選擇性多聚腺苷酸化及microRNAs對綿羊ACSL1基因表達的影響
2022-01-10 07:44:46張立春于永生馬惠海
中國農(nóng)業(yè)大學學報 2022年1期
關鍵詞:檢測
曹 陽 張立春 于永生 馬惠海 劉 宇 曹 陽*
(1.吉林省農(nóng)業(yè)科學院 動物生物技術研究所,吉林 公主嶺 136100;2.浙江省農(nóng)業(yè)科學院 畜牧獸醫(yī)研究所,杭州 310021)
許多基因在轉錄過程中存在多個多聚腺苷酸化位點,從而形成不同長度的3′UTR現(xiàn)象,稱為選擇性多聚腺苷酸化(Alternative polyadenylation,APA)。APA在高等真核生物的轉錄中是一種普遍存在的現(xiàn)象,人類中50%以上、小鼠30%以上的基因存在APA現(xiàn)象,APA調(diào)控基因表達,從而調(diào)控該基因的特定生物學功能。基因不同長度的3′UTR 會調(diào)控蛋白表達,較短3′UTR的轉錄本會逃避可能與長3′UTR結合的microRNAs的抑制,并且不同長度的3′UTR轉錄本的亞細胞定位不同。研究表明,APA對轉錄活性的調(diào)控似乎是一種廣泛機制,細胞外刺激同樣能誘導APA現(xiàn)象的發(fā)生,因此,APA在調(diào)控基因穩(wěn)定、翻譯和轉運過程中有重要作用,特別是3′UTR上與microRNAs結合的位點。MicroRNA是指長度為22個核苷酸的非編碼單鏈RNA,通過基因3′UTR 上的結合位點調(diào)控基因表達,在細胞受到刺激和增殖過程中都會發(fā)生變化。MicroRNAs同樣被證明是潛在脂肪形成的調(diào)節(jié)劑,通過與靶向基因結合調(diào)控基因表達,這一特征在脂肪細胞分化和脂滴的生成這些脂質(zhì)代謝中有調(diào)節(jié)作用,miR-27a、miR-22、miR-335、miR-122和miR-155等microRNAs都被證明對脂肪的生成存在抑制作用。因此,APA和microRNAs在基因表達過程中的作用值得更深入研究。
ACSLs是酯酰CoA合成所必需的酶。它能夠活化游離脂肪酸合成脂酰CoA將并引導到特定的代謝通路中。ACSL
1含有多個啟動子,導致ACSL1
存在多個轉錄本,不同轉錄本在不同組織中具有不同的功能。許多microRNAs會通過與ACSL1
基因結合來影響脂肪沉積。有研究表明miR-205能通過靶向ACSL1
基因來調(diào)節(jié)肝癌細胞的脂代謝,破壞肝癌細胞的脂肪生成。Bta-miR-181a通過靶向ACSL
1基因促進牛乳腺上皮細胞中TAG的合成,從而調(diào)節(jié)牛乳脂肪的合成。在肉雞腹部脂肪沉積的研究中發(fā)現(xiàn)miR-19b-3P能夠靶向ACSL1
基因,使ACSL1
基因下調(diào)并促進脂肪細胞的增殖。因此,microRNAs對綿羊ACSL1
基因表達的影響同樣值得探究。本研究通過對ACSL1
基因3′UTR克隆和雙熒光素酶檢測確定microRNAs對ACSL1
基因表達的影響,明確綿羊ACSL1
基因表達規(guī)律,以期為分析ACSL1
基因在綿羊脂肪代謝的作用提供理論依據(jù)。1 材料與方法
1.1 試驗材料
試驗所用綿羊前體脂肪細胞、293T細胞和脂肪組織儲存于吉林省農(nóng)業(yè)科學院動物生物技術研究所。
1.2 試驗方法
1
.2
.1
cDNA末端快速擴增技術根據(jù)Genbank公布的ACSL1
基因序列信息(登錄號:XM_015104562.2),依照RACE引物設計原則,使用Primer Premier 6.0設計引物SP1、SP2、SP3和SP4。使用 RACE試劑盒(Takara)進行擴增。1
.2
.2
RT-
PCR根據(jù)克隆測序所得的序列設計引物,檢測不同長度的3′UTR在誘導分化過程中表達量的變化。引物序列如下表1所示。
表1 本研究中用到的引物信息
Table 1 Information of primers used in the study
引物名稱Primername引物序列(5′-3′)Sequence(5′-3′)目的片段長度/bpProductsizeSP1CCTTGGCAGCCAGATAATTCASP2GGACAAGCAAACACCACGCTGA450SP3CTGGCACAAGGGGAGTACATAGCTCC750SP4TTCGGAATTATTTCAGGTCACAGATCGATG1570ACSL1-as-FACSL1-as-RCAGTGAAGCAAAGGGCAGAGGCGACAGATGGAGGAA160ACSL1-al-FACSL1-al-RAACTGTCCACTTTTCCTTTGGCATTTATTTATCACCATCC215
熒光定量PCR反應體系及條件:2×PCR Mix 10 μL,上下游引物各10 mmol/L,cDNA 1 μL,ddHO補齊至20 μL。95 ℃5 min,95 ℃ 10 s,60 ℃ 15 s,72 ℃ 20 s,從第二步開始40個循環(huán)。95 ℃ 5 s,65 ℃1 min,然后溫度以5 ℃/s的速率從65 ℃增加到97 ℃,最后溫度降到40 ℃。
1
.2
.3
載體構建microRNAs模擬物、抑制物購自上海吉瑪制藥技術有限公司。通過克隆得到不同長度的3′UTR,分別構建pmiR-GLO 載體,載體構建由上海吉瑪制藥技術有限公司完成。在ACSL1
基因3′UTR 1 264~1 271 bp處定點突變,突變序列由上海吉瑪制藥技術有限公司生物合成和測序,并連接到pmiR-GLO載體上。ACSL
1基因過表達載體構建:以綿羊前體脂肪組織為模版,RT-PCR擴增目的片段,與pbi-CMV3載體雙酶切后(Bam
HI和Not
I),經(jīng)T4連接酶(Transgen)連接,轉化后挑取陽性克隆,PCR鑒定,送生工生物工程(上海)有限公司測序。1
.2
.4
雙熒光素酶活性檢測microRNAs模擬物與上述表達載體利用脂質(zhì)體法分別共轉染至293T細胞,細胞轉染后24 h,收集細胞檢測細胞內(nèi)雙熒光素酶活性(Promega)。去除舊培養(yǎng)基,PBS清洗2次,加入250 μL稀釋好的1×Passive lysis buffer,置于水平搖床室溫下晃動15 min,使細胞被動裂解。在管中加入100 μL Luciferase assay buffer Ⅱ,加入20 μL細胞裂解液,混合均勻后,檢測螢火蟲熒光素酶的活性。檢測后,在原管中迅速加入100 μL Stop&GloReagent,混勻檢測海腎熒光素酶活性。計算比值(螢火蟲熒光素酶的活性RULs1/海腎熒光素酶活性RULs2)。
1
.2
.5
Western blot使用裂解緩沖液(RIPA,Thermo Scientific)收集細胞蛋白質(zhì),并使用增強型BCA蛋白質(zhì)測定試劑盒(Beyotime)測定蛋白質(zhì)質(zhì)量濃度。蛋白質(zhì)樣品在95 ℃下變性10 min,在12%SDS-PAGE凝膠中分離,并在200 mA轉膜(PVDF)90 min。將膜用TBST洗滌3次,然后在室溫下用5%脫脂乳封閉2 h。加入一抗(兔抗,ACSL1,Bioss,貨號:bs5022R,1∶1 000)在4 ℃下孵育過夜,并與相應的二抗(羊抗兔,anti-rabbit,CST,貨號:14708,1∶2 000)在室溫下孵育1.5 h。將膜用TBST洗滌3次,并通過ECL-Plus試劑盒(Beyotime)處理后拍照。
1
.2
.6
統(tǒng)計分析進行3次獨立的生物學實驗,并計算相應的平均值。所有數(shù)據(jù)均以平均值±標準誤差的形式計算,所有統(tǒng)計分析進行t
檢驗。P
<0.05的差異為顯著差異,P
<0.01為極顯著差異。2 結果與分析
2.1 誘導分化不同時期ACSL1基因3′RACE擴增
通過3′RACE檢測ACSL1
3′UTR在誘導分化不同時期的表達,并克隆3′UTR序列,發(fā)現(xiàn)ACSL1
基因存在2個不同長度的3′UTR,長度分別為1 579和748 bp(圖1(a)),2個不同轉錄本具有相同的編碼區(qū)。ACSL1
基因2個長度的3′UTR在不同誘導分化時期都有表達。比對不同長度的3′UTR的核苷酸序列,兩者相差831 bp。帶有不同長度的3′UTR的ACSL1
基因分別命名為ACSL1-
al和ACSL1-
as(圖1(b))。
Lane M:DL2000 Plus DNA Marker; Lane 1、2、3:誘導分化0、4和8 d綿羊脂肪細胞引物SP4 3′RACE
qRT-PCR檢測不同長度3′UTR表達變化,綿羊ACSL1
基因2個不同長度的3′UTR表達量變化與ACSL1
基因趨勢相一致,隨著綿羊前體脂肪細胞被誘導分化,在誘導分化第4天達到最高值(圖2(a))。根據(jù)2種不同長度的3′UTR在誘導分化過程中表達量的比值發(fā)現(xiàn)在分化前期表達量比值下降(圖2(b)),所以ACSL1-
as的3′UTR表達量比ACSL1-
al增長速度更快,這表明ACSL1-
as在誘導分化前期更活躍。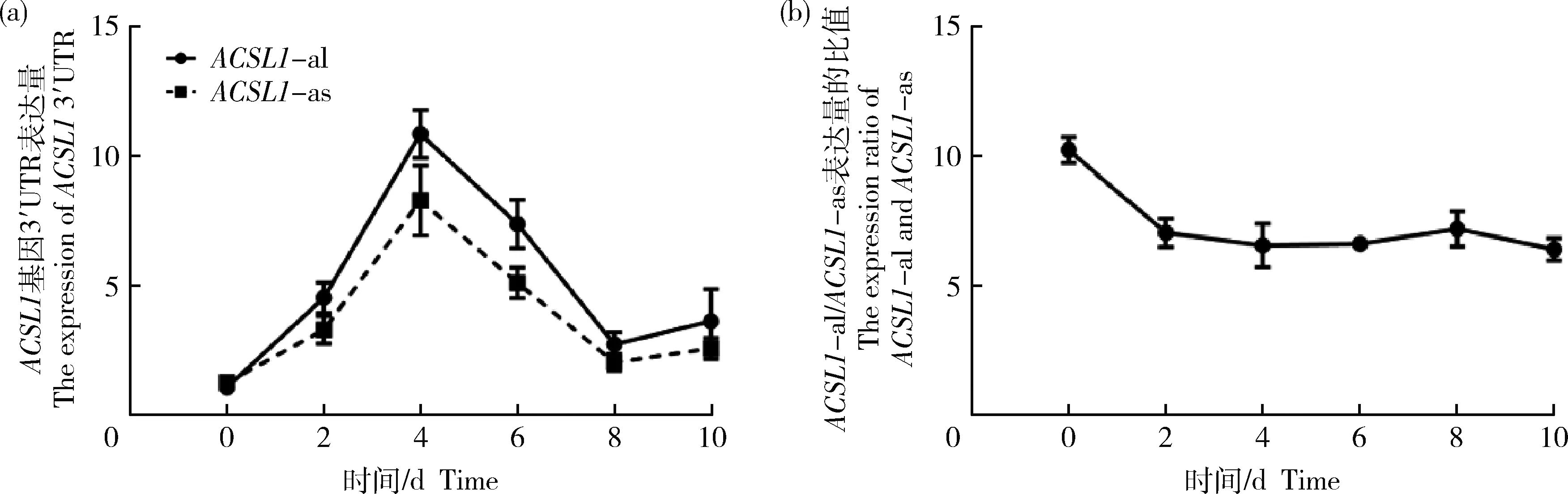
圖2 誘導分化不同時期ACSL1不同3′UTR表達量(a)和ACSL1-al與ACSL1-as表達量比值(b)
2.2 ACSL1不同長度3′UTR對基因表達的影響
為確定不同長度的3′UTR對ACSL1蛋白表達的影響,將過表達ACSL1-
al和ACSL1-
as 3′UTR 的pmiR-GLO載體轉染至293T細胞,24 h后檢測雙熒光素酶活性,結果顯示與ACSL1-
al 3′UTR 相比,ACSL1-
as 3′UTR具有更高的熒光素酶活性(圖3(a))。克隆不同長度3′UTR的ACSL1
基因,CDS區(qū)和3′UTR總長度分別為3 670和2 848 bp,并連接到pBI-CMV3載體中,轉染載體至293T細胞中,Western blotting檢測細胞內(nèi)ACSL1蛋白的表達。結果顯示,ACSL1-as蛋白表達量高于ACSL1-al(圖3(b)和(c))。表明因APA所產(chǎn)生的短3′UTR與長3′UTR相比更有助于ACSL1蛋白的表達。
*表示差異極顯著(P<0.05);**表示差異極顯著(P<0.01)。下同。
2.3 誘導分化不同時期microRNAs表達量的檢測
通過Targetscan預測ACSL1
3′UTR可能結合的microRNAs,挑選5個microRNAs進行驗證(圖4(a))。其中miR-202、miR-449a和miR-124a在ACSL1
-as 3′UTR、ACSL1
-al 3′UTR均存在結合位點,miR-218、miR-205只在ACSL1-
al 3′UTR存在結合位點。檢測microRNAs在誘導分化過程中表達量的變化,miR-449a和miR-124a的表達量在誘導分化前期(0~4 d)下降,后期(4~8 d)明顯上升,miR-218和miR-205表達量先下降后上升,與ACSL1
mRNA表達趨勢相反(圖4(b))。提示這些microRNAs可能與ACSL1
表達存在相關性。
圖4 microRNAs與ACSL1 3′UTR預測結合位點(a)和誘導分化不同時期綿羊脂肪細胞內(nèi)microRNAs表達量變化(b)
2.4 MicroRNAs對不同長度3′UTR和ACSL1表達的影響
為確定microRNAs對ACSL1
表達的影響,分別將miR-202 mimics、miR-449a mimics、miR-124a mimics、miR-218 mimics和miR-205 mimics與ACSL1-
al 3′UTR pmiR-GLO載體共轉染至293T細胞中,檢測雙熒光素酶活性,結果顯示共轉染miR-202、miR-449a、miR-124a和miR-218后細胞內(nèi)熒光素酶活性均顯著降低,其中與miR-218共轉染后活性降低極顯著(圖5(a))。然后將miR-202 mimics、miR-449a mimics、miR-124a mimics和ACSL1-
as 3′UTR pmiR-GLO載體共轉染至293T細胞中,檢測雙熒光素酶活性結果顯示共轉染后重組體的熒光素酶活性顯著降低(圖5(b))。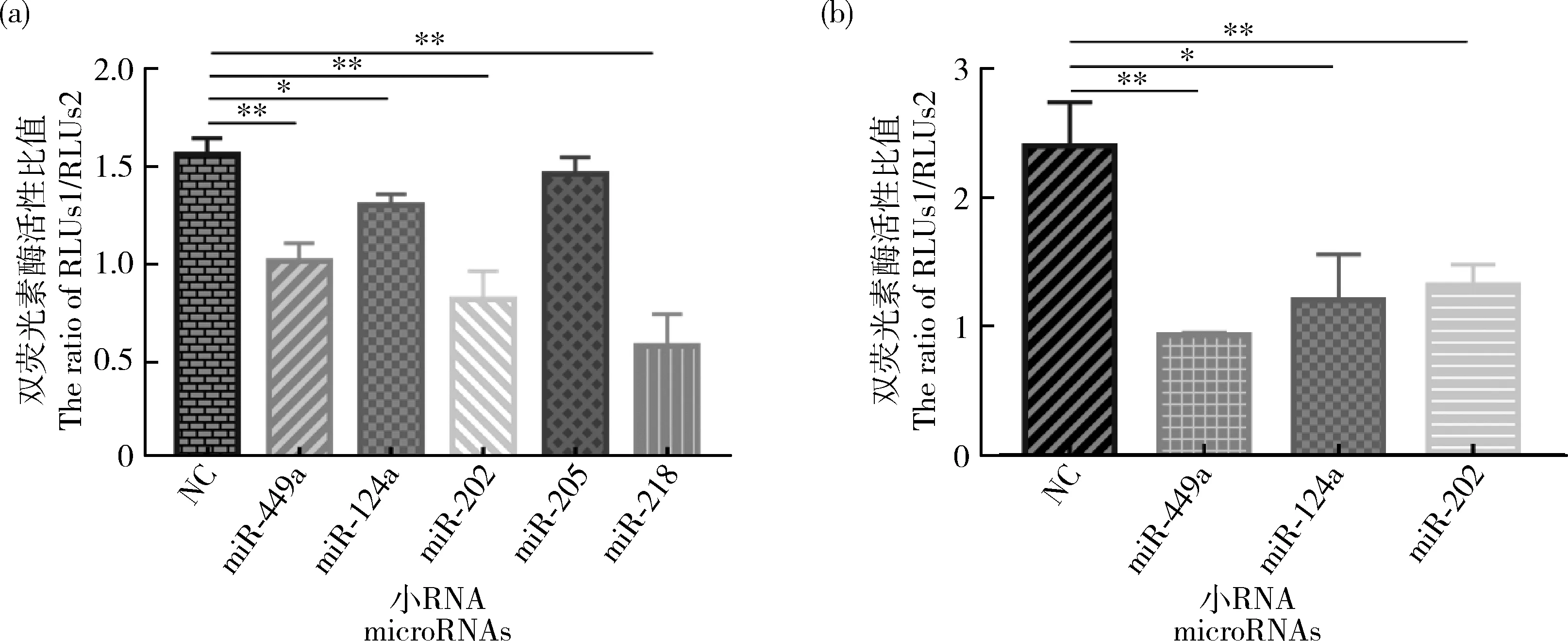
圖5 MicroRNAs與ACSL1-al(a)、ACSL1-as(b)3′UTR共轉染雙熒光素酶活性檢測
為了進一步確定miR-218是否是通過預測結合位點1 264~1 271 bp處與ACSL1
基因3′UTR結合,將ACSL1-
al 3′UTR表達載體中預測位點進行定點突變(圖6(a)),然后將突變后的載體與miR-218 mimics共轉染293T,突變后熒光素酶活性不再降低(圖6(b)),與轉染對照組熒光素酶活性幾乎相同,表明miR-218是通過該位點與ACSL1
3′UTR結合。綜上,結果顯示,ACSL1
-as 3′UTR具有較高的熒光素酶活性,并且更有助于ACSL1
的表達,同時miR-202、miR-449a、miR-124a和miR-218能夠與ACSL1
基因3′UTR結合,顯著降低細胞內(nèi)的熒光素酶活性。
圖6 miR-218預測結合位點(a)和ACSL1-al 3′UTR MUT與miR-218共轉染雙熒光素酶活性檢測(b)
為進一步確定在脂肪組織中miR-218對ACSL1
基因表達的影響,將miR-218 mimics和miR-218 inhibitor分別轉染至綿羊前體脂肪細胞中,檢測ACSL1 mRNA表達量和蛋白表達量。轉染miR-218 mimics后綿羊ACSL1
mRNA表達量顯著降低(圖7(a)),蛋白表達量顯著降低(圖7(b)和(c))。轉染miR-218 inhibitor后綿羊ACSL1
mRNA表達量顯著升高,蛋白表達量顯著升高。結果表明miR-218可通過3′UTR結合位點調(diào)節(jié)綿羊前體脂肪細胞中ACSL1表達量。
圖7 miR-218對ACSL1 mRNA表達量的影響(a)和miR-218對ACSL1蛋白表達量的影響(b)及灰度值檢測(c)
3 討 論
APA普遍存在于真核生物中,并且能夠調(diào)控基因表達,ACSL1
基因存在APA現(xiàn)象,有兩個不同長度的3′UTR,長度分別為1 570和748 bp。檢測ACSL1
3′UTR在誘導分化過程中表達量的變化,2種長度的3′UTR都是先上升后下降,與ACSL1
表達趨勢相同,并且ACSL1-
al 3′UTR表達量明顯高于ACSL1-
as,但是在誘導分化初期,ACSL1-
as 3′UTR的表達量上升速度顯著高于ACSL1-
al,這表明在綿羊前體脂肪細胞分化過程中,可能與T細胞被激活一樣,部分ACSL1
會切換成近端的ployA位點。研究表明,通過轉錄組分析脂肪干細胞脂肪生成,結果顯示半衰期的變化與3′UTR的延長有關,更長的UTR穩(wěn)定了主要的轉錄本。從而,這些延伸的長度可能與多核糖體結合能力的改變或半衰期有關。雖然從遠端到近端和近端到遠端的這類APA變化事件和剪接依賴性事件之間沒有顯著性差異,但有研究顯示T淋巴細胞活化的過程中86%的基因增加了短3′UTR亞型的相對表達量,表明這類3′UTR縮短與細胞增殖存在重要聯(lián)系。在雙熒光素酶活性檢測中,ACSL1-
as 3′UTR高于ACSL1-
al,并且連接不同長度3′UTR的ACSL1蛋白表達檢測中發(fā)現(xiàn)帶有短3′UTR的ACSL1蛋白表達量更高,說明短3′UTR更能促進ACSL1
基因表達并且在綿羊脂肪細胞誘導分化的初期更活躍。miR-202、miR-449a、miR-124a和miR-218均能顯著降低ACSL1
3′UTR活性,并且miR-218降低極顯著。在檢測誘導分化不同時microRNAs表達量的變化中,miR-218的表達是先下降后上升,與ACSL1
表達趨勢相符合,并且miR-218 的過表達可降低綿羊ACSL1
表達量。前期研究表明,綿羊ACSL1
基因過表達后脂肪細胞中甘油二酯含量和花生四烯酸含量顯著增加,因此miR-218可能通過ACSL1
基因表達影響細胞內(nèi)甘油二酯和花生四烯酸的含量,值得進一步探究。ACSL1
的APA現(xiàn)象已被證實在生物體中有特殊功能,有研究表明ACSL1
基因可能通過脂代謝調(diào)節(jié)結腸直腸癌,ACSL1
基因3′UTR多樣性可能作為治愈后復發(fā)可能性的標志,基因型為TT的患者較帶有一個C的患者更容易復發(fā)。這表明APA具有可以作為一種分子標記應用于各個領域的潛能。ACSL1
基因3′UTR被證明可與多個microRNAs結合。在小鼠的肝星狀細胞中miR-34a能夠與ACSL1
基因3′UTR結合,并靶向ACSL1
基因影響肝星狀細胞激活并引起肝纖維化。miR-34a作為哺乳動物脂質(zhì)代謝的轉錄調(diào)節(jié)因子,同樣在家禽的肝臟脂代謝中具有調(diào)節(jié)作用,在產(chǎn)蛋高峰期的母雞中,miR-34a-5p在肝臟中的表達明顯高于產(chǎn)蛋早期母雞,能夠調(diào)節(jié)肝細胞內(nèi)TAG和總膽固醇酯的合成。在甲狀腺癌的研究中,SNHG7在甲狀腺癌細胞系中的表達較高,SNHG7敲低后對ACSL1
基因有一定的抑制作用,研究發(fā)現(xiàn)SNHG7通過阻止miR-449a與ACSL1
基因結合來調(diào)控ACSL1
基因表達,進一步調(diào)控甲狀腺癌細胞的增殖。因為ACSL1
基因的3′UTR較長,許多microRNAs都可能與ACSL1
基因3′UTR結合,影響ACSL1
基因表達。綿羊ACSL1
基因有2個不同長度的3′UTR,其中較長的3′UTR為1 570 bp,可能存在很多與microRNAs結合的位點,并且有研究表明,帶有較長3′UTR的基因更穩(wěn)定和不容易被激活。因此綿羊帶有長3′UTR的ACSL1-
al這一轉錄本更穩(wěn)定,在綿羊前體脂肪細胞誘導分化的過程中更難被激活。可以利用ACSL1
基因這一特性為其相關的脂代謝等疾病提供新的靶標,為綿羊育種提供新的分子標記。4 結 論
綿羊ACSL1
基因因APA存在2個不同長度的3′UTR,ACSL1
-as在綿羊前體脂肪細胞誘導分化前期更活躍,并且短3′UTR更有助于ACSL1蛋白表達。miR-218通過與ACSL1
3′UTR 的1 264~1 271 bp處結合顯著降低ACSL1
基因表達。因此,APA和miR-218可調(diào)控綿羊ACSL1
表達,并可能通過ACSL1影響綿羊脂肪代謝。猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:36
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:34
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:50
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:48
