCR檢測各組大鼠occludin mRNA表達水平 取小腸組織約2 g,加入TRIZOL試劑后進行勻漿。逆轉錄成cDNA,取2 μl cDNA為模板,配制25 μl Realtime PCR反應體系,具體反應體系參照試劑盒說明書,95℃預變性15 min,95℃變性10 s,60℃退火延伸30 s,擴增40個循環。甘油醛-3-磷酸脫氫酶(Glyceraldehyde-3-phosphate Dehydrogenase,GAPDH) 為 內 參, 采 用2-△△Ct法 計 算。occludin引物為:上游5’-ACGGTGCCATAGAATGAGATGTTG-3',下 游5’-CAGCTAGTTGTTCATTTCTGCACCA-3';GAPDH引 物 為: 上 游5’-TCCCTCAAGATTGTCAGCAA-3’, 下 游5’-AGATCCACAACGGATACATT-3’。
1.8 統計學處理 采用SPSS 20.0統計軟件進行數據分析,結果用表示,采用單因素方差分析對多樣本實驗結果進行統計分析,進一步兩兩比較采用最小顯著性差異法(Least-Significant Difference,LSD-t)檢驗。以P<0.05為差異有統計學意義。
2 結 果
2.1 各組大鼠小腸黏液SIgA含量及血漿IL-6、IL-10表達水平 與假手術組相比,模型組大鼠IL-6表達水平均顯著升高而小腸黏液SIgA含量、IL-10表達水平降低(P<0.05);與模型組相比,各治療組大鼠IL-6表達水平均降低而SIgA含量、IL-10表達水平升高(P均<0.05);與普通腸內營養組和普通腸外營養組相比,腸內免疫營養組和腸外免疫營養組IL-6表達水平均有不同程度降低而SIgA含量、IL-10表達水平均升高(P均<0.05);與腸外免疫營養組相比,腸內免疫營養組IL-6表達水平更低而SIgA含量、IL-10表達水平更高(P<0.05)。見表1。
表1 各組大鼠 SIgA含量及血漿IL-6、IL-10表達水平比較(±s,n=48)
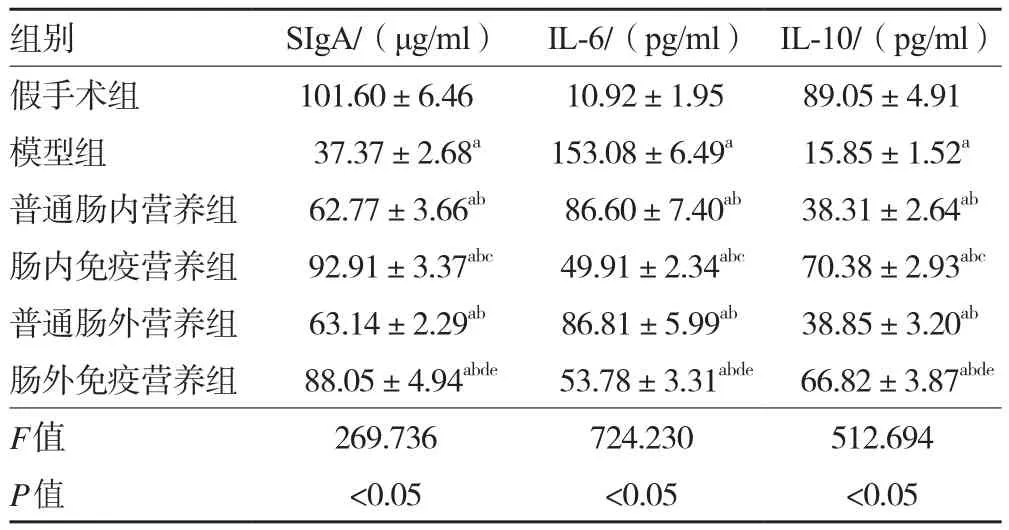
表1 各組大鼠 SIgA含量及血漿IL-6、IL-10表達水平比較(±s,n=48)
注:與假手術組比較,aP<0.05;與模型組比較,bP<0.05;與普通腸內營養組比較,cP<0.05;與普通腸外營養組比較,dP<0.05;與腸內免疫營養組比較,eP<0.05
組別 SIgA/(μg/ml) IL-6/(pg/ml) IL-10/(pg/ml)假手術組 101.60±6.46 10.92±1.95 89.05±4.91模型組 37.37±2.68a 153.08±6.49a 15.85±1.52a普通腸內營養組 62.77±3.66ab 86.60±7.40ab 38.31±2.64ab腸內免疫營養組 92.91±3.37abc 49.91±2.34abc 70.38±2.93abc普通腸外營養組 63.14±2.29ab 86.81±5.99ab 38.85±3.20ab腸外免疫營養組 88.05±4.94abde 53.78±3.31abde 66.82±3.87abde F值 269.736 724.230 512.694 P值 <0.05 <0.05 <0.05
2.2 各組大鼠胰腺組織病理形態學變化 假手術組大鼠胰腺細胞結構正常,包膜完整。模型組大鼠出現廣泛凝固性壞死,間質水腫明顯,大量單核和中性粒細胞浸潤。普通腸內營養組和普通腸外營養組大鼠胰腺組織結構與模型組相比,較完整,壞死組織較少,中性粒細胞浸潤減少,病變程度相對減輕;腸內免疫營養組和腸外免疫營養組大鼠胰腺組織結構較普通腸內營養組和普通腸外營養組完整,壞死組織減少,中性粒細胞浸潤減少,且腸內免疫營養組大鼠胰腺組織結構較腸外免疫營養組大鼠更完整。見圖1。

圖1 各組大鼠胰腺組織病理形態學變化(HE染色,×100)
2.3 各組大鼠血漿內毒素含量 與假手術組相比,模型組大鼠血漿內毒素含量顯著升高(P<0.05);與模型組相比,各治療組大鼠內毒素含量均降低(P均<0.05);與普通腸內營養組和普通腸外營養組相比,腸內免疫營養組和腸外免疫營養組內毒素含量均有不同程度降低(P均<0.05)。見表2。
表2 各組大鼠血漿內毒素含量(±s,n=48)
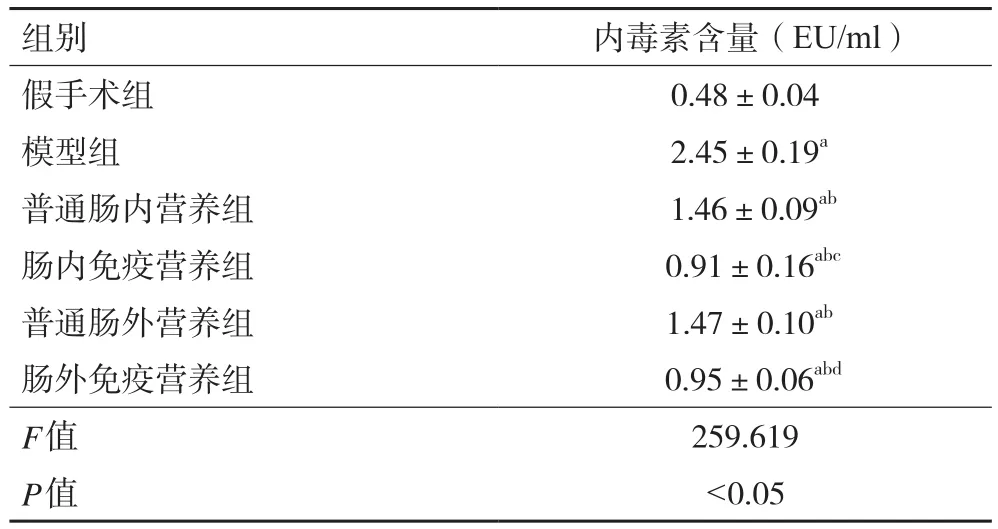
表2 各組大鼠血漿內毒素含量(±s,n=48)
注:與假手術組比較,aP<0.05;與模型組比較,bP<0.05;與普通腸內營養組比較,cP<0.05;與普通腸外營養組比較,dP<0.05
組別 內毒素含量(EU/ml)假手術組 0.48±0.04模型組 2.45±0.19a普通腸內營養組 1.46±0.09ab腸內免疫營養組 0.91±0.16abc普通腸外營養組 1.47±0.10ab腸外免疫營養組 0.95±0.06abd F值 259.619 P值 <0.05
2.4 各組大鼠外周血CD4+T、CD8+T、CD4+T/CD8+T淋巴細胞 與假手術組相比,模型組大鼠外周血CD4+T、CD4+T/CD8+T淋巴細胞均顯著降低(P<0.05);與模型組相比,各治療組大鼠CD4+T、CD4+T/CD8+T淋巴細胞均升高(P均<0.05);與普通腸內營養組和普通腸外營養組相比,腸內免疫營養組和腸外免疫營養組CD4+T、CD4+T/CD8+T淋巴細胞均有不同程度升高(P均<0.05);而各組CD8+T淋巴細胞差異無統計學意義(P>0.05)。見表3、圖2。
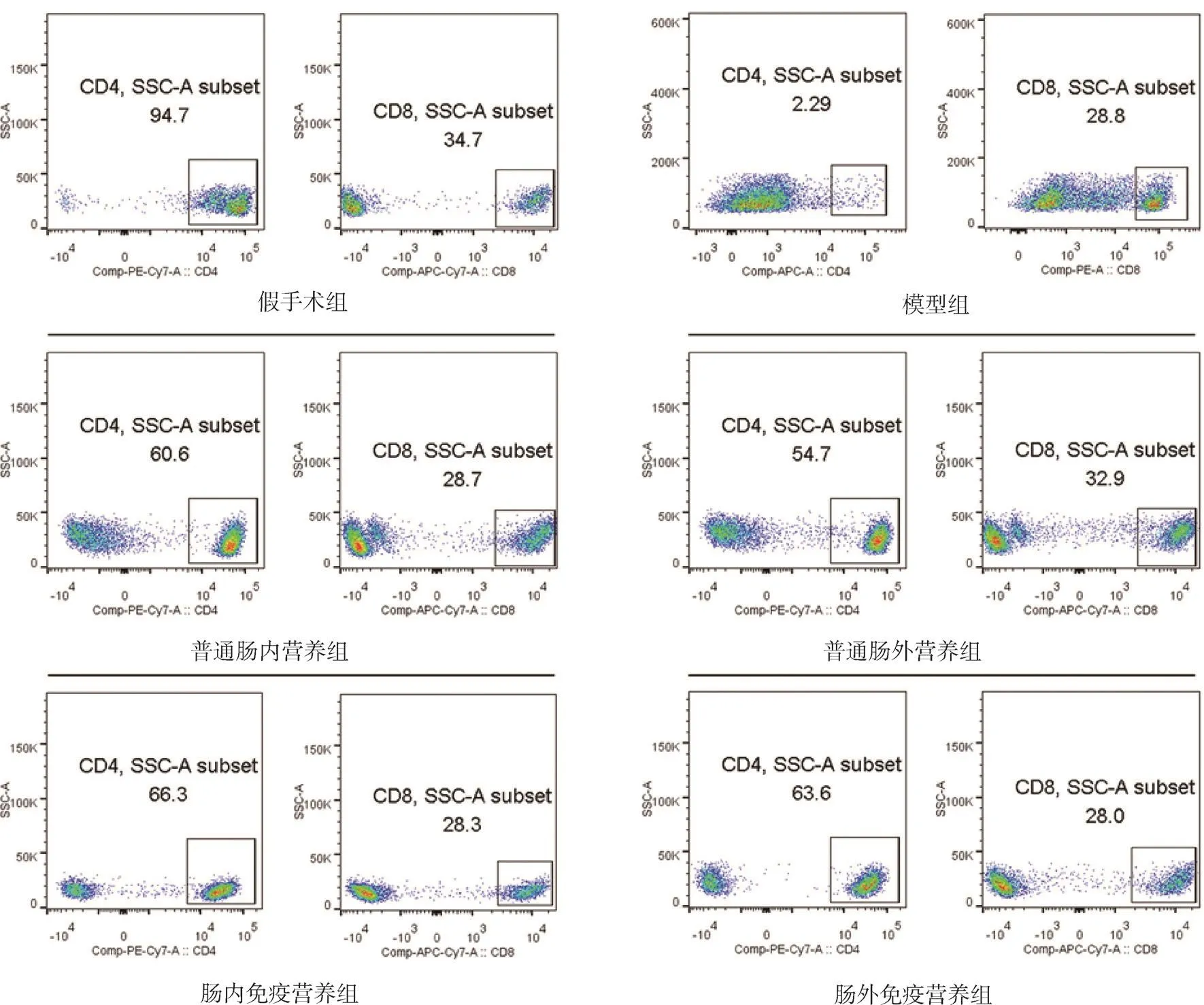
圖2 各組大鼠外周血CD4+T、CD8+T淋巴細胞
表3 各組大鼠外周血CD4+T、CD8+T、CD4+T/CD8+T淋巴細胞比較(±s,n=48)
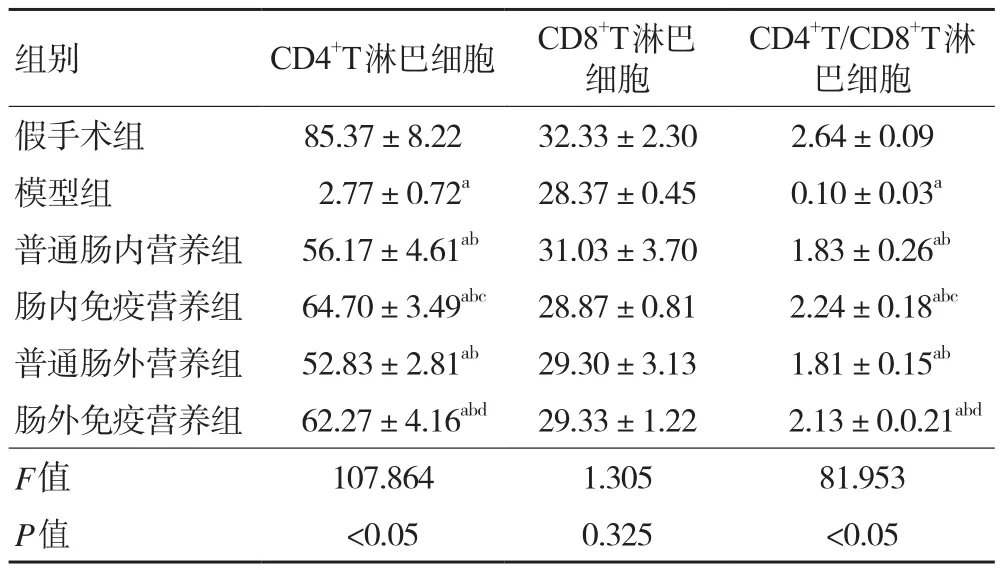
表3 各組大鼠外周血CD4+T、CD8+T、CD4+T/CD8+T淋巴細胞比較(±s,n=48)
注:與假手術組比較,aP<0.05;與模型組比較,bP<0.05;與普通腸內營養組比較,cP<0.05;與普通腸外營養組比較,dP<0.05
組別 CD4+T淋巴細胞 CD8+T淋巴細胞CD4+T/CD8+T淋巴細胞假手術組 85.37±8.22 32.33±2.30 2.64±0.09模型組 2.77±0.72a 28.37±0.45 0.10±0.03a普通腸內營養組 56.17±4.61ab 31.03±3.70 1.83±0.26ab腸內免疫營養組 64.70±3.49abc 28.87±0.81 2.24±0.18abc普通腸外營養組 52.83±2.81ab 29.30±3.13 1.81±0.15ab腸外免疫營養組 62.27±4.16abd 29.33±1.22 2.13±0.0.21abd F值 107.864 1.305 81.953 P值 <0.05 0.325 <0.05
2.5 各組大鼠小腸粘黏膜occludin mRNA表達水平 與假手術組相比,模型組大鼠小腸黏膜occludin mRNA表達水平顯著降低(P<0.05);與模型組相比,各治療組大鼠occludin mRNA表達水平升高(P<0.05);與普通腸內營養組和普通腸外營養組相比,腸內免疫營養組和腸外免疫營養組occludin mRNA表達水平均有不同程度升高(P<0.05)。見表4。
表4 各組大鼠小腸粘膜occludin mRNA半定量表達水平(±s,n=48)
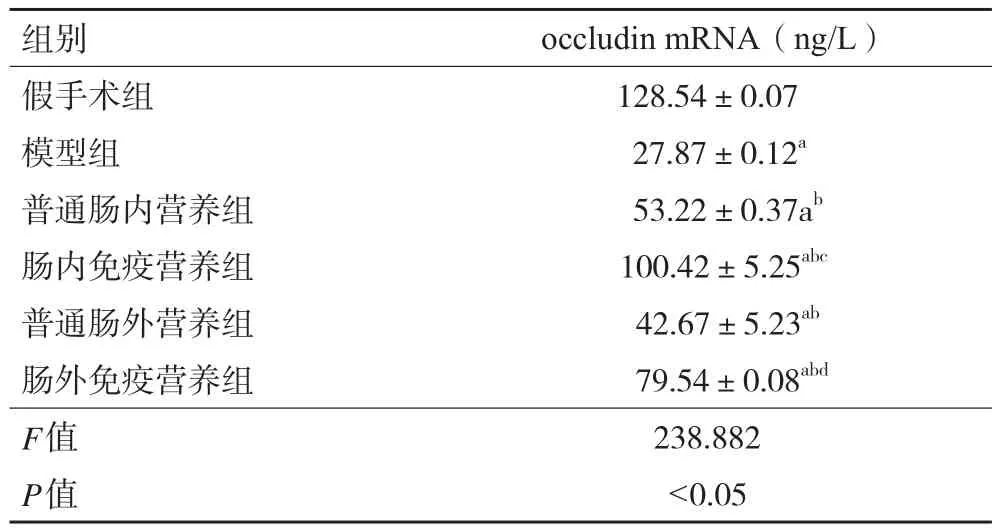
表4 各組大鼠小腸粘膜occludin mRNA半定量表達水平(±s,n=48)
注:與假手術組比較,aP<0.05;與模型組比較,bP<0.05;與普通腸內營養組比較,cP<0.05;與普通腸外營養組比較,dP<0.05
組別 occludin mRNA(ng/L)假手術組 128.54±0.07模型組 27.87±0.12a普通腸內營養組 53.22±0.37ab腸內免疫營養組 100.42±5.25abc普通腸外營養組 42.67±5.23ab腸外免疫營養組 79.54±0.08abd F值 238.882 P值 <0.05
3 討 論
腸道是機體最大的細菌存儲所和內毒素中心,SAP在急性反應期免疫過激,會引起腸屏障的損傷,導致大量腸源性內毒素釋放入血并激活體內炎癥因子,從而促進全身炎癥反應引起多器官衰竭,免疫異常在SAP病情的發展中起重要作用[7-8]。有研究表明,腸屏障功能的損傷所引發內毒素及細菌移位被認為是胰腺繼發感染的重要原因[9]。腸道免疫屏障主要依靠腸黏膜表面黏液及腸腔中的免疫球蛋白(以SIgA為主)和淋巴細胞,共同完成腸道的局部免疫防御功能[10]。有研究發現,SAP大鼠腸黏膜組織中產生了嚴重的病理損傷,血清和腸組織中炎癥因子水平升高提示系統性及腸道出現局部炎癥環境[11]。另有研究發現,在SAP發生進展中,由于循環血液中腸源性內毒素水平升高,導致NF-κB、IL-1β、IL-6、IL-8等多種細胞炎性因子分泌增加[12-13]。本研究建立SAP大鼠模型,結果發現與假手術組相比,模型組大鼠血漿內毒素含量、IL-6表達水平均顯著升高而小腸黏液SIgA含量、IL-10表達水平降低;光鏡結果顯示與假手術組相比,模型組大鼠出現廣泛凝固性壞死,間質水腫明顯,大量單核和中性粒細胞浸潤。此外,流式細胞術結果發現,模型組大鼠外周血CD4+T、CD4+T/CD8+T淋巴細胞均顯著降低。提示SAP大鼠模型建立成功,大鼠免疫功能被破壞,體內炎癥水平及內毒素升高,導致腸組織損傷,促進SAP發展。
緊密連接蛋白是腸黏膜機械屏障重要組成部分,occludin作為腸上皮重要的緊密連接蛋白,主要參與腸壁通透性的調節,對于維護黏膜屏障的完整性有非常重要的意義[14-15]。有研究發現,occludin蛋白表達的變化是引起細菌移位、維持腸黏膜屏障功能的關鍵[16]。本研究結果發現,與假手術相比,模型組大鼠小腸黏膜occludin mRNA表達水平顯著降低;與模型組相比,各治療組occludin mRNA表達水平明顯升高。提示SAP大鼠小腸黏膜occludin表達降低會導致腸道通透性增高,從而導致內毒素等從腸腔中進入血液。
目前臨床對于SAP的治療主要采取營養支持治療作為重要的輔助手段。有研究發現,將腸內免疫微生態營養應用于重癥急性胰腺炎,有利于維持抗炎反應的平衡,實現對病情及預后的改善[17]。另有研究表明,谷氨酰胺增強的腸外營養能顯著改善SAP大鼠機體免疫、降低血漿內毒素,有利于細胞與器官功能的恢復[18]。本研究采用腸外免疫營養和腸內免疫營養進行治療并進行比較,以分析腸外和腸內營養對SAP大鼠腸組織免疫功能的影響。結果發現,與普通腸內營養組和普通腸外營養組相比,腸內免疫營養組和腸外免疫營養組內毒素含量、IL-6表達水平均有不同程度降低而SIgA含量、IL-10表達水平、CD4+T、CD4+T/CD8+T淋巴細胞及occludin mRNA表達均升高;與腸外免疫營養組相比,腸內免疫營養組大鼠各項指標改善更為顯著。提示腸內外免疫營養支持均可顯著改善SAP。
綜上所述,SAP大鼠小腸黏膜occludin表達降低導致內毒素等從腸腔中進入血液,破壞大鼠免疫功能,體內炎癥水平升高,導致腸組織損傷;腸內外免疫營養支持均可顯著改善SAP,但腸內免疫營養支持在提升SAP大鼠免疫機能和改善免疫狀態方面效果優于腸外營養。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
今日農業(2021年11期)2021-08-13 08:53:34
河北畫報(2020年10期)2020-11-26 07:20:50
人大建設(2019年12期)2019-05-21 02:55:32
中國生殖健康(2019年8期)2019-01-07 01:18:24
幸福(2018年33期)2018-12-05 05:22:46
海峽姐妹(2016年5期)2016-02-27 15:20:20
中國衛生標準管理(2015年3期)2016-01-14 03:41:47
中國中醫藥現代遠程教育(2014年13期)2014-03-01 04:26:36
共產黨員(遼寧)(2011年23期)2011-10-18 09:29:10
