基于耳石微化學的黃河墾利段刀鱭生活史初步研究*
2022-01-14 07:48:04叢旭日李秀啟董貫倉王亞楠孫魯峰
漁業科學進展 2022年1期
叢旭日 李秀啟 ① 董貫倉 王亞楠 孫魯峰 客 涵 楊 健 姜 濤
(1. 山東省淡水漁業研究院 山東省淡水水產遺傳育種重點實驗室 山東 濟南 250013;2. 中國水產科學研究院淡水漁業研究中心 中國水產科學研究院長江中下游漁業生態環境評價與資源養護重點實驗室 江蘇 無錫 214081)
刀鱭(Coilianasus)是一種極為名貴的洄游性經濟魚類。20世紀60年代黃河刀鱭年產量可達1500 t (蔡德霖等, 1980),是黃河下游的重要物種,但受過度捕撈、水利工程、黃河斷流等影響,山東東平湖產卵場已被嚴重破壞,黃河口及河段很長時間內已無刀鱭分布。近年來,根據山東省淡水漁業研究院對黃河口漁獲物調查數據及對當地漁民的走訪發現,黃河刀鱭再一次在黃河下游出現,但資源量依然較低。隨著刀鱭經濟價值的提高,相關針對資源保護的科研工作正在開展,但多數集中于長江河段。近日,黃河流域生態保護和高質量發展已上升為國家戰略,漁業資源的合理開發和保護顯得尤為重要,但黃河河段刀鱭研究依然空白,相關工作嚴重滯后。
耳石微化學分析能有效地反演洄游性魚類的“履歷”,在耳石中,Sr、Ca元素沉積變化被用作研究不同鹽度海、淡水生境條件的“指紋”(竇碩增等, 2011;楊健等, 2010;姜濤,2014;姜濤等,2016),結果具有獨特的生境相關性及客觀性的優勢,即使僅有1尾樣本也能有效把握其生境狀態(徐鋼春等, 2014)。姜濤等(2013)通過分析鄱陽湖刀鱭耳石微化學特征發現,鄱陽湖內存在2種不同生活史的刀鱭群體;盧明杰等(2015)研究大遼河口紅狼牙虎魚(Odontamblyopus rubicundus)生活史類型后發現,其可以適應廣鹽度水體。近幾年來,該技術也已經應用到大水面放流效果評價中。李秀啟等(2017)驗證了耳石鍶標記技術在識別鳙(Aristichthy snobilis)放流個體的可行性并提出了最佳標記濃度;司飛等(2019)探究了耳石微化學技術在牙鲆(Paralichthysolivaceus)增殖放流過程中的應用。為更好地保護黃河口刀鱭,本研究依托EPMA技術,對黃河刀鱭進行耳石微化學分析,以探究其生活史,為刀鱭種質資源的保護、資源量的提高、漁業資源的可持續發展提供可行性建議。
1 材料與方法
1.1 實驗材料
刀鱭于2016年5月采自黃河墾利段(圖1),主要捕撈網具為刺網。共取得刀鱭 5尾(表1),捕獲后置于-20℃冷凍待用。解剖后取出矢耳石,為降低誤差,統一使用左矢耳石進行分析,使用去離子水清洗耳石表面雜質及薄膜,置于室溫晾干待用。
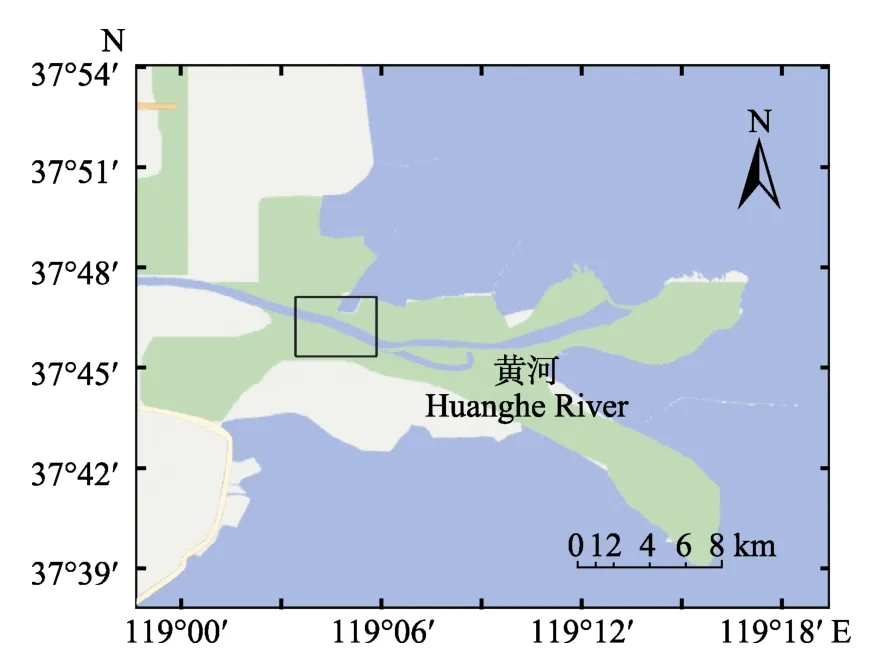
圖1 刀鱭采樣地點Fig.1 The sampling sites of C. nasus
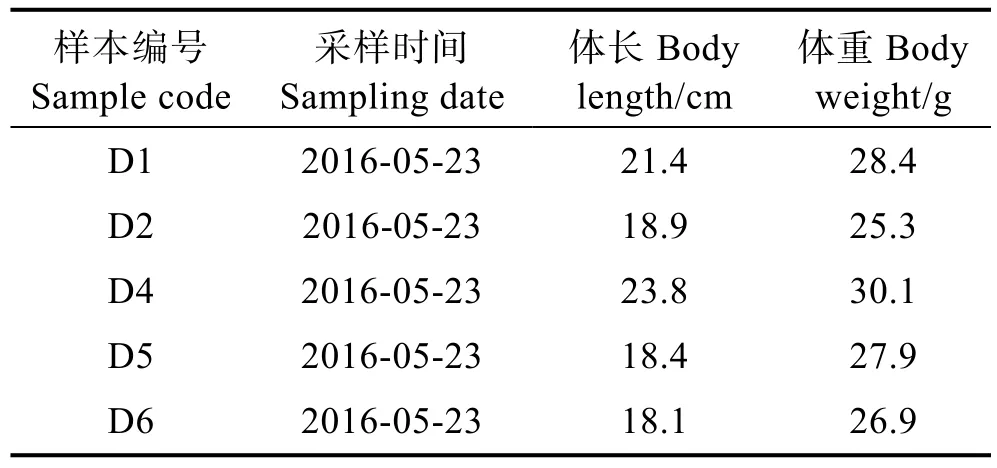
表1 刀鱭采樣資料Tab.1 Sample details of C.nasus from the Kenli section of the Huanghe River
耳石樣品使用環氧樹脂包埋固定制成樹脂塊,然后將樹脂塊固定于載破片上,凝固后切去多余樹脂部分。使用Discoplan-TS研磨機進行粗磨至耳石剛要暴露,再用1200目的砂紙精磨至耳石核心完全暴露,最后采用Roto Pol-35磨拋機配合拋光液拋光至耳石表面無劃痕。處理好的耳石置于Milli-Q超純水中超聲清洗5 min,于烘箱內38℃過夜烘干。
1.2 耳石微化學分析
參考Yang等(2006)的方法,使用X射線EPMA電子探針微區分析儀(JXA-8100,日本)分析耳石樣品。自耳石核心沿最長徑至耳石邊緣的一條直線進行定量線分析(line transect analysis)。EPMA 加速電壓和電子束電流分別為 15 kV 和 2×10-8A,束斑直徑為 5 μm,每點駐留時間為15 s,以10 μm為間隔連續打點測定。標準樣品取用碳酸鈣(CaCO3)和鈦酸鍶(SrTiO3)。定量線分析完成后,再對耳石樣品進行面分布分析(mapping analysis)。此時,EPMA加速電壓和電子束電流分別為 15 kV 和 5×10-7A,束斑直徑為 5 μm,像素為 7 μm×7 μm,每點駐留時間為 30 ms。
1.3 數據處理
使用Excel 2010軟件進行數據處理,使用SPSS 17.0軟件進行非參數檢驗(Mann-Whitney U-test)。引入進行格局轉變(Regime shifts)分析的STARS方法(Sequential T-test Analysis of Regime Shifts),當多于1個連續點出現顯著變化(基于數據方差和t檢驗的P值),將視為“轉變”,生成新的移動平均(Rodionov,2004; Rodionovet al, 2005)。本研究設定截斷長度為10,Huber權重為1,置信度P為0.1,按照慣例Sr/Ca值標準化為Sr/Ca×103。
2 結果
2.1 定量線分析比較
定量線分析結果顯示(表2),每尾刀鱭生長階段不盡相同。本研究中,根據Sr/Ca值可分為不同生活史階段,且相鄰不同階段差異顯著(P<0.05, Mann-Whitney U-test)(表2)。個體D1的Sr/Ca值可分為4個階段,分別為從核心沿矢狀面像邊緣至240 μm耳石徑的低值區域,平均值為 3.63±0.61;241~1320 μm 區域,平均值為2.24±0.72;1321~1960 μm 區域,平均值為3.57±0.68;1961~2310 μm 區域,平均值為 4.90±0.69。個體D4可分為3個階段,0~1180 μm區域,平均值為 1.90±0.76;1181~1570 μm 區域 , 平均值為3.83±0.74;1571~2190 μm 區域,平均值為 4.82±0.77。個體D5可分為3個階段,0~1080 μm區域,平均值為 2.15±0.78;1081~1520 μm 區域,平均值為5.61±0.75;1521~1930 μm 區域,平均值為 4.74±0.70。個體D6可分為3個階段,0~660 μm區域,平均值為2.13±0.77; 661~1120 μm 區域,平均值為 6.17±0.99;1121~1440 μm區域,平均值為 4.84±0.76。圖 2以折線圖的形式表現不同類型黃河刀鱭與東平湖刀鱭耳石矢狀面從核心(0 μm)到邊緣定量線分析Sr/Ca值變化對比。個體D2線分析顯示,生活史無規律,故不做階段劃分。
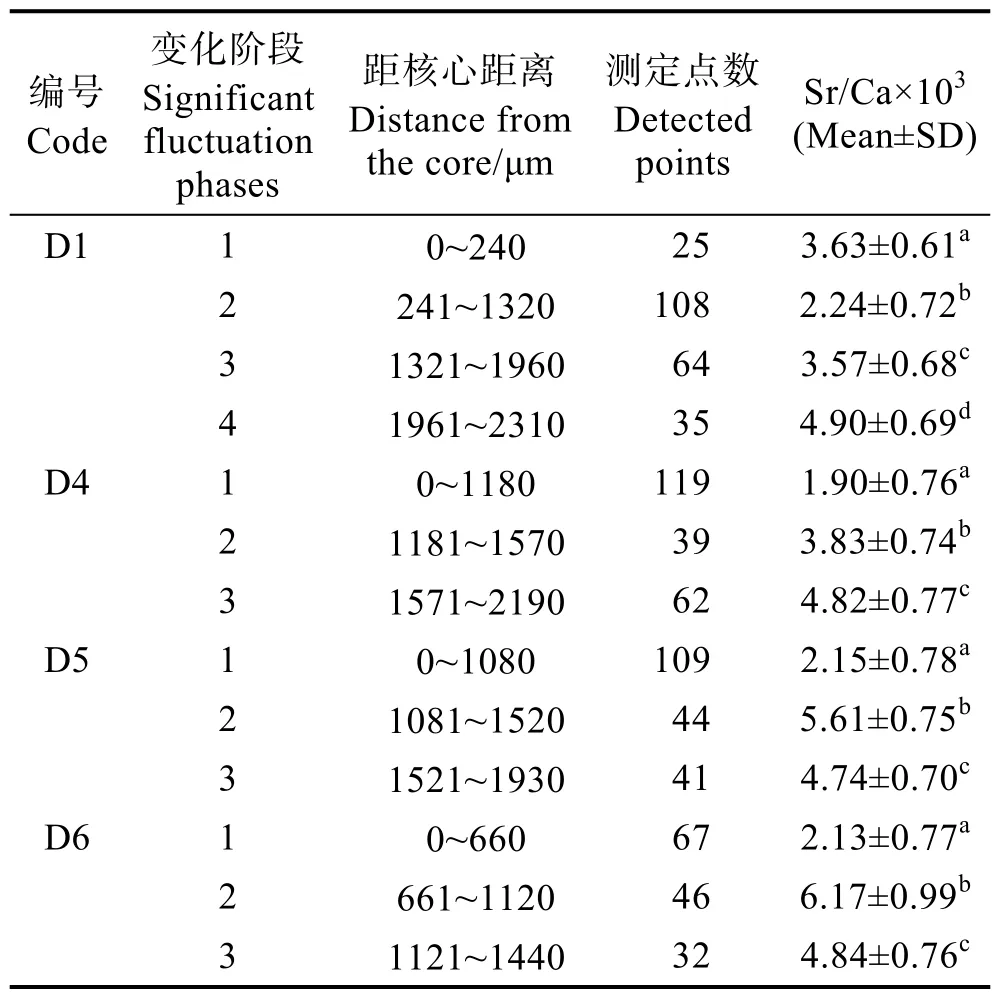
表2 刀鱭耳石鍶鈣比微化學Tab.2 Fluctuation of Sr and Ca microchemistry in otoliths of C.nasus
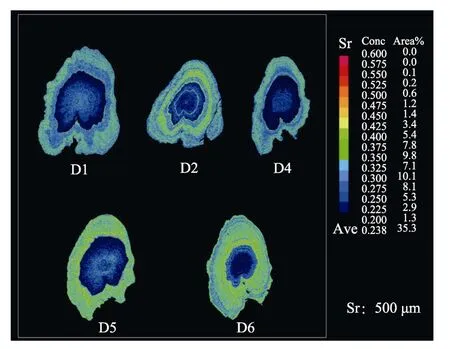
圖2 耳石矢狀面二維Sr含量分布Fig.2 Two-dimensional imaging using mapping analysis of the Sr in otoliths of C.nasusSr:鍶;Conc:Sr濃度;Area/%:面積占比;Ave:Sr濃度表平均值Sr:Strontium; Conc: Concentration of Sr;Area/%: The area proportion;Ave: The average concentration of Sr
2.2 面分析比較
面分析的結果更能直觀清晰地顯示出黃河刀鱭個體的生境履歷。Yang等(2011)關于長江刀鱭的研究表明,刀鱭對應淡水、淡咸水、海水生境的Sr/Ca有十分明顯的微化學特征,具體表現為<3(藍色)、3~7(黃綠色)、>7(黃紅色),Ⅲ類水體中,Sr/Ca×103值平均為海水區8.3±4.5、河口區5.6±1.1、淡水區2.7±1.5(Sectoret al, 2000; Yanget al, 2011)。個體 D2核心區域表現為藍色,隨著個體的生長,黃色、藍色交替出現;個體D1、D4核心區表現為藍色,外緣區域表現為藍綠色;個體D5、D6核心區域表現為Sr值較低的藍色區域,隨著刀鱭生長,耳石外圍逐漸形成較為明顯的黃色圓環,表明其已經進入Sr值較高的生活環境。
3 討論
耳石中元素Sr和Ca濃度比值的變化在國內外研究中已經被廣泛應用于魚類生活史中(Wellset al, 2000;Gillanderset al, 2005; Araiet al, 2006; 陳婷婷等, 2016;熊瑛等, 2014; 叢旭日等, 2019),但有關黃河刀鱭的研究尚未見報道。蔡德霖等(1980)研究表明,黃河刀鱭的生活史特征較為單一,成魚每年 4—5月沿黃河溯河洄游至東平湖,6—7月產卵,等孵化后于次年初進入黃河口海域。由此可見,歷史上黃河墾利段是刀鱭重要的洄游通道,東平湖是主要產卵場,黃河口海域為索餌場、越冬場。對于現階段黃河刀鱭是否為洄游型刀鱭的爭論較大,有的學者認為,黃河墾利段已不存在洄游型刀鱭,其再次出現是由于東平湖內定居型刀鱭被動進入黃河所致,但此觀點缺乏充足論據。本研究從耳石微化學的角度出發,獲得的結果客觀真實地反映出黃河墾利段刀鱭的生境利用情況,為證明“三場一通道”依然分布洄游型刀鱭及其生活史多樣性提供了理論基礎。
本研究結果顯示,黃河墾利段刀鱭的生境履歷具有復雜性的特點。Brown等(2009)研究表明,海水的Sr/Ca×103值相對穩定(8.17~8.87),而河流(0.27~19.18)和淡水湖泊波動較大(0.20~5.02)。然而,孫珊等(2019)和徐江玲(2007)研究表明,受調水、調沙的影響,黃河口區域鹽度的時空變化較為劇烈,調水、調沙前后表層鹽度變化范圍約為6~25,垂直方向亦波動較大,故不能通過傳統研究方法判斷黃河刀鱭是否到達鹽度較高的海水區域。本研究的個體可分為2種類型。個體D2屬于第Ⅰ類型,其生活史較復雜,淡水孵化后其生長軌跡多次往返于淡水—半咸水生境,格局發生多次轉變,不具備洄游型刀鱭應有的規律性特征,此外,耳石邊緣Sr/Ca值反映樣品被捕獲時的環境條件,個體 D2耳石邊緣 Sr/Ca值(<3)較其他個體低,這是因為Sr/Ca值變化存在時滯效應,表明個體 D2進入此水域的時間較短,耳石尚未響應生境的變化。大麻哈魚(Oncorhynchusketa)耳石的此種效應可持續 12 d (王臣等, 2015);李孟孟等(2017)對長江安慶江段刀鱭的研究也存在此種效應。個體 D1、D4、D5和D6屬于第Ⅱ類型,孵化和早期的生活均在鹽度較低的淡水中進行,淡水生境履歷較長。線分析顯示,個體D1和D4能達到的Sr/Ca最大值分別為6.50和6.29,并未達到海水區的平均值(8.3)。D5和D6個體Sr/Ca的最大值雖然達到8.44和8.63,但由于黃河口區域鹽度二維變化受調水、調沙影響較大,尚不能判斷其生境履歷已延伸到鹽度較高的海水區。此外,個體D5雖然進入Sr/Ca值較高的區域生活,但停留時間較為短暫。STARS法計算顯示,個體D5沒有形成“轉變”,而個體D6形成了“轉變”(圖3)。
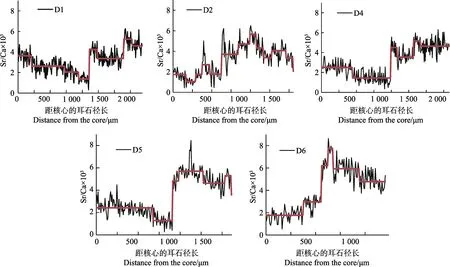
圖3 不同類型刀鱭耳石矢狀面從核心(0 μm)到邊緣定量線分析Sr/Ca值的變化Fig.3 Fluctuation of otolith Sr/Ca concentration ratios along line transects from the core (0 μm) to the edge in otoliths of C. nasus
通過EPMA技術分析顯示,本研究首次直觀地了解黃河口刀鱭的生境履歷特征。以個體D1、D4、D5和D6為代表的群體孵化、育幼階段均在淡水中進行,索餌階段進入Sr/Ca值較高的河口區,個體的Sr/Ca值均有一個從低值階段(<3)突然升高至高值階段(3~7)的變化,根據對長江刀鱭多年研究,第Ⅰ階段到第Ⅱ階段突越性的變化(從淡水進入半咸水生活)對洄游性刀鱭來說是至關重要的(姜濤等, 2013),個體D1、D4、D5和D6變化劇烈,都存在突越性變化,表明其主動進入鹽度較高的區域,符合洄游型刀鱭的特征。以個體D2為代表的群體生活史較為復雜,無規律,推測可能是由于當年東平湖汛期放水而被動進入黃河的定居型刀鱭,其生活史軌跡受黃河水流的影響較大,雖然該個體無洄游型刀鱭的明顯特征,但據徐鋼春等(2014)、李孟孟等(2017)對于長江刀鱭的研究表明,長頜鱭(Coilianasus)與短頜鱭(Coilia brachygnathus)均參與江海洄游,長江安徽和縣江段的短頜鱭也存在洄游個體,而短頜鱭屬于湖泊定居型刀鱭,分子層面上基于Cyt b和D-loop序列的分子標記亦不能區分洄游型刀鱭與定居型刀鱭(孫超等, 2018),表明刀鱭具有較強的環境適應性,也表明汛期從東平湖進入黃河的定居型刀鱭經過自然演化,一旦具備條件也會進行溯河洄游。此外,個體D1和D5核心區域的 Sr/Ca×103值較高(>3),D2、D4和 D6核心區域的 Sr/Ca×103值較低(<3),這也在某種程度上反映了存在個體起源于不同產卵場的可能性。姜濤(2014)研究發現,耳石的長度與魚體的年齡有非常高的相關性,可以作為估算刀鱭生活史的時間尺度,姜濤(2014)的研究還表明,黃河刀鱭大約在1齡離開淡水生活[區域為(980±111) μm]。本研究中,個體D1(1210 μm)、D4(1160 μm)和 D5(1080 μm)均符合這一特征,均在1齡或1齡+離開淡水生活,但個體 D2(590 μm)和 D6(560 μm)在不到1齡就已經離開淡水生活。其中,推測個體D2受黃河水流的影響,尚未具備洄游特征,而個體D6顯示出黃河刀鱭可能來自不同的群體,也表明其較強的環境適應性,與Jiang等(2014)在2009年捕獲的10尾黃河刀鱭耳石微化學分析結果相似,視其來源于同一群體。但在本研究中,黃河刀鱭個體表現出不同的生活史特征,2013年南水北調東線正式通水至今,東平湖定居型刀鱭已成為當地主要經濟性魚類,推測東平湖與黃河聯通河道不定期泄洪致使一部分定居型刀鱭進入黃河,是黃河刀鱭的資源群體組成較為復雜、多樣的重要原因。
隨著黃河流域生態保護和高質量發展國家戰略的提出,黃河流域漁業資源的可持續發展變得尤為重要。近年來,由于產卵場被破壞、閘壩工程的興建、過度捕撈等因素,黃河刀鱭的種群結構及生活史演變較大。根據調查,黃河與東平湖之間的聯通河道大清河只有在每年夏季泄洪時才開閘放水,目前,尚不能證明東平湖具備產卵場的條件,在今后的工作中,應進一步對早期發育階段的刀鱭進行基礎生物學及耳石微化學的分析,結合年齡結構、生長死亡參數及親體產卵狀況更為精確的掌握東平湖、黃河下游及河口地區刀鱭生活史,監測其是否形成固定的洄游種群,從而驗證歷史上東平湖、黃河山東段、黃河口是刀鱭洄游的“三場一通道”。相關部門也可以利用已有先例的河壩魚道技術,保護洄游通道及產卵場的生態功能,確保種群正常的繁衍。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
今日農業(2021年9期)2021-11-26 07:41:24
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
