南亞熱帶馬尾松人工林近自然恢復過程中林下木本植物生態位動態*
2022-01-07 09:16:12王永琪馬姜明黃柳欣莫燕華李明金
廣西科學 2021年5期
黎 露,王永琪,馬姜明**,張 惠,黃柳欣,莫燕華,李明金
(1.廣西師范大學可持續發展創新研究院,廣西桂林 541006;2.廣西師范大學生命科學學院,廣西桂林 541006;3.廣西優良用材林資源培育重點實驗室,廣西南寧 530002;4.橫州鎮龍林場,廣西南寧 5300327)
0 引言
生態位是指物種在特定環境中的功能地位,包括環境對物種的影響以及物種對環境的響應規律[1],并可作為定量探索物種在特定環境中的資源利用及競爭能力的指標[2],也是解釋群落中物種共存與競爭機制的理論之一。生態位的主要特征是生態位寬度和生態位重疊,對前者的研究有助于了解物種在群落中的適應和資源利用能力,后者用于說明群落中物種利用資源的狀態[3]。近幾十年來,生態位理論被廣泛應用于生物多樣性[4]、群落結構、種間關系[5-7]和群落演替[8]等方面的研究。林下物種是森林群落的重要組成部分,在保護物種多樣性、維護生態系統穩定等方面發揮著重要的作用。評估各演替階段林下木本植物的種間關系及種群在群落中的功能,有助于人工林物種多樣性的增加和生態功能的恢復。
馬尾松Pinusmassoniana是亞熱帶常綠闊葉林遭到破壞后最早出現的先鋒樹種之一[9],適應性強,在我國南方廣泛種植,產生較大的經濟、生態效益。目前,關于馬尾松人工林生態位的研究較多,主要集中在研究馬尾松群落某一階段主要種群的生態位[10]和種間聯結[11],但對不同林齡馬尾松人工林林下物種生態位的動態變化研究較少并多為現象描述[12]。因此,本文通過對廣西南亞熱帶不同林齡馬尾松人工林林下主要木本植物的生態位研究,揭示不同恢復階段馬尾松人工林林下木本植物資源利用和種間關系的動態變化,為加速人工林近自然演替進程提供重要理論依據。
1 材料與方法
1.1 研究區概況
研究樣地位于廣西橫州市北部鎮龍林場,地理坐標為109°08′-109°19′ E,23°02′-23°08′ N,海拔高度為400-700 m的低山丘陵。年平均氣溫為21.5℃,極端低溫-1℃,極端高溫39.2℃;屬于南亞熱帶季風氣候;年平均降水量為1 477.8 mm,降水量充沛;年平均日照時長為1 758.9 h,日照充足;土壤多為赤紅壤,呈酸性或微酸性[13]。
1.2 樣地設置與群落調查
2018年7-9月,在廣西橫州市鎮龍林場選擇立地條件大致相似的中齡林(1999年植)、成熟林(1986年植)和過熟林(1960年植)3種馬尾松人工林類型。3種馬尾松人工林的初植密度均為2 500株/hm2,營林措施為種植第1-3年進行除雜草保育措施,第8-9年進行衛生伐撫育措施之后不再進行人為干預,馬尾松人工林進入近自然恢復狀態。每種人工林類型在立地條件相似,土壤質地、土層厚度和林木長勢較為一致的地段分別設置3個20 m×20 m的標準樣方,并將每個標準樣方劃分為4個10 m×10 m的小樣方。對樣方中除馬尾松外的胸徑≥1 cm的木本植物進行每木檢尺、坐標定位,其中高度≥5 m劃分為亞喬木層,高度<5 m劃分為灌木層。記錄各樣方內木本植物種名、樹高、胸徑、基徑和株數等信息,樣方基本情況見表1。
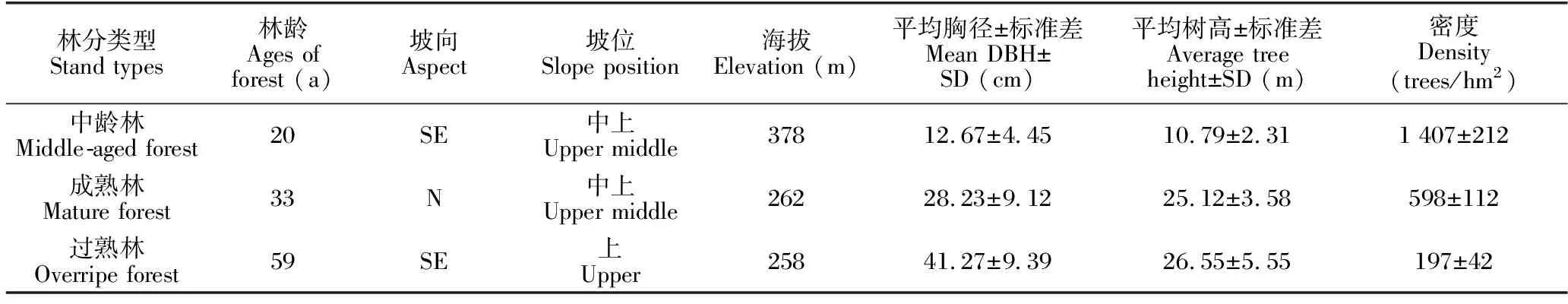
表1 樣方基本情況
1.3 數據處理與分析
1.3.1 重要值
利用物種的相對多度、相對顯著度和相對頻度計算喬木重要值,灌木重要值利用相對多度、相對蓋度和相對頻度進行計算。重要值表示群落中不同物種的相對重要程度,能夠反映不同物種在群落中的地位和重要性,重要值越大說明在群落中越有優勢[8],計算公式如下:
亞喬木層物種重要值=(相對多度+相對顯著度+相對頻度)/3×100,
灌木層物種重要值=(相對多度+相對蓋度+相對頻度)/3×100。
1.3.2 生態位特征分析
分別選取馬尾松中齡林、成熟林和過熟林3個階段的亞喬木層和灌木層中重要值排前10和前20的主要物種進行生態位寬度和生態位重疊的分析,生態位寬度采用Levins指數(Bi)[14]進行計算分析,生態位重疊采用Pianka重疊指數(Oik)[15]進行計算分析。
(1)Levins生態位寬度指數(Bi)
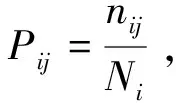
(2)Pianka生態位重疊指數(Oik)
式中,Oik為物種i和物種k的生態位重疊指數,Pij和Pkj分別表示物種i和物種k在第j個樣方中的重要值,r為樣方的總數。當Oik=1時,表明兩個物種間所利用的資源狀態完全相同,生態位完全重疊;當Oik=0時,表明兩個物種間沒有利用相同資源,生態位不重疊。
利用Excel 2019計算各群落中的物種重要值,R4.0.4軟件種間聯結程序分析包spaa中的niche.width()、niche.overlap()函數計算物種生態位寬度指數和生態位重疊指數。
2 結果與分析
2.1 不同恢復階段主要木本植物重要值
隨著馬尾松人工林從中齡林到成熟林和過熟林近自然恢復的進行,林下主要木本植物重要值存在明顯差異,如表2所示。馬尾松中齡林亞喬木層物種重要值排序為山烏桕(58.87)>鵝掌柴(13.07)>廣西水錦樹(5.84)>長毛柃(5.10)>楓香(4.91)>烏桕(3.71)>牛耳楓(2.76)>樟(2.52)>狹基潤楠(2.19)>黃毛榕(0.51);成熟林亞喬木層物種重要值排序為狹基潤楠(25.38)>廣西水錦樹(13.11)>楓香(10.87)>黃毛榕(9.11)>毛桐(4.59)>黃連木(4.58)>黃牛木(3.99)>南酸棗(3.96)>山烏桕(3.34)>木油桐(3.09);過熟林亞喬木層物種重要值排序為海南冬青(19.79)>鵝掌柴(18.15>大葉櫟(11.78)>廣西石楠(7.55)>狹基潤楠(6.88)>變葉榕(4.17)>橄欖(3.68)>石斑木(3.37)>石楠(3.26)>芳槁潤楠(2.81)。如表3所示,馬尾松中齡林林下灌木層重要值排前5的物種分別為鵝掌柴(9.34)、三椏苦(9.33)、草珊瑚(6.06)、鼠刺(5.91)、鯽魚膽(5.04),重要值排后3位的是山烏桕(2.06)、短柱柃(2.05)、藤黃檀(1.97);成熟林林下灌木層物種重要值最大的前5種分別是三椏苦(16.22)、狹基潤楠(11.45)、廣西水錦樹(5.90)、鵝掌柴(4.69)、黔桂冬青(4.08),重要值排后3位的物種是鯽魚膽(1.58)、草珊瑚(1.45)、臺灣榕(1.43);過熟林林下灌木層重要值排前5的物種為紐子果(23.81)、海南冬青(8.77)、九節(8.39)、大葉櫟(7.08)、三椏苦(6.52),重要值排后3位的是廣西石楠(1.24)、鼠刺(1.20)、香楠(1.16)。

表2 馬尾松人工林不同恢復階段亞喬木層物種重要值和生態位寬度指數

續表2

表3 馬尾松人工林不同恢復階段林下灌木層物種重要值和生態位寬度指數
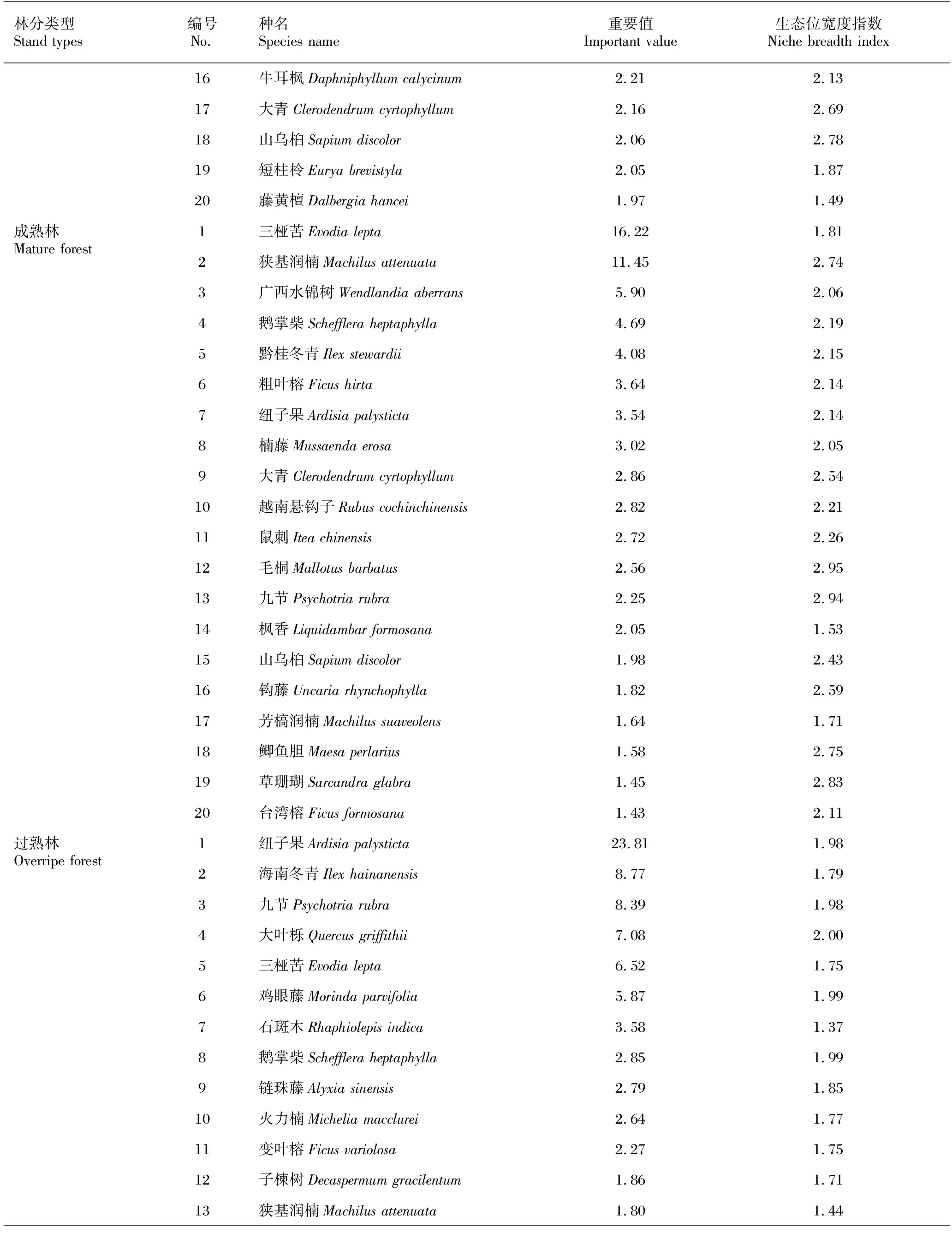
續表3

續表3
2.2 不同恢復階段主要木本植物生態位寬度
隨著馬尾松人工林從中齡林到成熟林和過熟林近自然恢復的進行,林下主要木本植物生態位寬度指數有所不同,如表2所示。在所選擇的重要值排前10的亞喬木層木本植物中,中齡林生態位寬度指數最大的是山烏桕(3.00),其次是鵝掌柴(2.97)和廣西水錦樹(2.89)等,生態位寬度指數最小的是黃毛榕(1.00);成熟林生態位寬度指數最大的是狹基潤楠(2.86),其次是廣西水錦樹(2.81)、楓香(2.00)和黃毛榕(2.00)等,生態位寬度指數最小的是南酸棗(1.00)、山烏柏(1.00)和木油桐(1.00);過熟林生態位寬度指數最大的是鵝掌柴(2.83),其次是狹基潤楠(2.56)和廣西石楠(2.53)等,生態位寬度指數最小的是變葉榕(1.00)、橄欖(1.00)、石楠(1.00)和芳槁潤楠(1.00)。如表3所示,在重要值排前20的灌木層木本植物中,中齡林生態位寬度指數最大的是三椏苦(3.00),其次是火力楠(2.99)和白花酸藤果(2.86)等,生態位寬度指數最小的是山雞椒(1.12);成熟林生態位寬度指數最大的是毛桐(2.95),其次是九節(2.94)和草珊瑚(2.83)等,生態位寬度指數最小的是楓香(1.53);過熟林生態位寬度指數最大的是大葉櫟(2.00),其次是雞眼藤(1.99)、鵝掌柴(1.99)和瓜馥木(1.99)等,生態位寬度指數最小的是石斑木(1.37)。
2.3 不同恢復階段亞喬木層物種生態位重疊
隨著馬尾松人工林從中齡林到成熟林和過熟林近自然恢復的進行,不同林齡亞喬木層呈現出不同的生態位重疊程度,如表4所示。中齡林亞喬木層45個種對中,生態位Pianka重疊指數(Oik)平均值為0.74;Oik≤0.5的有8對,占總種對數的17.78%,其中烏桕-狹基潤楠種對Oik值最小(0.22);Oik>0.5的有37對,占總種對數的83.22%;完全重疊(Oik=2)的種對有2對,占總種對數的4.44%,分別為山烏桕-鵝掌柴、牛耳楓-樟。成熟林亞喬木層的Oik平均值為0.53;Oik≤0.5的有20對,占總種對數的44.44%;Oik>0.5的有25對,占總種對數的55.56%;不發生重疊(Oik=0)的種對有10對,占總種對數的22.22%,分別為黃毛榕-木油桐、毛桐-黃連木、毛桐-南酸棗、毛桐-山烏桕、黃連木-黃牛木、黃連木-木油桐、黃牛木-南酸棗、黃牛木-山烏桕、南酸棗-木油桐、山烏桕-木油桐;完全重疊(Oik=1)的種對有3對,占總種對數的6.67%,分別為黃連木-南酸棗、黃連木-山烏桕、南酸棗-山烏桕。過熟林亞喬木層的Oik平均值為0.50;Oik<0.5的有22對,占總種對數的48.89%;Oik>0.5的種對有23對,占總種對數的51.11%;不發生重疊(Oik=0)的種對有7對,占總種對數的15.56%,分別為大葉櫟-石楠、變葉榕-石楠、變葉榕-芳槁潤楠、橄欖-石楠、橄欖-芳槁潤楠、石斑木-石楠、石楠-芳槁潤楠;完全重疊(Oik=1)的種對有1對,占總種對數的2.22%,為變葉榕-橄欖。
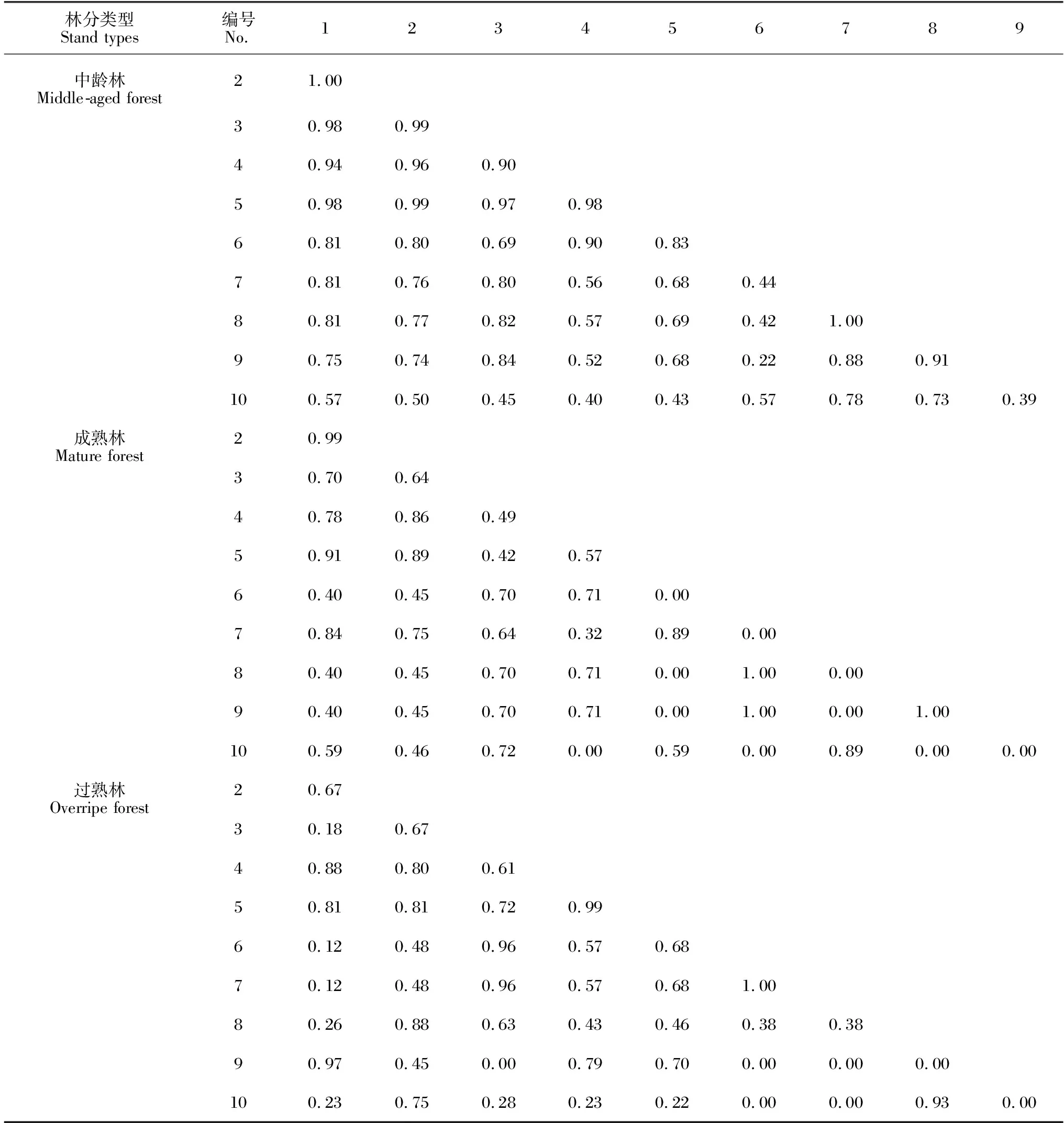
表4 不同恢復階段馬尾松人工林亞喬木層生態位重疊指數
2.4 不同恢復階段灌木層物種生態位重疊
如表5所示,隨著馬尾松人工林從中齡林到成熟林和過熟林近自然恢復的進行,不同林齡林下灌木層物種重疊程度不同。中齡林林下灌木層的20種植物共組成190個種對,該群落中的生態位重疊指數(Oik)平均值為0.74;Oik≤0.5的種對有31對,占總種對數的16.32%;Oik>0.5的種對有159對,占總種對數的83.68%;完全重疊(Oik=1)的種對有2對,占總種對數的1.05%,分別為三椏苦-火力楠、長毛柃-藤黃檀。如表6所示,成熟林林下灌木層物種Oik平均值為0.76;Oik≤0.5的種對有24對,占總種對數的12.63%;Oik>0.5的種對有166對,占總種對數的87.37%;完全重疊(Oik=1)的種對有8對,分別為狹基潤楠-鯽魚膽、鵝掌柴-鼠刺、黔桂冬青-紐子果、黔桂冬青-鼠刺、粗葉榕-楠藤、紐子果-越南懸鉤子、大青-山烏桕、鼠刺-山烏桕。由表7可知,過熟林林下灌木層物種的Oik平均值為0.89,該群落中20個物種均發生生態位重疊,其中,Oik>0.5的種對有190對,占總種對數的100%;完全重疊(Oik=1)的種對有39對,占總種對數的20.53%。
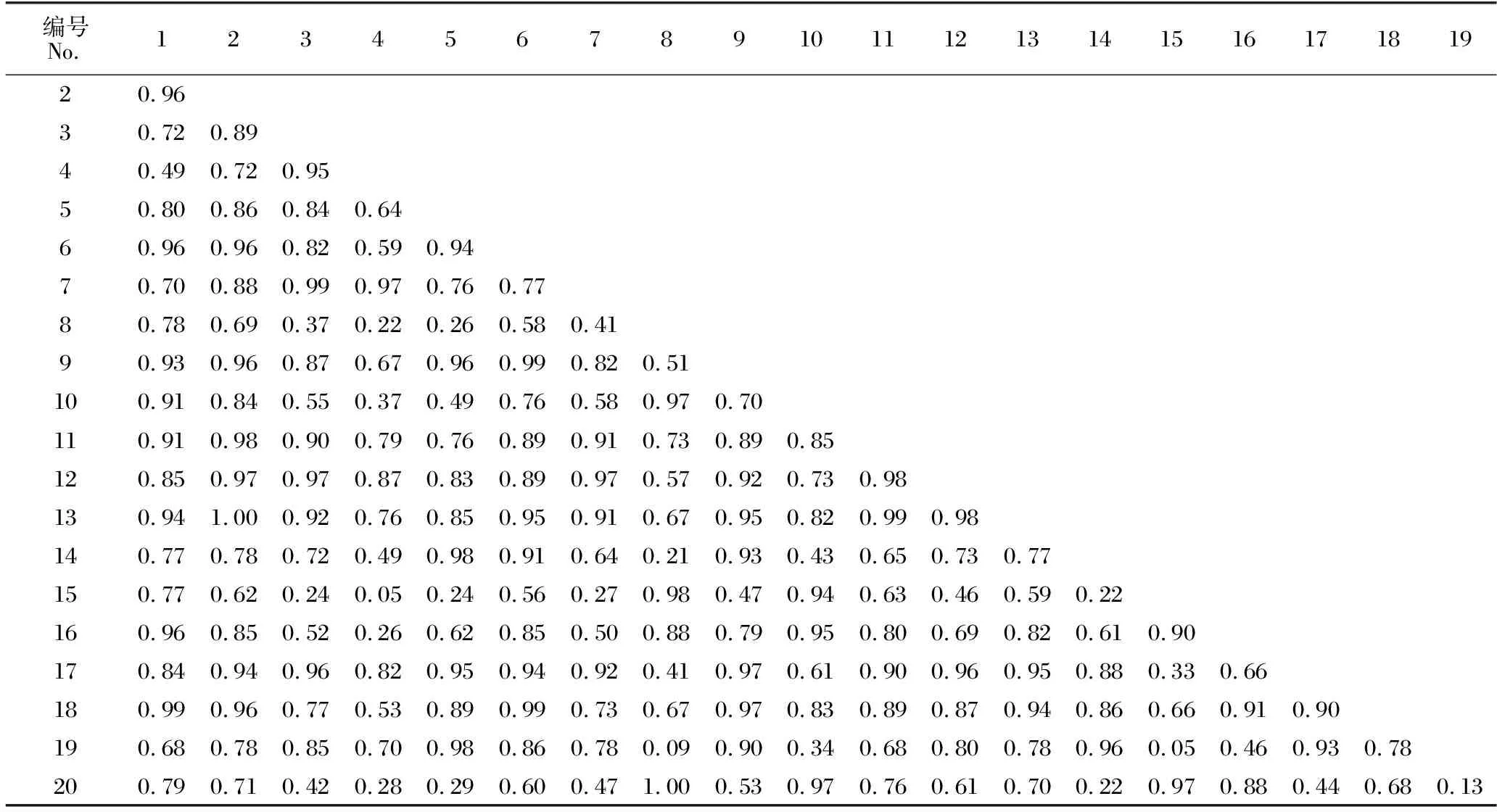
表5 中齡林林下灌木層物種生態位重疊指數
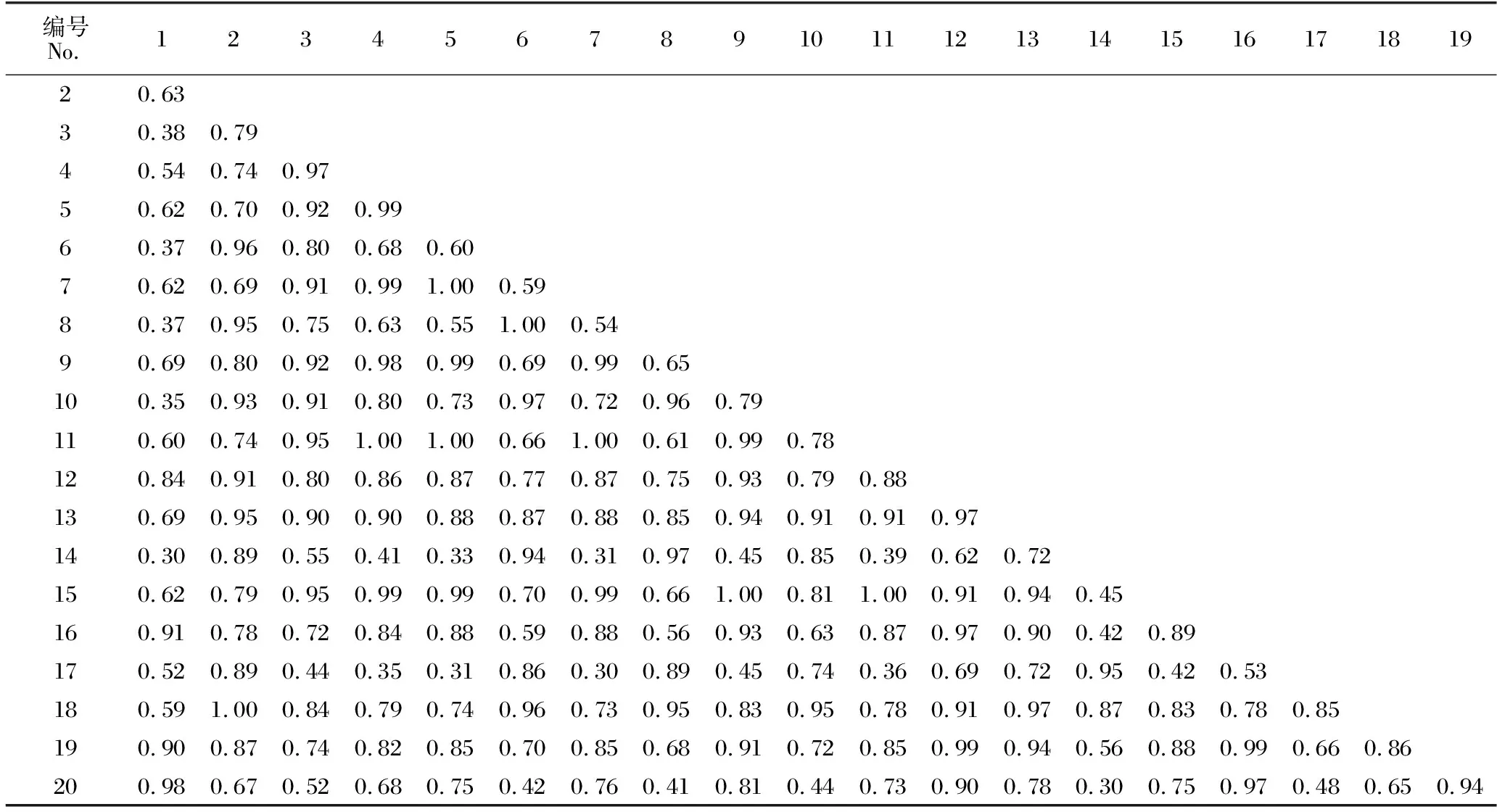
表6 成熟林林下灌木層物種生態位重疊指數
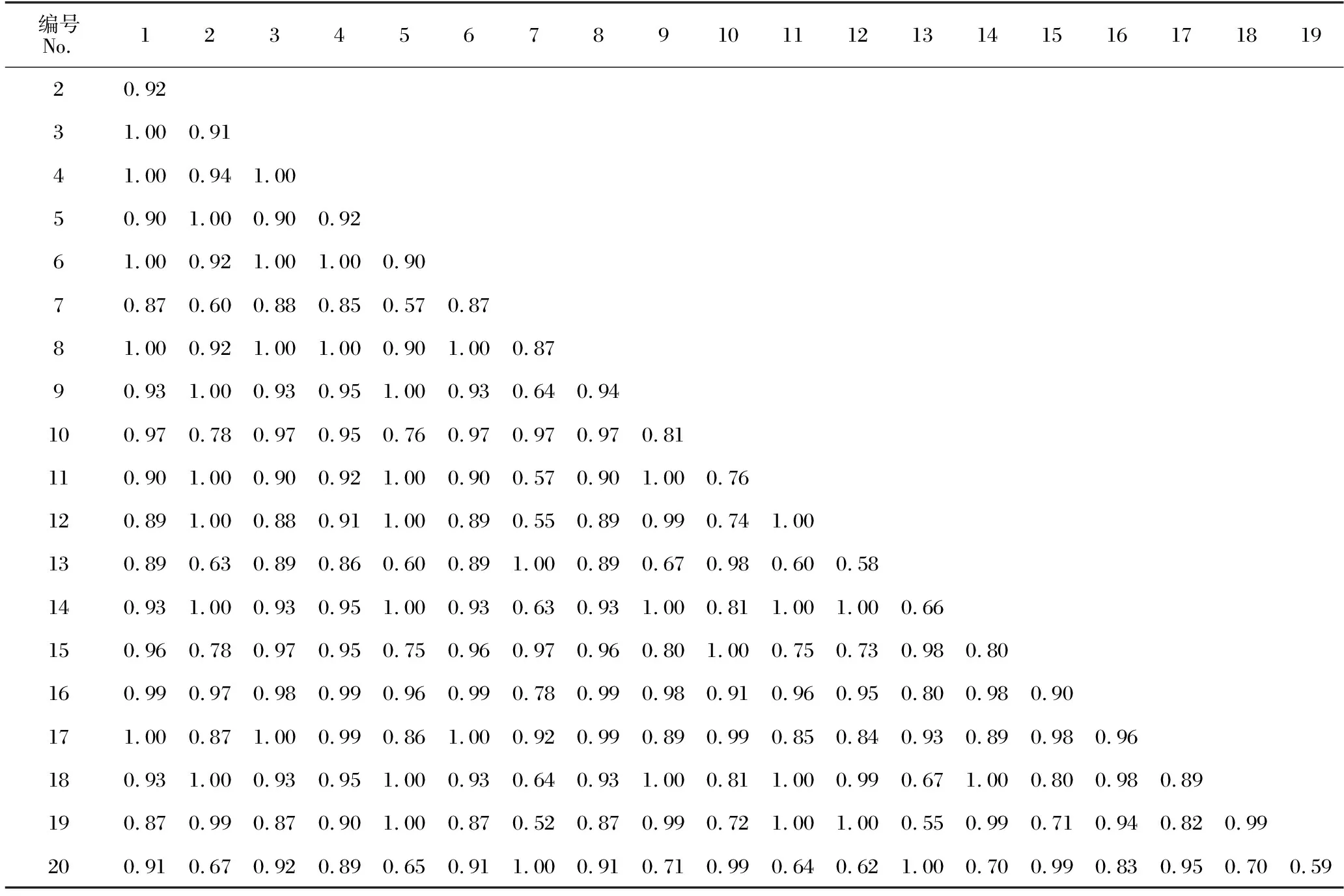
表7 過熟林林下灌木層物種生態位重疊指數
3 討論
3.1 主要物種重要值與生態位寬度
重要值與生態位寬度指數均可表征物種在群落中的地位和作用,但又有所不同[16]。重要值是通過簡單量化的方式來反映物種在群落中的地位和作用的綜合指標,利用重要值確定群落中的優勢種[17],體現的是物種在群落中的優勢程度[18]。生態位寬度可作為衡量物種或種群適應環境和利用資源的實際廣度和潛能的指標,也可表示物種在群落中的地位[19],生態位寬度指數的大小反映的是物種在群落中的資源利用能力和對生態的適應能力[20]。大量研究表明,通常物種的重要值較大,其生態位寬度也較大[21]。本研究中馬尾松中齡林亞喬木層的山烏桕和成熟林亞喬木層的狹基潤楠重要值最大,生態位寬度也最大,但是重要值與生態位寬度的排序并不完全相同,如中齡林亞喬木層的長毛柃重要值大于楓香,但是其生態位寬度卻小于楓香;成熟林亞喬木層的黃連木重要值大于黃牛木,但其生態位寬度卻小于黃牛木。這一現象與馬姜明等[8]在研究漓江流域巖溶區檵木群落不同恢復階段主要共有種生態位變化中的結論相似。這可能是因為重要值不是唯一影響物種生態位寬度的因素,分布頻度同樣可以影響物種生態位寬度的大小。物種在資源位上出現的頻度越大,說明其對環境的適應性越強,分布范圍越廣,所以就會出現物種重要值雖小,但生態位寬度卻較大的情況。
一般而言,物種的生態位寬度越大,其生態位特化程度越低,更傾向于泛化種,通常就是群落的優勢種[22]。本研究發現中齡林亞喬木層的山烏桕、灌木層的鵝掌柴,成熟林亞喬木層的狹基潤楠、灌木層的三椏苦,過熟林亞喬木層的鵝掌柴、灌木層的紐子果,在各自的群落中對資源的利用能力和環境適應能力強,在群落中分布廣泛,物種個體數量多,所以它們的生態位寬度較大,在群落中屬于優勢種群。而中齡林亞喬木層的黃毛榕、灌木層的藤黃檀;成熟林亞喬木層的黃連木、南酸棗、木油桐、山烏桕,灌木層的臺灣榕;過熟林亞喬木層的石楠、芳槁潤楠,灌木層的香楠等分布較為集中,種群數量少,重要值較小,在群落中的地位較低,對環境適應能力較差,生態位寬度較小,說明這些物種分布范圍較窄,傾向于特化種[23],在群落演替進行階段容易被其他物種所取代。
3.2 生態位寬度與生態位重疊
生態位重疊指的是兩個或兩個以上的生態位相似的物種生活于同一空間時分享或競爭共同資源的現象[24]。生態位重疊指數可表征物種間對同一資源利用能力的差異性和競爭關系,生態位重疊指數越大說明物種間對資源的需求越相似,物種間可能會產生激烈的競爭[25]。往往生態位寬度大的物種,都會產生較大的生態位重疊[26,27]。本研究中馬尾松中齡林亞喬木層的山烏桕與鵝掌柴,灌木層的三椏苦和火力楠之間的相互關系符合這一規律。生態位寬度較小的兩個物種間有較高的生態位重疊指數,如中齡林亞喬木層的牛耳楓和樟,成熟林亞喬木層的南酸棗和山烏桕等,說明生態位重疊與生態位寬度之間沒有絕對的正相關關系。這可能與馬尾松人工林林窗的形成有關。相關研究表明[28],林窗的形成可改變馬尾松人工林林下的小生境。由于生境的差異性,有些物種會傾向于在局部適宜的區域有較高的聚集度,比如牛耳楓、樟、南酸棗和山烏桕均為喜光植物,在林窗處生長良好,而適宜生境外的地方分布較少,以致于這些物種生態位寬度較小,但具有較高的生態位重疊。
本研究中,各群落物種生態位重疊指數均較大,反映的是馬尾松林下物種生態位特性較為相似,能較好地利用林下資源,這與康冰等[12]的研究結論一致。隨著近自然恢復演替的進行,馬尾松人工林亞喬木層物種生態位重疊程度逐漸降低,如中齡林生態位重疊指數均值為0.74,成熟林均值為0.53,過熟林均值為0.50;灌木層物種生態位重疊程度隨著演替進行而增加,如中齡林生態位重疊指數均值為0.74,成熟林均值為0.76,過熟林均值為0.89。造成該現象的原因可能是在演替的過程中亞喬木層比灌木層更早地進入穩定階段。相關研究表明[29],隨著森林群落的恢復,喬木層在針闊混交林階段群落結構趨于穩定,灌木層需演替到常綠闊葉林階段結構才穩定。亞喬木層物種隨著群落演替的進行,生態位的分化程度越高,物種間的競爭逐漸變弱,林下亞喬木層物種間關系趨向穩定,所以在群落中隨著演替的進行物種間的生態位重疊指數均值出現下降的趨勢。這一結論與張明霞等[30]在秦嶺天然次生林優勢種群的研究中所得出的,在針闊混交林階段,喬木層優勢種群種間聯結較為松散,具有一定的獨立性,基本趨于穩定,灌木層優勢種群則處于動態演替的不穩定階段的結論基本一致。而灌木層物種由于受可利用資源和自身生物學特性的限制,擴散范圍相對較小,物種分布較為集中,物種種類比同恢復階段的喬木層更為豐富[31],物種間的生態位分離程度較低,導致隨著演替的進行灌木層物種的生態位重疊指數均值增加。
4 結論
隨著近自然恢復的進行,馬尾松人工林林下優勢種發生改變,優勢種的變化趨勢為強生陽性樹種-陽性樹種-半陽性樹種。亞喬木層中,中齡林優勢種為山烏桕,成熟林為狹基潤楠,過熟林為鵝掌柴;灌木層中,中齡林優勢種為鵝掌柴,成熟林為三椏苦,過熟林為紐子果;隨著亞喬木層物種間生態位特化程度增加,物種間的生態位重疊程度逐漸降低,灌木層物種可利用資源受限,各物種間資源利用相似度增加,生態位重疊程度逐漸升高,亞喬木層較灌木層具有更高的穩定性。在人工林近自然恢復演替進程中,宜優化物種間的配置,充分利用資源。物種的選擇上,將生態位寬度大的物種作為先鋒物種,以與其生態位重疊小的物種作為伴生種,以加速近自然恢復演替的進行,同時又能保持群落物種的多樣性。群落物種間的配置需注意,不可將生態位重疊很大的物種配置在一起,以減少物種間的競爭,同時也要注意避免將生態位重疊程度較小的物種配置在一起,充分利用林下資源,維持群落的穩定性,加速人工林近自然恢復演替的進行。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
小學科學(學生版)(2020年4期)2020-05-21 07:30:46
小學科學(學生版)(2020年3期)2020-03-25 13:31:22
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26