TGF-β1受體對(duì)ClC-3氯通道在正畸骨代謝中表達(dá)的影響
2022-01-17 13:18:44盧曉琳金作林
口腔醫(yī)學(xué) 2021年12期
盧曉琳,王 歡,曹 猛,金作林
牙槽骨改建是維持顱頜面骨骼正常形態(tài)結(jié)構(gòu)的重要環(huán)節(jié),也是正畸牙齒移動(dòng)的生物學(xué)基礎(chǔ)。牙槽骨在正畸力的作用下處于骨吸收和骨形成的動(dòng)態(tài)平衡過(guò)程,壓力側(cè)破骨細(xì)胞分化成熟,促進(jìn)骨吸收;張力側(cè)成骨細(xì)胞增殖分化,增強(qiáng)新骨形成[1]。但是關(guān)于牙槽骨代謝的具體作用機(jī)制目前尚不明確。ClC型氯通道是一種電壓門控型氯通道。其中,ClC-3作為電壓門控型氯通道之一,存在于細(xì)胞膜或細(xì)胞器膜上,參與調(diào)節(jié)細(xì)胞容積、細(xì)胞遷移、增殖與凋亡以及細(xì)胞器酸化等多種生物活動(dòng)[2-3]。而且,ClC-3氯通道在骨代謝方面也發(fā)揮著重要的調(diào)節(jié)作用,其基因敲除小鼠自出生后就表現(xiàn)出發(fā)育遲緩、鈣磷代謝異常等骨骼畸形,伴隨較高的死亡率[4-6]。轉(zhuǎn)化生長(zhǎng)因子-β1(TGF-β1)調(diào)控相應(yīng)靶基因表達(dá)的前提是通過(guò)TGF-β1受體(TGF-β1 receptor,TβR)介導(dǎo)的,即處于活化狀態(tài)的TGF-β1分子與細(xì)胞膜上的TβR相結(jié)合。TβR主要包括Ⅰ、Ⅱ、Ⅲ型三種,其中Ⅰ型和Ⅱ型受體位于細(xì)胞膜上,直接參與信號(hào)的轉(zhuǎn)導(dǎo)[7-8]。研究發(fā)現(xiàn),在ClC-3氯通道調(diào)控成骨分化過(guò)程中,TGF-β1信號(hào)分子的表達(dá)與ClC-3氯通道有十分密切的聯(lián)系[9]。
ClC-3氯通道和TβR在促進(jìn)成骨分化中是否具有某種內(nèi)在的聯(lián)系,目前還不明確。本研究旨在初步探索TβR對(duì)ClC-3氯通道在成骨細(xì)胞中的促成骨分化作用,擬通過(guò)觀察成骨細(xì)胞中不同表達(dá)水平的ClC-3氯通道和TβR表達(dá)的相互影響,為ClC-3氯通道在促成骨分化機(jī)制方面研究提供新的思路。
1 材料與方法
1.1 材料
α改良伊格爾基本成分(α-MEM)培養(yǎng)液、胰蛋白酶、青霉素及鏈霉素(Hyclone公司,美國(guó));MC3T3-E1細(xì)胞系(此細(xì)胞株是小鼠成骨前體細(xì)胞,可以分化為成熟的成骨細(xì)胞。由西安飛揚(yáng)生物科技有限公司提供);TGF-β1受體抑制劑(LY2109761,Selleck公司,美國(guó));CCK-8試劑(Sigma公司,美國(guó));Trizol(Invitrogen公司,美國(guó));逆轉(zhuǎn)錄試劑盒(TaKaRa公司,日本);Real-Time PCR試劑盒(TaKaRa公司,日本);脂質(zhì)體2000(LipofectamineTM2000,Invitrogen公司,美國(guó));ClC-3抗體(CST,美國(guó));二抗(CST,美國(guó))。
1.2 TβR抑制劑(LY2109761)的配制
將5 mg TβR抑制劑(LY2109761)溶解于1.132 45 mL的二甲基亞砜(DMSO)溶液中,得到濃度為10 mmol/L的儲(chǔ)存液,以每管20 μL分裝于EP管中,-80 ℃保存?zhèn)溆谩9ぷ饕菏菍?chǔ)存液按濃度梯度稀釋到所需的20、2、0.2 μmol/L的濃度。
1.3 細(xì)胞培養(yǎng)與分組
將細(xì)胞系MC3T3-E1以1×104個(gè)/mL的密度接種于六孔板中,選取細(xì)胞生長(zhǎng)狀況良好,融合面積為80%左右的細(xì)胞給予LY2109761的刺激。分為三大組,A:空白對(duì)照組;B:0.2、2、20 μmol/L作用24 h組;C:0.2、2、20 μmol/L作用48 h組。
1.4 LY2109761細(xì)胞毒性檢測(cè)(CCK-8比色法)
以每孔約100 μL的細(xì)胞懸液(2×104個(gè)/mL)接種到96孔板中,在37 ℃、5%CO2的培養(yǎng)箱中培養(yǎng)24 h使細(xì)胞貼壁。向培養(yǎng)板中加入10 μL三種濃度的抑制劑(LY2109761)分別培養(yǎng)24和48 h。向每孔中分別加入10 μL的CCK-8溶液后在培養(yǎng)箱中繼續(xù)孵育4 h,測(cè)定各組細(xì)胞在450 nm處的吸光度值,以上實(shí)驗(yàn)步驟重復(fù)至少三次。其中細(xì)胞死亡率=100%-細(xì)胞活力,細(xì)胞活力=(A(加藥)-A(空白))/(A(0加藥)-A(空白))×100%。A(加藥)為具有細(xì)胞、CCK-8溶液和藥物溶液的孔的吸光度;A(空白)為具有培養(yǎng)基和CCK-8溶液而沒(méi)有細(xì)胞的孔的吸光度;A(0加藥)為具有細(xì)胞、CCK-8溶液而沒(méi)有藥物溶液的孔的吸光度。
1.5 RNA 提取和Real-time PCR
按照Trizol試劑盒說(shuō)明提取細(xì)胞總RNA,將LY2109761不同濃度不同作用時(shí)間處理后的各組細(xì)胞提取總RNA:選取生長(zhǎng)良好的細(xì)胞,棄去培養(yǎng)液,使用PBS輕輕沖洗兩遍;在每孔中加入Trizol,裂解,靜置。轉(zhuǎn)移上清液至新的EP管。在上清液中加入氯仿,至樣品分為透明上清、白色蛋白和有機(jī)物三層。吸出透明血清,加入異丙醇,棄去上清液。向沉淀物中加入75%的乙醇,棄去上清液。向沉淀中加入焦碳酸二乙酯(DEPC)水,使用分光光度計(jì)將提取物定量。
按照TaKaRa逆轉(zhuǎn)錄試劑盒說(shuō)明合成cDNA,然后將其稀釋10倍,將稀釋后的cDNA進(jìn)行Real-time PCR反應(yīng)。特異性引物序列如下(表1)。

表1 PCR引物序列及引物大小
1.6 Western Blot檢測(cè)
將細(xì)胞置于冰上,加入蛋白裂解液,離心,提取細(xì)胞總蛋白,蛋白質(zhì)定量試劑盒(BCA)法進(jìn)行蛋白定量。將蛋白放至100 ℃加熱5 min使其變性,凝膠電泳,轉(zhuǎn)膜,孵育一抗4 ℃過(guò)夜,孵育二抗,化學(xué)發(fā)光試劑盒(ECL)發(fā)光,使用Image-J軟件對(duì)蛋白進(jìn)行掃描及灰度值分析。
1.7 siRNA基因轉(zhuǎn)染
將MC3T3-E1細(xì)胞以2×104個(gè)/mL接種于六孔板中,待細(xì)胞密度融合至70%左右時(shí)進(jìn)行siRNA的轉(zhuǎn)染。分別使用Opti-MEM稀釋ClC-3 siRNA和Lipo2000轉(zhuǎn)染試劑,使轉(zhuǎn)染細(xì)胞的最終濃度為50 nmol/L,二者輕輕混合,室溫下靜置20 min。將轉(zhuǎn)染復(fù)合物加入到細(xì)胞中,孵育6 h,更換常規(guī)培養(yǎng)液繼續(xù)培養(yǎng)48 h。
1.8 統(tǒng)計(jì)學(xué)方法
以上實(shí)驗(yàn)結(jié)果應(yīng)用SPSS 17.0統(tǒng)計(jì)學(xué)軟件對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析,實(shí)驗(yàn)數(shù)據(jù)均以“均數(shù)±標(biāo)準(zhǔn)差”表示,采用單因素方差分析各實(shí)驗(yàn)組。以P<0.05為差異具有統(tǒng)計(jì)學(xué)意義。
2 結(jié) 果
2.1 不同作用方式的TβR抑制劑細(xì)胞毒性的結(jié)果
與空白對(duì)照組相比,0.2、2、20 μmol/L濃度的TβR抑制劑分別作用24 h和48 h對(duì)MC3T3-E1細(xì)胞都具有細(xì)胞毒性(P<0.05)。相同作用時(shí)間時(shí),抑制劑的細(xì)胞毒性隨作用濃度的增大而增大,不同作用濃度的抑制劑之間對(duì)細(xì)胞毒性作用均有統(tǒng)計(jì)學(xué)差異(P<0.05),其中0.2、2 μmol/L濃度的抑制劑作用24 h時(shí)的細(xì)胞毒性無(wú)統(tǒng)計(jì)學(xué)意義(P>0.05)。TβR抑制劑在相同作用濃度時(shí),作用時(shí)間越短,細(xì)胞毒性越小(P<0.05),以濃度為0.2 μmol/L的TβR抑制劑作用24 h對(duì)細(xì)胞的毒性作用最小(圖1)。
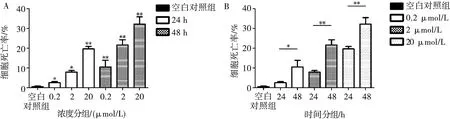
A:不同作用濃度的TβR抑制劑在相同作用時(shí)間對(duì)細(xì)胞毒性的影響;B:相同作用濃度的TβR抑制劑在不同作用時(shí)間對(duì)細(xì)胞毒性的影響;*:P<0.05,**:P<0.01
2.2 TβR抑制劑對(duì)TβRⅠ、TβRⅡ基因表達(dá)的影響
Real-Time PCR結(jié)果顯示,0.2、2 μmol/L濃度的TβR抑制劑作用MC3T3-E1細(xì)胞24 h后,都可以抑制TβRⅠ、TβRⅡ基因的表達(dá)(P<0.05)。而對(duì)TβRⅠ、TβRⅡ基因表達(dá)的抑制效果隨濃度增大而增強(qiáng)(P<0.05)(圖2)。
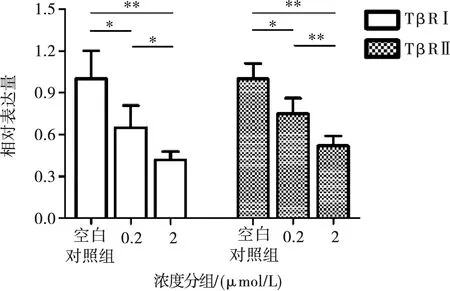
*:P<0.05,**:P<0.01
2.3 TβRⅠ、TβRⅡ?qū)lC-3氯通道表達(dá)的影響
濃度為2 μmol/L的TβR抑制劑對(duì)MC3T3-E1細(xì)胞作用24 h后的Real-Time PCR和Western結(jié)果表明,與空白對(duì)照組相比,TβR抑制劑組促進(jìn)ClC-3氯通道基因和蛋白的表達(dá)(圖3)。

A:阻斷TβRⅠ、TβRⅡ的表達(dá)對(duì)ClC-3基因的影響;B:阻斷TβRⅠ、TβRⅡ的表達(dá)對(duì)ClC-3蛋白的影響;C:ClC-3蛋白灰度分析;*:P<0.05
2.4 ClC-3氯通道對(duì)TβRⅠ、TβRⅡ表達(dá)的影響
對(duì)MC3T3-E1細(xì)胞進(jìn)行ClC-3 siRNA基因轉(zhuǎn)染,ClC-3 siRNA轉(zhuǎn)染序列如下,Sense:5′-CGA GAG AAG UGU AAG GAC ATT-3′;Anti-sense:5′-UGU CCU UAC ACU UCU CUC GTT-3′。轉(zhuǎn)染序列的有效性在前期實(shí)驗(yàn)已得到驗(yàn)證。Real-Time PCR結(jié)果顯示,阻斷ClC-3氯通道的表達(dá),可以促進(jìn)TβRⅠ和TβRⅡ基因的表達(dá)(P<0.05)(圖4)。

*:P<0.05,**:P<0.01
2.5 TβR對(duì)成骨分化相關(guān)基因(Alp、Runx2)表達(dá)的影響
分別使用ClC-3 siRNA和TβR抑制劑阻斷氯通道和TβR的表達(dá),觀察Alp、Runx2的變化。Real-Time PCR結(jié)果顯示,TβR抑制劑組的Alp和Runx2的表達(dá)均升高(P<0.05),而ClC-3抑制劑組的Alp和Runx2基因表達(dá)下降(P<0.05)(圖5)。

與空白對(duì)照相比,*:P<0.05,**:P<0.01
3 討 論
正畸牙齒的移動(dòng)是通過(guò)壓力側(cè)牙槽骨吸收,張力側(cè)牙槽骨新骨形成穩(wěn)定在新的位置。ClC-3氯通道作為電壓門控型氯通道之一,存在于細(xì)胞膜或細(xì)胞器膜上,在正畸骨改建過(guò)程中,ClC-3氯通道被證實(shí)參與骨代謝的生物信號(hào)傳導(dǎo)過(guò)程[10]。ClC-3氯通道在小鼠的骨髓間充質(zhì)干細(xì)胞、成骨細(xì)胞以及成骨前體細(xì)胞系中均可表達(dá), 能夠促進(jìn)成骨細(xì)胞的成骨分化[11]。TGF-β1生長(zhǎng)因子在骨架形態(tài)發(fā)生和成骨細(xì)胞分化過(guò)程中具有重要作用[12-14]。研究表明,TGF-β1與ClC-3氯通道的表達(dá)有緊密聯(lián)系[9]。阻斷TGF-β1表達(dá)可促進(jìn)ClC-3氯通道對(duì)骨代謝的調(diào)控。TGF-β1通常是以異構(gòu)受體復(fù)合體發(fā)揮作用,與TβRⅠ、TβRⅡ在細(xì)胞表面結(jié)合后引發(fā)后續(xù)的細(xì)胞應(yīng)答[15]。但是關(guān)于TβR在ClC-3氯通道調(diào)控成骨分化過(guò)程中的作用目前還未見(jiàn)相關(guān)研究。
TβR抑制劑按作用方式分為選擇性單獨(dú)抑制TβRⅠ或TβRⅡ以及同時(shí)抑制TβRⅠ和TβRⅡ表達(dá)等幾種類型[16]。本研究通過(guò)選用同時(shí)抑制TβRⅠ和TβRⅡ表達(dá)的小分子受體激酶抑制劑(LY2109761)來(lái)阻斷TGF-β1下游受體,觀察對(duì)ClC-3氯通道在成骨分化過(guò)程中的作用。首先,本實(shí)驗(yàn)將TβR抑制劑對(duì)TβRⅠ和TβRⅡ基因的最佳作用方式進(jìn)行篩選,根據(jù)細(xì)胞的不同,許多文獻(xiàn)研究使用了0.2~10.0 μmol/L的濃度范圍的同種抑制劑作用不同時(shí)間來(lái)研究細(xì)胞功能[17-18],但是還沒(méi)有對(duì)TβR抑制劑作用于MC3T3-E1細(xì)胞進(jìn)行過(guò)多的研究。經(jīng)過(guò)預(yù)試驗(yàn)的篩選,最終選取了0.2、2、20 μmol/L三種濃度梯度分別作用24 h和48 h來(lái)觀察TβRⅠ和TβRⅡ抑制劑對(duì)MC3T3-E1的細(xì)胞毒性。細(xì)胞毒性試驗(yàn)結(jié)果表明隨著抑制劑濃度增大、作用時(shí)間延長(zhǎng)其細(xì)胞死亡率也在升高,這說(shuō)明抑制劑對(duì)細(xì)胞的毒副作用隨著劑量和作用時(shí)間而增大,并且抑制劑在0.2 μmol/L和2 μmol/L的濃度作用24 h時(shí)細(xì)胞的死亡率并無(wú)統(tǒng)計(jì)學(xué)意義,這可能說(shuō)明TβR抑制劑在對(duì)MC3T3-E1細(xì)胞作用時(shí)間較低時(shí),作用濃度在較小的范圍之間發(fā)生波動(dòng)對(duì)細(xì)胞的毒性沒(méi)有太大影響,而超出一定濃度,則會(huì)提高細(xì)胞的死亡率。而將0.2 μmol/L和2 μmol/L兩種濃度梯度的抑制劑作用24 h來(lái)觀察TβRⅠ和TβRⅡ兩種受體基因的表達(dá)結(jié)果顯示,2 μmol/L的TβR抑制劑對(duì)TβR I、TβRⅡ基因表達(dá)的抑制效果強(qiáng)于0.2 μmol/L濃度。濃度為2 μmol/L的TβR抑制劑作用于MC3T3-E1細(xì)胞24 h對(duì)TβRⅠ和TβRⅡ基因的抑制效果已經(jīng)達(dá)到65%左右,并且產(chǎn)生的細(xì)胞毒性在可接受的范圍內(nèi),不會(huì)對(duì)細(xì)胞的生長(zhǎng)和增殖產(chǎn)生過(guò)度的抑制。
綜上,本研究選取濃度為2 μmol/L的TβR抑制劑作用MC3T3-E1細(xì)胞24 h作為最佳作用方式抑制TβR的表達(dá),并進(jìn)一步觀察其對(duì)ClC-3氯通道的影響。結(jié)果顯示,當(dāng)抑制TβR表達(dá)時(shí),ClC-3氯通道的基因和蛋白水平表達(dá)升高。另一方面,當(dāng)阻斷ClC-3氯通道表達(dá)時(shí),同樣也會(huì)促進(jìn)TβR的表達(dá)。這說(shuō)明TβR和ClC-3氯通道的表達(dá)是相互抑制的。由此猜想,這可能和TβRⅠ、TβRⅡ以及ClC-3氯通道的細(xì)胞內(nèi)定位有關(guān)。TβRⅠ、TβRⅡ位于細(xì)胞膜上,ClC-3氯通道細(xì)胞膜和細(xì)胞器膜上也有表達(dá),基于此推測(cè),TβR和ClC-3在細(xì)胞膜上的表達(dá)存在競(jìng)爭(zhēng)性關(guān)系,當(dāng)其中一個(gè)的表達(dá)被抑制時(shí),可促進(jìn)對(duì)方的表達(dá),因此,ClC-3氯通道與TβR的表達(dá)可能表現(xiàn)為相互影響的關(guān)系。我們?cè)诟蓴_ClC-3氯通道表達(dá)時(shí),TβRⅡ的表達(dá)與空白對(duì)照組相比升高了80%,而TβRⅠ表達(dá)升高了50%,這說(shuō)明,TβRⅡ?qū)Σ煌磉_(dá)水平的ClC-3氯通道更敏感,這可能和TβRⅡ本身的生物學(xué)特性有關(guān),TβRⅡ通常首先通過(guò)自身的激活,才能使TGF-β1配體和胞膜上活化的Ⅱ型受體結(jié)合,形成異源二聚體,然后再結(jié)合Ⅰ型受體形成異三聚體復(fù)合物。本研究發(fā)現(xiàn)抑制TβR的表達(dá),可促進(jìn)成骨相關(guān)基因(Alp、Runx2)的表達(dá),而ClC-3抑制劑組的成骨相關(guān)基因表達(dá)與對(duì)照組相比卻下降,其中TβR抑制劑組的Alp基因升高程度較Runx2基因明顯。Alp是成骨分化過(guò)程中的早期標(biāo)志物,在骨形成早期活性較高[19],Runx2是成骨細(xì)胞分化過(guò)程中關(guān)鍵的轉(zhuǎn)錄調(diào)控因子,發(fā)揮中心調(diào)控的作用[20-21]。由此推測(cè)TβR在早期對(duì)ClC-3氯通道調(diào)控成骨分化的影響更顯著。綜上所述,在正畸骨改建過(guò)程中,TβR可抑制ClC-3氯通道對(duì)成骨細(xì)胞分化的調(diào)控,但是其具體的分子機(jī)制還有待進(jìn)一步深入的研究。
