利用微滴數字PCR技術分析轉基因大豆‘GE-J12’中外源基因的拷貝數
2022-01-10 07:44:32李瑞環曹英芳蘭青闊
中國農業大學學報 2022年1期
關鍵詞:大豆
趙 新 劉 雙 劉 娜 李瑞環 曹英芳 蘭青闊 王 永*
(1.天津市農業科學院 生物技術研究所,天津 300384;2.河北農業大學 食品科技學院,河北 保定 071001)
伴隨轉基因技術的發展,轉基因作物及其衍生食品的安全性問題受到社會強烈的關注,國家監管部門對轉基因作物的上市和推廣均制定了明確的政策和法規,并對其開展嚴格的生物安全評價。載體全序列分析、目的基因整合和外源插入序列表達等分子特征是轉基因生物安全評價的重要組成內容之一,其中轉基因作物中插入外源基因的拷貝數鑒定是關鍵參數。外源基因整合進入受體基因組的位置和拷貝數會影響目的基因的表達和遺傳穩定性,而插入基因的拷貝數相對其插入位點而言影響更為重要。當外源基因以低拷貝數(1~2個)整合到受體基因組時,通常能夠穩定高效轉錄表達,而多拷貝數的整合則會造成基因不穩定表達,甚至沉默。因此,鑒定轉基因作物的外源基因插入拷貝數非常重要。
目前,Southern 印跡雜交(Southern blot)和實時熒光定量PCR(Quantitative real-time PCR, qRT-PCR)是鑒定轉基因作物外源基因拷貝數常用的2種分析技術, 已廣泛應用。但這2種技術都存在一定缺陷,Southern blot方法操作步驟繁瑣、工作量大、檢測周期長、對試驗人員操作技術要求較高;qRT-PCR方法需要依賴標準物質,構建標準曲線,而利用標準曲線進行定量本身就是一種并不十分精確的相對定量方法,且構建標準曲線的過程中涉及體系的摸索和優化,也耗費大量的時間和精力,因此qRT-PCR不是理想的轉基因作物外源基因拷貝數的鑒定方法。微滴數字PCR(droplet-based digital PCR,ddPCR)是最新興起的一項核酸檢測技術,該技術不依賴任何校準物,無需構建標準曲線,以形成大量油包水微滴的形式對核酸進行成千上萬倍稀釋,然后以每個小微滴為獨立的反應單元進行PCR反應,最后利用泊松分布原理,實現對核酸檢測的絕對定量。在檢測目的基因拷貝數時,利用目標基因與內標基因的雙重反應,通過計算其比值,即可快速獲得目標基因拷貝數。相比其他方法,該方法操作簡便、快速高效、準確性高、重復性好,已成為檢測外源基因拷貝數的首選方法。諸多研究表明,dd PCR已用于玉米和水稻等作物轉基因插入拷貝數的檢測。
抗除草劑轉基因大豆‘GE-J12’是由中國農業科學院作物科學研究所研發的轉G2-EPSPS
基因和GAT
基因抗除草劑大豆新品種。采用農桿菌介導轉化法,將轉G2-EPSPS
和GAT
的2個基因的表達載體DNA導入大豆受體中,通過多代篩選獲得高耐草甘膦轉基因大豆新品種。培育和推廣耐草甘膦除草劑大豆新品種,對防除大豆田間雜草、降低生產成本,提高農民種豆積極性和種豆效益,增強我國大豆競爭力具有重要意義。目前,對于該轉基因大豆新品種的外源基因插入拷貝數的數字PCR分析方法的研究尚未見報道。本研究以‘GE-J12’的G2-EPSPS
和GAT
外源插入目的基因序列、3′端轉化體特異性序列為靶標,以Lectin
為內標基因,通過引物探針篩選、特異性測試分析,旨在建立微滴數字PCR拷貝數分析方法,以期為該轉化體的安全評價提供參考。1 材料與方法
1.1 材料及儀器
抗除草劑轉基因大豆品種‘GE-J12’,非轉基因大豆品種‘Jack’,由中國農業科學院作物科學研究所研發;轉基因玉米品種混合樣(‘MON863’、‘MON810’、‘MON88017’、‘MON87427’、‘MON87460’、‘MON89034’、‘雙抗12-5’、‘Bt176’、‘Bt11’、‘MIR604’、‘MIR162’、‘GA21’、‘DAS40278-9’、‘NK603’、‘TC1507’、‘T25’、‘3272’、‘4114’、‘5307’、‘59122’、‘C0030.3.5’、‘C0010.3.7’和‘IE09S034’每種轉化體質量分數為1%,以非轉基因玉米為填充物),其他轉基因大豆品種混合樣(‘A2704-12’、‘A5547-127’、‘CV127’、‘DAS68416-4’、‘FG72’、‘GTS40-3-2’、‘MON87705’、‘MON87769’、‘MON88302’、‘MON89788’、‘SHZD32-1’、‘73496’、‘356043’和‘305423’每種轉化體質量分數為1%,以非轉基因大豆為填充物),轉基因油菜品種混合樣(‘MON88302’、‘RF1’、‘RF2’、‘RF3’、‘Ms1’、‘Ms8’、‘Topas19/2’、‘T45’、‘oxy235’和‘73496’每種轉化體質量分數為1%,以非轉基因油菜為填充物),轉基因水稻品種混合樣(‘科豐2號’、‘科豐6號’、‘科豐8號’、‘克螟稻’、‘TT51-1’、‘T1C-19’、‘G6H1’、‘M12’每種轉化體質量分數為1%,以非轉基因水稻為填充物),轉基因棉花品種混合樣(‘COT102’、‘GHB614’、‘LLCOTTON25’、‘MON1445’、‘MON15985’、‘MON88913’、‘MON531’每種轉化體質量分數為1%,以非轉基因棉花為填充物),由本實驗室收集保存。
1.2 試驗方法
1
.2
.1
引物及探針的設計轉基因大豆‘GE-J12’的外源基因插入位置在大豆3號染色體上,插入位點包含1個G2-EPSPS
和1個GAT
基因的表達框,不同外源基因片段之間以大豆基因組DNA片段相連接,見圖1。本研究以抗除草劑轉基因大豆‘GE-J12’的G2-EPSPS
和GAT
外源插入基因序列,T-DNA與大豆基因組連接區域的3′端轉化體特異性序列為靶標,以Lectin
為內標,應用Primer Express 3.0設計引物和探針。引物和探針由上海生工生物工程有限公司合成,見表1。
圖1 抗除草劑轉基因大豆‘GE-J12’外源基因插入位點
表1 PCR引物和探針序列
Table 1 PCR primers and probe sequences
名稱Name引物序列(5′→3′)Primersequence(5′→3′)片段大小/bpSize序列來源SequencesourceLectin基因QF:5′-GCCCTCTACTCCACCCCCA-3′QR:5′-GCCCATCTGCAAGCCTTTTT-3′QP:5′-FAM-AGCTTCGCCGCTTCCTTCAACTTCAC-BHQ1-3′118農業部2031號公告-8-2013G2-EPSPS基因QF:5′-TCGAGATTGATGGTGGTTTGTC-3′QR:5′-TCAGCGCCACTTCAATCG-3′QP:5′-FAM-GGGCAAACTGATTTCCATAGCTT-BHQ1-3′ 90自行設計GAT基因QF:5′-GGGCAAACTGATTTCCATAGCTT-3′QR:5′-CCTCGGAGCTGGTACTGTTTCT-3′QP:5′-FAM-ATTCCACCAGGCCGAGCACTCAGA-BHQ1-3′ 81自行設計3′轉化體特異性序列QF:5′-GCAGCTTGAGCTTGGATCAGA-3′QR:5′-TCATAGCGTGGTGTTTGACATAAA-3′QP:5′-FAM-TGTCGTTTCCCGCCTTCAGTTTAAACC-BHQ1-3′ 91自行設計
1
.2
.2
DNA模板的提取用TIANGEN高效植物基因組DNA提取試劑盒(天根生化科技北京有限公司),提取轉基因大豆品種‘GE-J12’的DNA;非轉基因大豆受體品種‘Jack’、轉基因玉米混合樣、其他轉基因大豆混合樣、轉基因油菜混合樣、轉基因水稻混合、轉基因棉花混合樣,各取100 mg粉末樣品,用TIANGEN高效植物基因組DNA提取試劑盒,提取DNA。用美國Bio-Rad公司的NanoDrop-100核酸蛋白定量儀測定DNA的濃度和質量,將DNA濃度統一稀釋至25 ng/μL,備用。
1
.2
.3
微滴數字PCR檢測體系的建立在20 μL的PCR反應體系中,包括2×ddPCR Supermixfor probes預混液10 μL,上下游引物和探針各1 μL,模板1 μL,無菌超純水補足20 μL,混勻后離心。將其轉移至微卡發生器,切忌產生氣泡,同時與微卡發生器相應位置加入重油70 μL,蓋上墊片,放入微滴生成器中生成微滴。吸取40 μL生成的微滴,轉移至數字PCR 96孔反應板中,170 ℃熱封后,在美國Bio-Rad公司的QX-100微滴數字PCR儀上開始擴增循環,PCR儀爬坡速度設置為2 ℃/s,循環參數為:95 ℃,10 min;94 ℃,30 s,60 ℃,1 min,40 個循環;98 ℃,10 min。
1
.2
.4
方法特異性檢測以轉基因玉米品種混合樣,其他轉基因大豆品種混合樣,轉基因油菜品種混合樣,轉基因水稻品種混合樣,轉基因棉花品種混合樣和非轉基因大豆品種樣品的基因組DNA為模板,以去離子水作為陰性對照,按1.2.3反應程序及條件進行微滴數字PCR擴增,對檢測方法的特異性進行測試,每個反應設置2個重復。
1
.2
.5
微滴數字PCR拷貝數分析以抗除草劑轉基因大豆‘GE-J12’的3個單株基因組DNA為模板,以大豆內標基因Lectin
為參照,應用構建的微滴數字PCR檢測體系,對3′轉化體特異性序列、G2-EPSPS
和GAT
外源插入基因進行微滴數字PCR分析,測定每個基因的拷貝數濃度,每個反應設置2次重復。1
.2
.6
微滴數字PCR拷貝數計算微滴數字PCR儀自動計算每個反應的拷貝數濃度、總微滴數和陽性微滴數,總微滴數≥10 000視為有效結果,按照公式計算出試樣中轉基因大豆的外源目的基因在基因組插入的拷貝數。公式如下:
式中:A
,轉基因大豆的外源目的基因在基因組插入的拷貝數;B
,轉基因大豆的外源目的基因的拷貝數濃度,個/μL;C
,內標基因Lectin
基因的拷貝數濃度,個/μL。1
.2
.7
Southern blot方法探針制備PCR擴增使用G2-EPSPS
和GAT
基因引物,見表2,由上海生工生物工程有限公司合成。總體積為50 μL,包括10×buffer(plus Mg)預混液5 μL、dUTP標記混合物5 μL、上下游引物各1 μL、Taq酶 1 μL、1 μL質粒DNA、ddHO補足50 μL,進行PCR擴增,擴增結束后,對擴增產物進行電泳檢測(1% 瓊脂糖)及地高辛探針制備。根據轉基因大豆‘GE-J12’轉化載體全序列信息和酶切位點信息,選擇常用的內切酶進行基因組 DNA酶切,酶切體系總體積為600 μL,包括 10×Buffer 60 μL,BSA 60 μL,Hin
d III/Spe I/Xba
I/Dra
I內切酶30 μL,基因組 DNA 10 μg。探針和酶切產物經電泳、轉膜和雜交等后續步驟獲得 Southern雜交結果。表2 Southern 雜交檢測引物
Table 2 Southern blot PCR primers sequences
探針名稱Name引物序列(5′→3′)Primersequence(5′→3′)片段大小/bpSize序列來源SequencesourceTP3TP3-F:GCTTCTATCGGCTGGTTTP3-R:GCCAATACGCAAACCGC902GATTP5TP5-F:ACAAACCACCATCAATCTP5-R:CGTTGTTAGCCTTGCGG724G2-EPSPS
2 結果與分析
2.1 引物和探針的特異性試驗
由圖2可知,只有以抗除草劑轉基因大豆‘GE-J12’的基因組DNA為模板時才能獲得陽性微滴和對應的拷貝數,且陽性微滴與陰性微滴界限明顯,而以其他轉基因作物品種混樣和其他非轉基因大豆品種植株受體的基因組DNA為模板均無陽性微滴,說明本方法篩選的引物探針特異性良好,可以用于檢測抗除草劑轉基因大豆‘GE-J12’的外源插入基因的拷貝數。

1和2,空白對照;3和4,轉基因大豆‘GE-J12’;5和6,非轉基因大豆(受體);7和8,轉基因玉米混合樣;9和10,其他轉基因大豆混合樣;11和12,轉基因油菜混合樣;13和14,轉基因水稻混合樣;15和16,轉基因棉花混合樣。
2.2 微滴數字PCR拷貝數分析
3′轉化體特異性序列可在分析外源基因拷貝數的同時,通過與大豆內標基因Lectin
的比值,作為植株純合性鑒定的引物探針,純合陽性植株接近于1個拷貝,雜合植株接近于0.5個拷貝。由表3可知,經微滴數字PCR驗證,轉基因大豆品種‘GE-J12’的 3個單株中,以Lectin
作為內標,3’轉化體特異性序列的拷貝數比值分別為0.96、1.03和1.02,均接近于1個拷貝,驗證結果顯示,轉基因大豆品種‘GE-J12’的 3個單株均為純合陽性轉基因植株;3個單株外源基因G2-EPSPS
和GAT
的拷貝數比值分別為1.02、1.16、0.81和1.04、1.12、0.89,均接近于1個拷貝,參照‘GE-J12’研發者提供和全序列分析結果,驗證轉基因大豆‘GE-J12’的載體T-DNA在大豆基因組上為單拷貝插入,具體拷貝數濃度,見表3,微滴擴增結果,見圖3、圖4和圖5。表3 轉基因大豆‘GE-J12’數字PCR檢測分析
Table 3 Analysis of digital PCR detection of transgenic soybean‘GE-J12’
基因Gene外源序列拷貝數/(個/μL)Foreignsequencecopynumber內源Lectin拷貝數/(個/μL)EndogenousLectincopynumber拷貝數比值Copynumber單株拷貝數均值Averagecopynumberperplant619.50606.501.02G2-EPSPS614.00531.501.160.99658.50813.000.81633.50606.501.04GAT595.00531.501.121.01721.00813.000.89582.00606.500.963’旁側序列546.50531.501.031.00831.00813.001.02
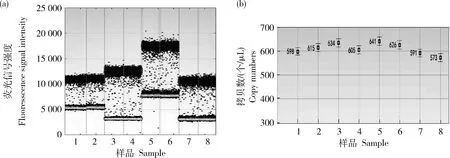
1和2,Lectin基因;3和4:G2-EPSPS基因;5和6,GAT基因;7和8,3′轉化體特異序列。下同。
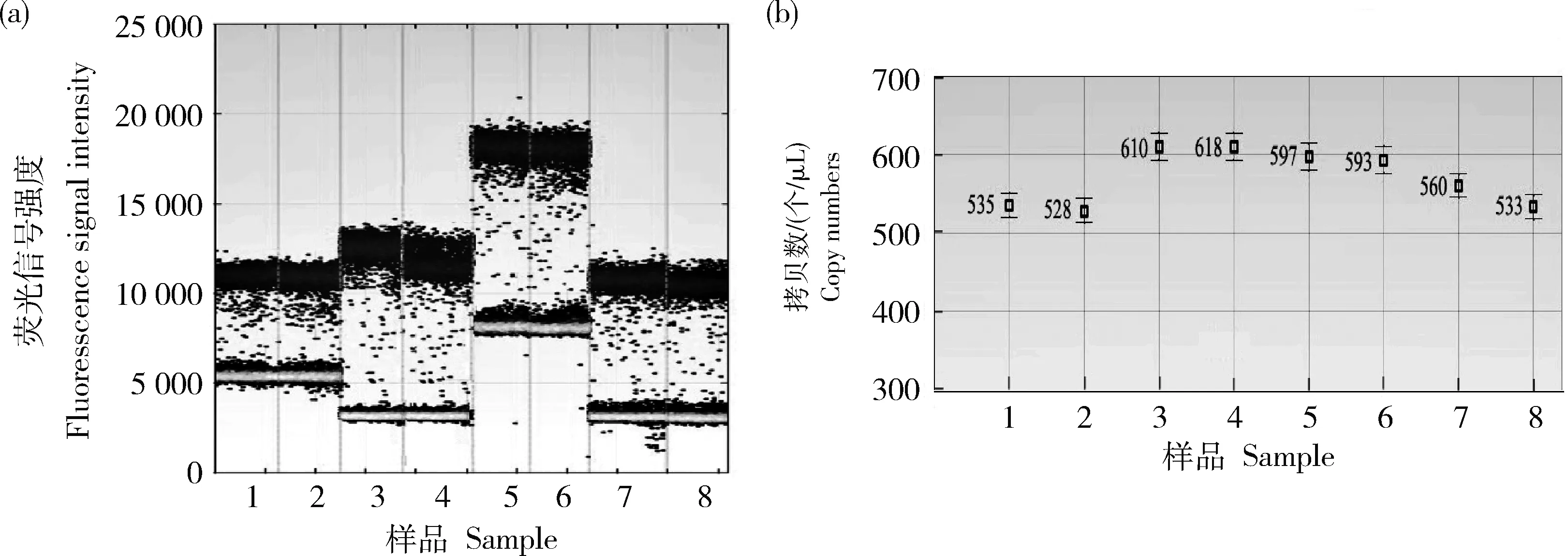
圖4 轉基因大豆‘GE-J12’2號單株的數字PCR擴增微滴圖(a)和拷貝數濃度圖(b)
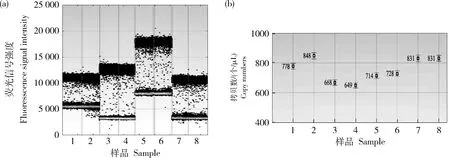
圖5 轉基因大豆‘GE-J12’3號單株的數字PCR擴增微滴圖(a)和拷貝數濃度圖(b)
2.3 Southern blot結果
由圖6可知,應用上述Hin
d III、Spe
I、Xba
I、Dra
I等4種內切酶酶切后轉基因大豆品種‘GE-J12’和非轉基因大豆品種‘Jack’的基因組DNA泳道均呈彌散狀,無明顯條帶,說明基因組 DNA 酶切效果較好。由圖7可知,用G2-EPSPS
和GAT
擴增產物為雜交探針,對酶切產物進行Southern blot檢測,質粒陽性對照雜交結果正常,非轉基因大豆品種(受體)‘Jack’經Hin
d III、Spe
I、Xba
I和Dra
I的酶切雜交后均未出現雜交信號,轉基因大豆品種‘GE-J12’經上述4種酶切雜交后,雜交片段大小與預期大小一致(Hind III、Spe I、Xba I、Dra
I酶切后預期雜交片段大小分別為4 859、8 638、11 236和4 754 bp),且探針雜交信號均為1個,說明目的基因的拷貝數為1個。
M,1 kb DNA Ladder;1:非轉基因大豆受體(Hind III),2,轉基因大豆‘GE-J12’(Hind III);3,非轉基因大豆受體(Spe I);4,‘GE-J12’(Spe I);5,非轉基因大豆受體(Xba I);6,‘GE-J12’(Xba I);7,非轉基因大豆受體(Dra I);8,‘GE-J12’(Dra I)。
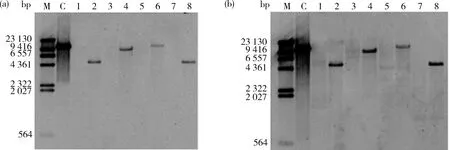
M,DNA molecular-weight marker;C,陽性質粒;1,非轉基因大豆受體(Hind III);2,轉基因大豆‘GE-J12’(Hind III);3,非轉基因大豆受體(Spe I);4,‘GE-J12’(Spe I);5,非轉基因大豆受體(XbaI);6,‘GE-J12’(XbaI);7,非轉基因大豆受體(Dra I);8,‘GE-J12’(Dra I)。
3 討 論
目前的轉基因作物研究中,能夠準確、快速地鑒定轉基因作物中外源基因的拷貝數,已經成為轉基因作物檢測的關鍵內容。傳統的Southern blot方法操作復雜、檢測周期較長,通常需要2周以上。該方法不僅對DNA的純度和濃度要求較高(根據不同的作物DNA的需求量在20~100 μg),而且對人員操作的技術要求比較嚴格,期間很多中間環節(如,DNA提取質量不佳、酶切效果不好、轉膜不充分、雜交條件或漂洗不能使陽性結果與背景產生明顯反差等)都可能導致無法得到準確的檢測結果,且重復性差,使原本較長的檢測周期二次延長,且容易在內切酶的選擇上受到限制,因此,自2014年來轉基因作物中外源基因的拷貝數鑒定方法,逐漸被數字PCR等新興技術所取代。
數字PCR技術已經在轉基因水稻、轉基因玉米和轉基因小麥等轉基因作物的外源基因拷貝數鑒定中廣泛應用,且在基因表達研究、癌癥標志物稀有突變檢測和轉基因成分鑒定等諸多領域顯示出廣闊的應用前景。本研究基于微滴數字PCR技術,建立了轉基因大豆‘GE-J12’中外源基因拷貝數的分析方法。該方法操作步驟簡便,僅需DNA提取、PCR擴增、微滴生成與讀取等常規步驟,即可在6~8 h完成對轉基因作物外源基因拷貝數的鑒定,不同試驗人員間可穩定重復。同時該方法對DNA需求量最低只需200 ng,遠遠低于Southern雜交所需DNA的量,且結果與Southern雜交一致。因此本方法具有快速、簡便、高效、不依賴任何標準品、所需樣本量少、不受內切酶位點選擇限制等優點。
在進行轉基因作物微滴數字PCR拷貝數鑒定之前,通常需在該作物外源插入序列的兩側基因組DNA序列上和外源插入序列的5′或3′設計2對定性PCR引物,且2對引物共用同一上游或下游引物,應用這2對引物的擴增結果對植株進行純合性鑒定。而本研究在建立轉基因大豆‘GE-J12’中外源基因拷貝數分析方法的同時,應用3′端轉化體特異性序列可監測植株純合性,無需進行前期的定性PCR引物設計及擴增,簡化了試驗步驟,且保證了試驗結果的準確性。該方法不僅僅局限于外源目的基因G
2-EPSPS
和GAT
,同時也可應用于該作物基因型整合鑒定中所含有的CaMV
35S
啟動子、CaMV
35S
終止子和NOS
終止子等調控元件的拷貝數分析,從而實現通過一步反應對該品種T-DNA外源插入片段不同部位多種整合基因型拷貝數的全面分析。后續試驗可對多重微滴數字PCR檢測體系的建立進行摸索,進一步提高試驗準確性、節約試劑成本、提高試驗效率,同時對比NGS技術分析外源插入序列拷貝數,為傳統方法無法解析的復雜整合體的分子特征奠定試驗基礎。4 結 論
基于微滴數字PCR技術,以轉基因大豆‘GE-J12’轉化體的外源插入目的基因G2-EPSPS
和GAT
的序列、3′轉化體特異性序列為靶序列,設計了3對引物及探針,以大豆Lectin
內標基因為參照,建立了微滴數字PCR拷貝數檢測方法。應用該方法,鑒定G2-EPSPS
基因、GAT
基因和3′轉化體特異性序列的拷貝數均值分別為0.99、1.01和1.00,均接近于1個拷貝,結果表明,轉基因大豆‘GE-J12’的外源基因在大豆基因組上為單拷貝插入,且為純合植株。該方法特異性強、準確性高、重復性穩定,操作簡便、快速,在6~8 h完成對轉基因作物外源基因拷貝數的鑒定。猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
