不同光周期對菊花生長發育及內源激素的影響
2022-01-10 07:10:58陸思宇楊再強張源達
華北農學報 2021年6期
陸思宇,楊再強,2,楊 立,張源達,鄭 涵
(1.南京信息工程大學,氣象災害預報預警與評估協同創新中心,江蘇 南京 210044;2.江蘇省農業氣象重點實驗室,江蘇 南京 210044)
隨著中國城市化進程的加快,切花需求量以每年20%的速度在快速增加,一年四季均有需求[1]。菊花作為世界四大切花品種之一,多年來在花卉市場上占有很大比重,但受長日照的限制,部分秋菊品種在春夏季節無法正常開花。光周期現象為植物通過感知日照長度的變化而控制植物開花的現象[2]。秋菊屬于短日植物,當日照長度小于其臨界日長時方才開花[3]。不同菊花品種的臨界日長不同,光周期誘導效應也不同。光周期誘導是指利用光周期對植物花期的調節作用,人為控制日長,進而控制植物的開花時間,實現反季開花[4]。目前,國內外已有關于光周期對菊花花期調控方面的研究,短日光照環境是秋菊開花的關鍵,通常認為暗期對菊花花芽分化的啟動起決定性作用,而光期通過光合作用影響植株的營養狀況,進而對開花數量產生影響[5]。光周期不僅影響菊花的成花誘導,作為光合作用的必要條件,直接影響著植株的營養生長和生理特性[6]。敖地秀等[7]研究發現,菊花在日長小于11 h的光照條件下開始花芽分化,延長至14 h則花期延遲,營養生長旺盛[8],而過分縮短日長則不利于菊花的生長。李海云等[9]發現,葉綠素含量隨日照時長的減少而減小,8 h菊花長勢較差,植株瘦弱,花卉質量下降[10]。汪菊淵等[11]發現,菊花從光周期處理到開花所需時間因品種而異,而與花型無關,10 h日長為46~87 d不等,神馬菊在9 h日長下花芽分化最快,歷時23 d[12]。持續的短日照處理可促使秋菊、寒菊提前開花,短日和長日交替則引發菊花毫無規律的花序分化,并出現花畸形[13],適宜光周期的中斷或打亂可顯著影響成花誘導過程及其規律性[14]。
光周期對花期的調控作用主要是通過紅光(Pr)、遠紅光((Pfr)2種光敏色素間的相互轉化,實現植物體內生理代謝的轉變過程[15]。當菊花受到持續性的短日光周期處理后,Pfr/Pr降低至臨界值,引發與促花反應相關的激素代謝[16],在激素的共同作用下形成開花刺激物傳遞至莖尖分生端,誘發一系列生理代謝促使花芽分化[17]。Torrey[18]認為,由光周期引發的這種激素類生長調節物的變化可影響作物根系對營養元素的吸收。植物接受光照時間長,有利于根系的生長,根系活力也較高,同時也有利于作物生長,葉片生長速率、地上部生長量和干物質量均較大[19]。隨著日長的縮短,菊花花期提前,但株高、莖粗和平均節間長度等減小,不同處理植株外觀形態有很大差異[20]。關于不同短日、長日光周期對菊花生長發育特性及其與內源激素、光合色素間的關系研究較少。姜貝貝等[21]發現,28~35 d是切花菊精云達到成熟的成花感受態的生長年齡,要獲得高品質切花,菊花前期的營養生長要達到35 d以上才能進行遮光處理。本研究通過設置系列梯度光周期處理,選取達到成花感受態的菊花苗,研究不同光照時長對菊花光合色素、生長形態,花序發育不同階段歷時及其內源激素、切花品質的影響,了解不同光周期的成花效應,找到菊花最適日長,為切花秋菊的反季栽培提供理論指導。
1 材料和方法
1.1 試驗材料
試驗于2020年7月17日在南京信息工程大學農業氣象試驗站日光溫室(118°42′E、32°12′N)內進行,供試菊花品種紅面為短日照植物,生長勢強,喜陽耐干,忌積澇,花瓣面呈紅色,屬于蓮座狀中輪花。于試驗前2020年6月10日將幼苗定植于口徑為25 cm的PVC盆中,栽培基質為微酸性土壤,每盆1株,陰涼處緩苗后移至向陽處長日照下生長,以備試驗使用。
1.2 試驗設計
試驗在罩黑膜的不透光棚(高1.5 m)中進行,采用定時開關棚的方法人工控制植株接受光照的時間。試驗共設5個處理,分別為晝長7(以下記為Ph7),8(記為Ph8),9(記為Ph9),10(記為Ph10),11 h(記為Ph11),并利用南京夏季自然光照長度作為CK(13~14 h)。其中Ph7光照時間為11:00-18:00,Ph8光照時間為10:00-18:00,Ph9光照時間為9:00-18:00,Ph10光照時間為8:00-18:00,Ph11光照時間為7:00-18:00,其余時間均處于黑暗不透光環境。于定植后的第37天即2020年7月17日,隨機選取暫未開始花芽分化的植株(40±2) cm,放入遮光棚的不同小區,每處理30株。不同處理除光照時長不同外,水肥管理措施均相同,盆土變干時將水澆透而盆內不汪水,不干則不澆;植株統一薄肥勤施,現蕾前主要施氮肥,孕蕾現花階段主要施磷鉀肥,后期生殖生長停止施肥。
7-8月所有處理光源均為自然光,9-11月由于自然光照時長縮短,自然光照外的時段均采用生長補光燈進行人工補光,補光燈光強為1 000 lx。冠層上方1.5 m太陽光合有效輻射由傳感器(WatchDog 2000,美國)自動采集,如圖1所示。
圖1 試驗期間光合有效輻射日變化Fig.1 Diurnal variation of photosynthetically active radiation during the experiment
1.3 測定指標與方法
1.3.1 生長指標及葉綠素含量的測定 于試驗后每30 d測定1次植株的株高、莖粗、葉片數和葉綠素,即30,60,90,120 d 4次測量,每個處理3次重復。
株高:由菊花植株基部至主莖生長點的自然高度;莖粗:距土面20 cm處的莖稈直徑;葉片數:葉長大于1 cm的可見葉片數;葉綠素含量:取中部成熟葉片0.2 g,參照李合生等[22]的方法在分光光度計(UV-1800,日本島津)中完成470,649,665 nm下的比色測定,并按下式分別計算葉綠素a(Chla)、葉綠素b(Chlb)、類胡蘿卜素(Car)以及葉綠素總含量(Chl)。
Chla=13.95D665-6.88D649
Chlb=24.96D649-7.32D665
Chl=Chla+Chlb
Car=(1 000D470-2.05Chla-114.8Chlb)/245
其中D470、D649、D665分別表示提取液在470,649,665 nm下的比色值。
1.3.2 發育期參考劉彩迎[23]的花期劃定方法 依據以下標準判斷并記錄下菊花植株達到現蕾(1/2以上植株出現花蕾)、破蕾(1/2以上植株花蕾頂端露白)、初綻(1/2以上植株舌狀花瓣形成,回心而抱)、初花(1/2以上植株外輪舌狀花瓣外展)、盛花(1/2以上植株舌狀花全展開,中部黃色雄蕊可見)以及初萎(1/2以上植株花序外2圈花瓣枯萎)的日期,并計算不同處理下菊花現蕾歷時、破蕾歷時、初綻歷時、初花歷時、盛花歷時以及初萎歷時。其中現蕾歷時為從試驗開始到植株現蕾的天數,破蕾歷時為從現蕾到破蕾的天數,初綻歷時為從破蕾到初綻的天數,初花歷時為從初綻到初花的天數,盛花歷時為從初花到盛花的天數,初萎歷時為從盛花到初萎的天數。
1.3.3 內源激素的測定 分別于以上6個發育期,取菊花中部成熟葉片1 g,用80%冷甲醇提取過夜,經多次萃取脫色、調節pH值和減壓蒸發后,過0.45 μm微孔濾膜,采用高效液相色譜法(HPLC)測定玉米素(ZT)、赤霉素(GA3)、生長素(IAA)和脫落酸(ABA)含量,具體參考劉英翠[24]的方法。
1.3.4 切花品質和根系活力的測定 于盛花期時測定并記錄單株花數、花徑(花序最大橫徑)、花瓣花青素含量以及根系活力。取1 g菊花花瓣,通過水浴振蕩快速提取菊花中的花青素,采用分光光度計法測定花青素含量,具體參考劉長姣等[25]的方法。根系活力的測定采用氯化三苯基四氮唑法(TTC法),無色的TTC被根系還原后生成紅色不溶于水的TTF,TTC的還原強度可表示根系脫氫酶活性,并作為根系活力指標[26]。
1.4 統計分析方法
采用SPSS 26對試驗所得數據進行ANOVA方差分析、Duncan多重比較(α=0.05),數據采用平均值±標準差表示。
2 結果與分析
2.1 光周期對菊花生長指標的影響
由圖2可知,菊花株高隨著發育期的推進均不斷增加。從第90天開始,CK株高均為最大,Ph11株高最小,差異顯著。從第60天開始,Ph7的莖粗始終最小,CK莖粗始終最大。Ph7、Ph8的莖粗隨著處理天數的增加始終在減小,這與植株所處的光照環境有關,植株需通過增加株高以在極有限的光照時間內捕獲更多光能,進而彌補光照時間的不足,受限于光照過短莖干變細、株莖比增加,植株徒長纖弱。試驗60 d開始,Ph7的株莖比始終最大,Ph11的株莖比最小,差異顯著。葉片作為植物重要的營養器官之一,其數目的多少直接受光照時間的影響。試驗30 d,Ph7、Ph8的葉片數已顯著小于其他處理組,Ph7葉片數僅為CK的53.3%,到120 d Ph7僅有CK的41.78%。整個試驗過程Ph7的葉片數始終最小,CK葉片數最大,差異顯著,Ph11與Ph9、Ph10的葉片數差異不顯著,次于CK。可見,光周期可直接影響植株的光形態建成,Ph7、Ph8由于光照時間過短,植株徒長高度而莖稈纖細,表現為極易倒伏,葉片小且少,長勢最差;長日照CK則生長最旺盛,株高、莖粗、葉片數均最大;Ph9、Ph10與Ph11差異未達顯著水平,與Ph11一樣,均具有矮壯粗的形態特征,植株體態矮化且勻稱。
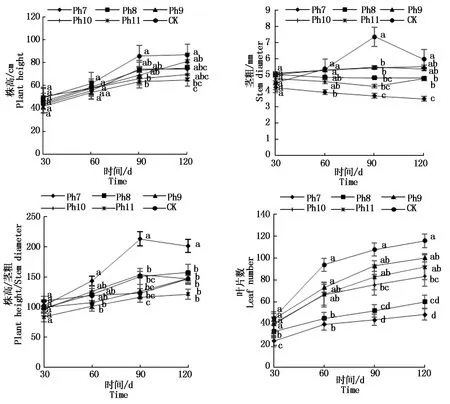
不同小寫字母表示不同光周期處理在0.05水平上的差異顯著性(P<0.05)。圖3,5-6同。Different lowercase letters indicate the significant difference at the level of 0.05 among different photoperiod treatments(P<0.05).The same as Fig.3,5-6.
2.2 光周期對菊花葉片葉綠素含量的影響
葉綠素為植物進行光合作用的主要捕光色素,與植物體內的代謝生長有關。菊花葉片內色素含量由高到低依次為葉綠素a、葉綠素b、類胡蘿卜素。由圖3可知,試驗第30天,以上3種色素均以Ph7的含量最高,葉綠素總量也最大,CK的光合色素含量最低,光合色素含量隨光照時長的增加而減少,其中CK的葉綠素a僅有Ph7的47.28%,類胡蘿卜素為Ph7的31.18%,差異顯著。60 d則以CK的類胡蘿卜素、葉綠素a、葉綠素b及葉綠素總量最高,Ph7的色素含量始終最低,光合色素含量隨光照時長的減少而減小,其中CK的類胡蘿卜素、葉綠素總量約為Ph7的2倍,差異顯著。90 d Ph9除外,光合色素含量隨著光照時長的減少而不斷減小,即以CK最大,Ph11次之,Ph7最小。120 d,隨著光照時長增加,葉綠素b、葉綠素總量均呈雙峰趨勢變化,第1峰值出現在Ph8,第2峰值出現在Ph11,且第2峰值均大于第1峰值,即Ph11光合色素含量最大,Ph7葉綠素含量最小,差異顯著。由此可見,光周期可顯著影響菊花葉片內光合色素含量,試驗初光合色素含量與光照時長呈反比,以極短日Ph7的色素含量最高,這可能是植物的一種抗逆應激反應,在光照時間嚴重不足時,植物在短時間內通過增加色素的合成以最大限度地捕獲光能進行光合作用,維持生長需要。光照可提高植物體內的葉綠素含量,隨著光周期試驗的推進,光合色素含量與光照時長呈正比。
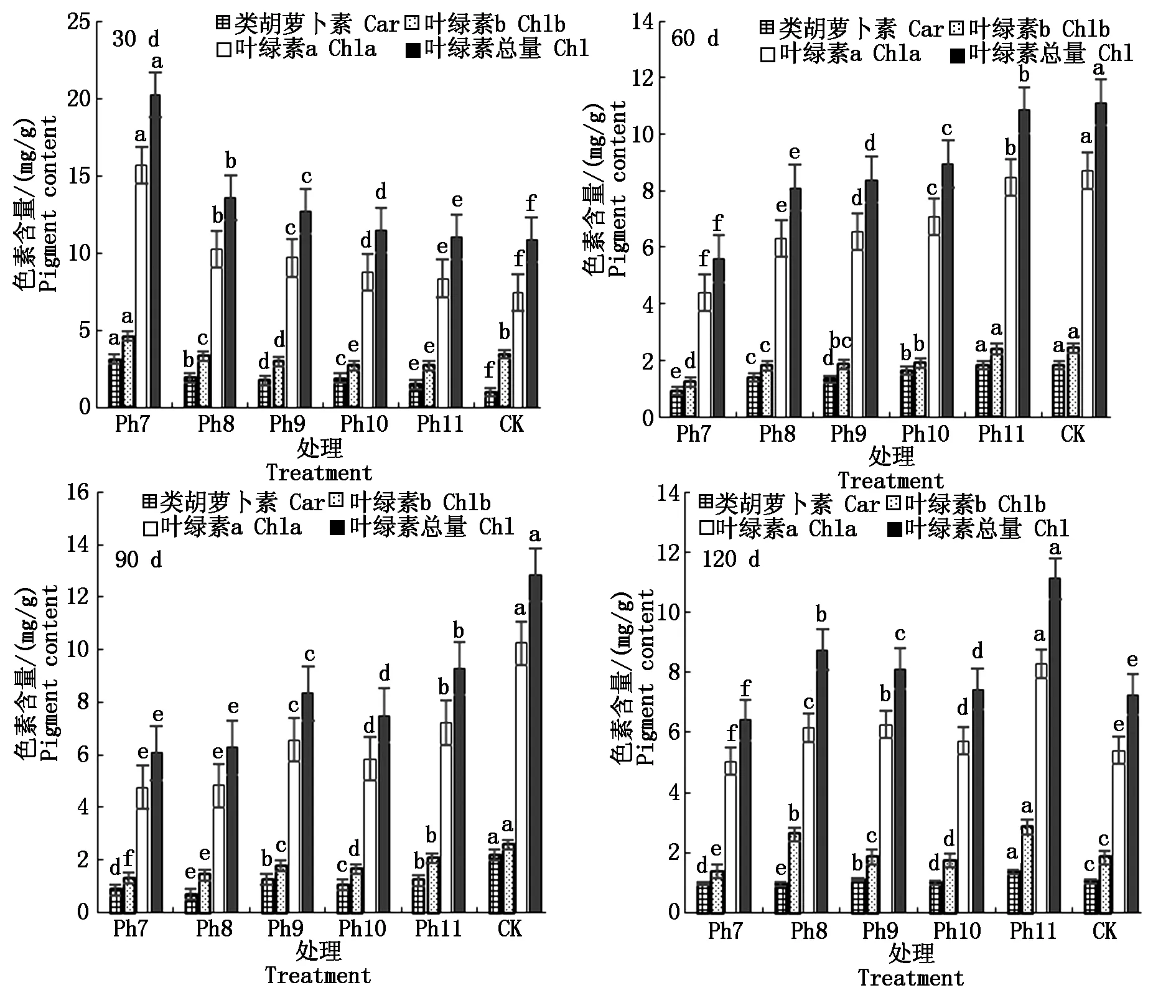
圖3 不同光周期處理下菊花光合色素的比較Fig.3 Comparison of photosynthetic pigments in chrysanthemum under different photoperiod treatments
2.3 光周期對菊花發育進程的影響
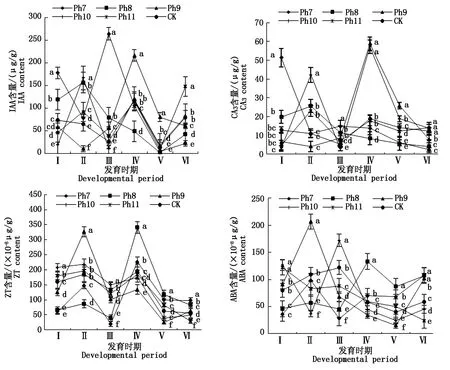
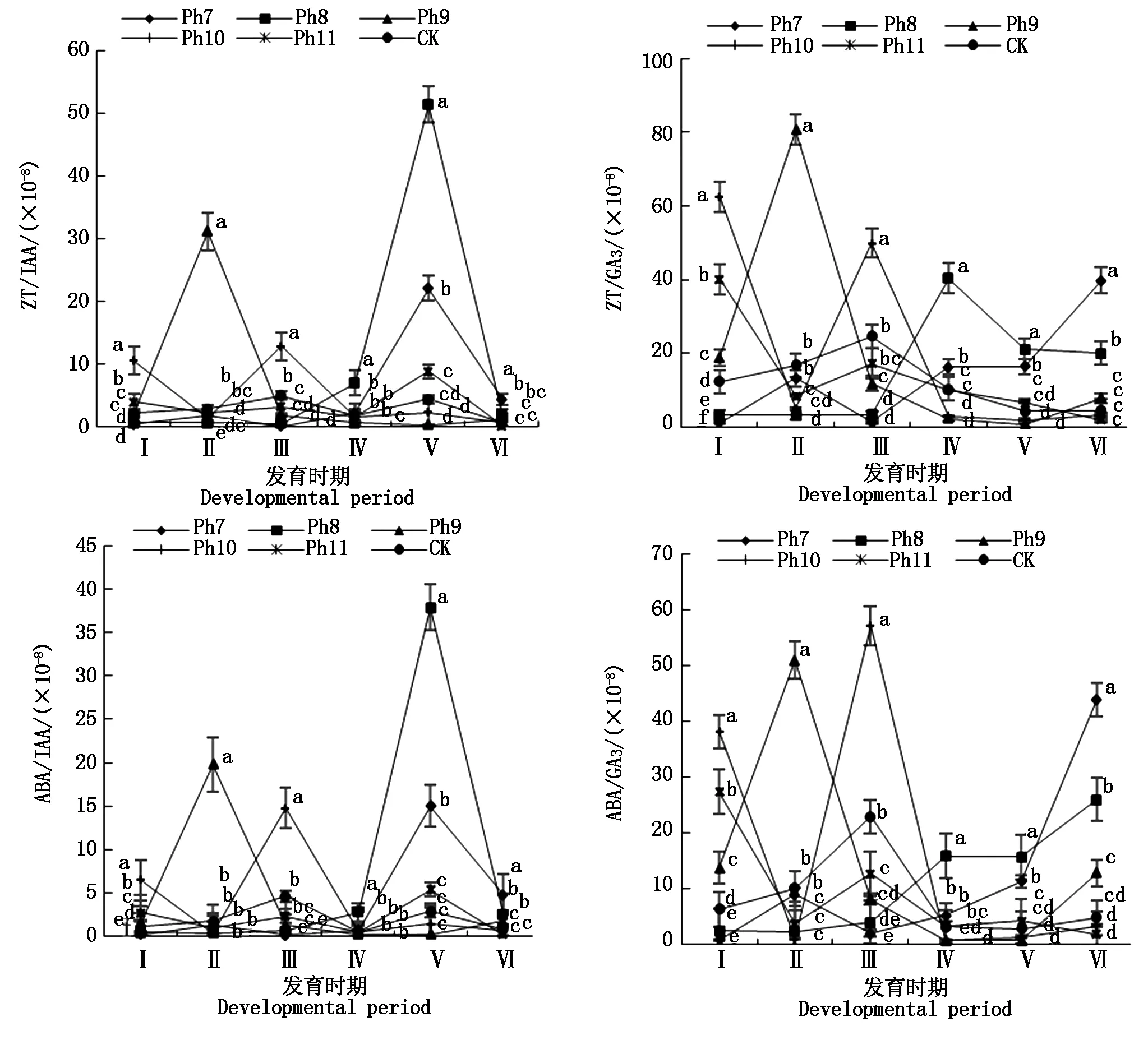
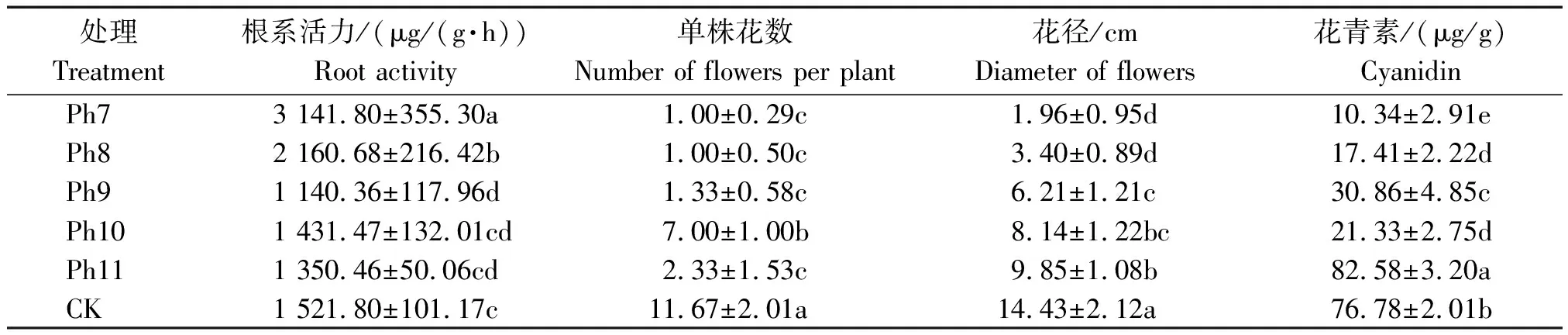
由圖4可知,不同光周期可明顯干擾菊花的花蕾發育進程,以現蕾歷時的組間差異最大。Ph10最先開始花芽分化,僅需35 d出現花蕾,Ph11次之,歷時39 d。Ph7從處理到現蕾歷時最長,105 d為Ph10的3倍,CK次之,歷時95 d,Ph8歷時78 d。可見,對菊花的成蕾過程起誘導作用的最適宜光周期為Ph10,誘導天數為35 d,Ph7與長日光周期下的菊花成蕾均嚴重滯后。不同光周期從處理到舌狀花完全外展的盛花歷時,總體表現為Ph10 圖4 不同光周期處理下菊花發育歷時的比較Fig.4 Comparison of chrysanthemum development duration under different photoperiod treatments 由圖5可知,光周期可顯著影響菊花葉片的內源激素含量,不同發育期下激素含量也不盡相同。菊花葉片IAA、GA3含量高于ZT、ABA。不同光周期的IAA和GA3含量在同一發育期的大小情況基本一致。現蕾期IAA、GA3均以Ph10含量最低,Ph7含量最高,Ph8次之,差異顯著;初綻期IAA、GA3以Ph10最低,Ph7最高,差異顯著;初花、盛花期IAA、GA3以Ph8最低,Ph9最高,差異顯著;Ph10的IAA含量次于Ph9,初萎期IAA、GA3以Ph7含量最低,Ph11含量最高,差異顯著,Ph10的IAA含量次于Ph11。隨著發育期的推進,ZT含量呈雙峰曲線變化,2次峰值出現在破蕾期和初花期,初綻、盛花期時ZT含量下降。破蕾期除外,不同光周期的ABA、ZT在同一發育期的大小情況基本一致。現蕾和初綻期ZT、ABA含量以Ph10最高,Ph7、Ph8較低,差異顯著;初花、盛花期Ph8含量最高,Ph7僅次之,Ph9、Ph10含量較低,差異顯著;初萎期依然以Ph7、Ph8含量較高,Ph11最低,Ph10僅次于Ph11,差異顯著。 Ph10在現蕾、初綻期,IAA、GA3含量最低,ZT、ABA含量最高,在初花、盛花以及初萎期IAA、GA3含量較高,ZT、ABA含量最低。Ph7、Ph8在現蕾、初綻期,IAA、GA3含量較高,ZT、ABA含量較低,初花、盛花以及初萎期IAA、GA3含量較低,ZT、ABA含量較高。可見,同一發育期內源激素含量的高低與光照周期有關,葉片內IAA、GA3與ZT、ABA存在著一定的拮抗作用。 Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ、Ⅵ分別表示現蕾期、破蕾期、初綻期、初花期、盛花期、初萎期。圖6同。Ⅰ,Ⅱ ,Ⅲ ,Ⅳ ,Ⅴ ,Ⅵ represent budding stage,bud breaking stage,petal formation stage,initial flowering stage,full blooming stage,early wilting stage,respectively.The same as Fig.6. 由圖6可知,不同光周期的4種激素比值在同一發育期的相對大小情況相同,現蕾期以Ph10比值最大,Ph11次之,Ph7最小;初綻期時以Ph10最大,Ph7、Ph8較小;初花、盛花期時均以Ph8最大,Ph9最小,Ph10次之;初萎期時以Ph7、Ph8比值較大,Ph11最小,Ph10次之,差異均顯著。 圖6 不同光周期處理下菊花內源激素比值的比較Fig.6 Comparison of endogenous hormone ratios in chrysanthemum under different photoperiod treatments 不同光周期ZT/IAA、ABA/IAA隨發育期變化的曲線走勢基本一致,ZT/GA3、ABA/GA3曲線走勢基本一致,這與菊花葉片內IAA、GA3含量大于ZT和ABA有關。Ph10現蕾期,ZT/IAA和ABA/IAA分別為最小值Ph7的30,31倍,初綻期Ph10的ZT/IAA和ABA/IAA快速增加,分別為最小值Ph7的170,140倍;Ph7、Ph8在盛花期,ZT/IAA和ABA/IAA快速增大,以Ph8的增加幅度最大,其中Ph8的ZT/IAA為最小值Ph9的170倍,差異顯著。對于ZT/GA3和ABA/ GA3而言,現蕾期Ph10遠遠大于其他處理組,ZT/GA3為最小值Ph7的52倍,ABA/GA3為Ph7的54倍;初綻期Ph10的ZT/GA3為最小值Ph7的37倍,ABA/ GA3為Ph7的30倍;初花期Ph8的ZT/GA3為最小值Ph9的18倍;初萎期時Ph7、Ph8的ZT/GA3和ABA/GA3快速增加,以Ph7的上升幅度最大,分別為最小值Ph11的19,26倍,差異顯著。可見,光周期可顯著影響不同發育期葉片內的激素平衡,并以Ph7、Ph8、Ph10的激素變化最大。 不同光周期下菊花根系活力差異顯著,Ph7、Ph8的根系活力均顯著大于CK,其中Ph7根系活力為CK的2倍,為最小值Ph9的2.7倍,差異顯著,這與Ph7、Ph8光照時間過短有關,根系活力的提高為植株抗逆性的表現。總體表現為Ph7>Ph8>CK>Ph10>Ph11>Ph9,Ph10、Ph11與Ph9根系活力差異不顯著。 不同光周期下的切花數量和切花質量有顯著差異。長日照CK的單株花數最多,其次為Ph10,Ph7、Ph8單株花數最少,均小于CK的10%,差異顯著。從表1可以看出,Ph7、Ph8的花徑和花青素最小。長日CK的花徑最大,Ph11次之,而花青素以Ph11最大,CK次之,可見,切花最大,花瓣不一定開得最鮮艷,同樣地,花瓣鮮艷程度也不一定與花徑呈正比。 表1 不同光周期處理下菊花根系活力和切花品質的比較Tab.1 Comparison of root activity and flower quality of chrysanthemum under different photoperiods CK由于光照充足,單株花數和花徑均最大,說明長日照有利于花徑和花數的提高,Ph10的單株花數和花徑較大,花青素含量較低,Ph7、Ph8不僅切花數量少且小,切花品質也最低。 光作為植物最重要的能量來源,對植物的營養積累與生長發育過程有著深刻的影響[27]。有研究表明,光周期能誘導植物體內與營養生長相關的基因表達[28],延長光照時間可增大油葵芽苗菜的植株高度和葉面積[29],增大黃瓜的莖粗并加快其生長速率[30]。在本試驗中,不同光周期下菊花的外觀形態存在差異顯著,長日照CK由于光合時間最長,葉綠素合成增加至最大,生長最旺盛,植株高大粗壯、葉片濃密。光周期的延長使植株進行光合作用的時間增加,合成積累更多有機物以支持植物的生長發育,反之光周期縮短則有機物合成減少,生長受阻[31]。極短日光周期Ph7、Ph8的莖粗試驗過程始終在減小,維持株高的不斷增加,說明此光照條件已不能滿足正常生長需要,植株受到光脅迫,需通過增加株高以捕獲到更多光能,屬于植物的一種抗逆性表現,其根系活力和葉綠素含量更是說明了這一點,Ph7、Ph8根系活力遠大于CK,且試驗初期葉綠素含量最大,這均為植株在不利環境下的應激表現,隨著不利環境的持續,光合色素含量變為最小,葉片數最少,長勢最差,極大的株莖比導致植株易倒伏,這與李海云等[32]的研究結果一致。Ph10與Ph11生長指標差異不顯著,介于CK和Ph7、Ph8之間,均具有矮壯粗的形態特征,Ph11葉綠素含量則顯著大于Ph10,由此可見,光周期可顯著影響葉片內的光合色素含量,這與薛歡等[33]的研究結果一致。 對于短日植物秋菊而言,光周期是決定其花芽分化與否的重要環境因子。本研究表明,不同光周期下菊花的小花發育差異較大,Ph10最先開始花芽分化,僅需35 d即完成花蕾的發育過程,在長日照、極短日光周期Ph7、Ph8條件下,植株在很長一段時間內都處于營養生長狀態,花期嚴重推遲,成蕾誘導天數長達105 d,這說明菊花對Ph10的光照環境(長暗期)較敏感,花芽分化的啟動需經過一定長度的暗期,且開花響應不是簡單地與暗期長度呈正比,這與房偉民等[34]的研究結果一致。植物的花芽分化和小花發育是一個受多物質調控的復雜過程,葉片內源激素就是其中一個關鍵因子,作物花芽發育進程與內源激素含量、激素平衡息息相關。IAA、GA3被認為是抑制小花發育的一種激素信號,對于短日植物而言,高含量的IAA、GA3抑制花芽分化[35]。林貴玉等[36]研究認為,高含量的ZT、ABA有利于短日植物的花芽分化。本研究表明,現蕾期時,Ph10的IAA、GA3含量最低,ZT、ABA含量最高,而CK和Ph7、Ph8的IAA、GA3含量較高,ZT、ABA含量較低,這與前人的研究結果一致。現蕾期時Ph10的ZT/IAA、ABA/IAA、ZT/GA3和ABA/GA3均較大,CK和Ph7、Ph8的激素比值較小更是說明了這一點,ZT/IAA、ABA/IAA、ZT/GA3以及ABA/GA3越大,越有利于菊花的花芽分化。 王彩俠[37]研究認為,菊花在破蕾顯色后,花期將不再受光周期的影響。本研究表明,在初花和盛花期,均以Ph7、Ph8的IAA、GA3含量較低,ZT、ABA較高,ZT/IAA、ABA/IAA、ZT/GA3和ABA/ GA3較大,對應的初花、盛花發育歷時較小,而從處理到盛花依舊是以Ph10的歷時最短,Ph7、Ph8歷時較長,說明光周期的促花作用主要是影響花芽分化啟動、花蕾形成過程而達到花期調控的目的,舌狀花瓣的展開過程雖然也受光周期的調節,但對花期的提前不起決定作用,這與前人研究結果基本一致。初萎歷時即盛花持續時間以Ph10較長,且大于CK,這與植株體內ZT、ABA含量較低,IAA、GA3含量較高有關。以Ph7、Ph8的盛花持續時間最短。 營養生長是生殖生長的基礎,開花質量與植株前期的營養生長狀況息息相關。本研究表明,長日照CK菊花由于光合色素含量高,生長旺盛,單株花數和花徑均最大,Ph10次之;Ph10植株體態矮化、勻稱,生長充分,在高含量的促花激素ZT、ABA作用下,最先啟動花芽分化出現花蕾,CK以抑花激素IAA、GA3含量較高而促花激素含量較低,植株保持過于旺盛的營養生長,花期滯后。Ph7、Ph8根系活力較大且在試驗初出現光合色素遠大于其余處理組的抗逆反應,光照時長過短植株已受到光脅迫,試驗后期色素含量低、植株徒長纖弱,極度匱乏的營養生長以及體內含量極低的促花激素導致生殖階段切花小且少、現蕾歷時最長,切花質量最低,這與朱玲俐[38]的研究結果一致。 花青素表征著花瓣的鮮艷程度,使花瓣呈現出紅色、紫色、藍色[39]。Ph9、Ph11和CK的花期雖都晚于Ph10,但花青素含量卻顯著大于Ph10,光周期在促花的同時,也會對花色和切花品質產生影響[40]。Ph9花瓣色澤鮮艷,但花心外露植株出現畸形花,這可能與管狀花發育不良有關。張翠華[41]研究表明,花心外露大多為光照時長過短造成,光周期下畸形花的形成機理還有待進一步研究。 綜上所述,Ph10即10 h/14 h(晝/夜)對菊花開花的誘導效果最好,植株體態矮化且勻稱,花芽分化早,花蕾發育周期短,花期集中,盛花產出快且不易萎蔫。生產上為獲得高品質切花菊,對長日照下生長35 d以上處于成花感受態的菊花進行10 h/14 h光周期處理,可使菊花最快啟動花芽分化,待破蕾顯色后移至長日照下正常開花。此結論可為夏季南京地區的菊花栽培提供理論依據,實現秋菊的反季開花。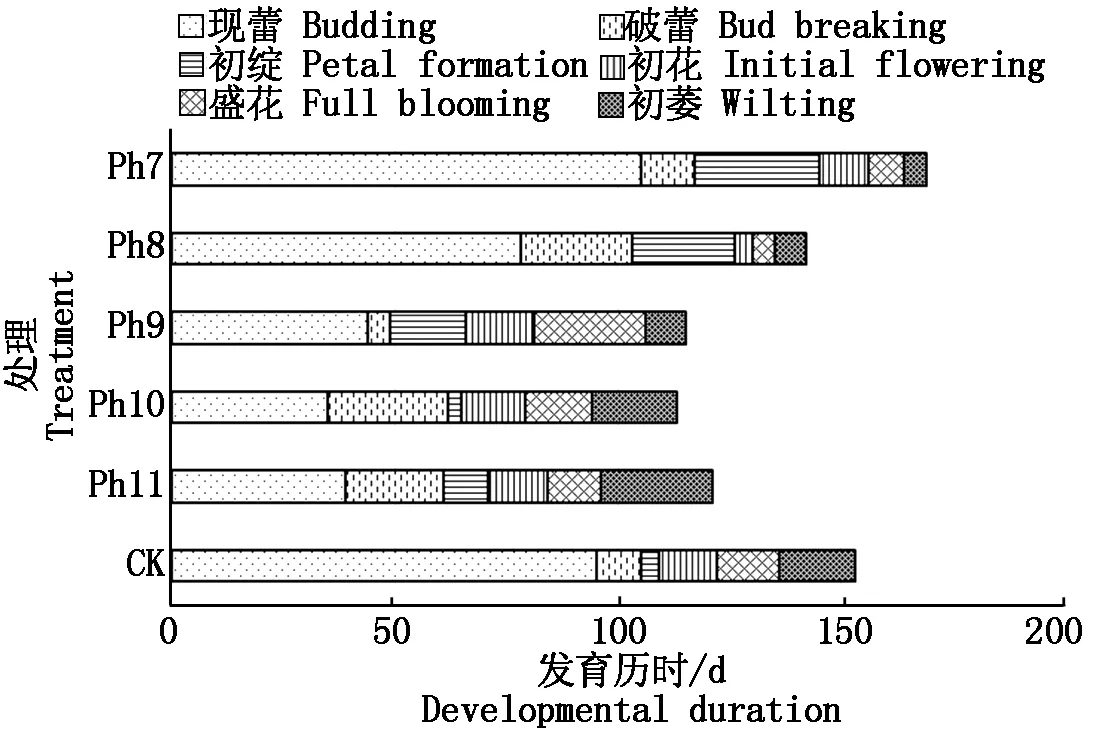
2.4 光周期對菊花葉片內源激素含量的影響
2.5 光周期對菊花葉片內源激素比值的影響
2.6 光周期對菊花根系活力和切花品質的影響
3 討論與結論
3.1 光周期對菊花營養生長的影響
3.2 光周期對菊花生殖生長的影響
3.3 光周期對菊花生長發育的影響
