羽衣甘藍BoWRKY10基因克隆及其轉基因煙草耐旱性研究
2022-02-08 12:09:06趙紅啟李鴻雁
西南農業學報 2022年11期
趙紅啟,楊 勇,閆 艷,范 杰,李鴻雁
(1.黃淮學院園林中心,河南 駐馬店 463000;2. 黃淮學院生物與食品工程學院,河南 駐馬店 463000)
【研究意義】植物生長在不斷變化的自然環境條件下,受到各種生物和非生物脅迫。為了提高生存率,植物必須通過進化和調節生長機制來適應這些惡劣的環境。作為主要的非生物脅迫,干旱是對植物的生長和發育產生不利影響,甚至對農業產量造成嚴重損失的重要環境脅迫因子之一。因此,研究與植物抗逆性相關的基因及其分子機制,對于植物抗逆力的遺傳改良和人類社會的可持續發展具有重要意義。WRKY轉錄因子在調控機制中扮演著十分重要的角色,近幾年備受關注。克隆WRKY相關基因,研究其調控機制,對于闡明植物轉錄因子的調控機制具有重要意義。【前人研究進展】轉錄因子(Transcription factors,TFs)是脅迫信號轉導和靶基因表達的重要介質。TFs通過調節下游靶基因的表達水平,在保護機體免受應激相關損傷中發揮重要作用[1-2]。大量研究表明,TFs基因的過表達激活了一組靶基因,這些靶基因協同抵御非生物脅迫的不利影響。因此,TFs基因工程是提高作物抗逆性的有效途徑。擬南芥(Arabidopsisthaliana)基因組中有1500多個TFs,約占所有基因的6%[3]。在這些TFs中,WRKY作為植物中最大的轉錄因子家族之一,在植物對干旱脅迫響應的調控網絡中發揮重要作用4]。WRKY蛋白是一種新型的植物鋅指轉錄調控因子,其N端含有7個由WRKYGQK組成的氨基酸序列。WRKY蛋白是由1個或2個WRKY保守結構域。WRKY-TF基因受生物和非生物脅迫誘導,參與植物脅迫響應[5]。WRKY-TFs是植物基因組中的一個大基因家族。擬南芥中有74個基因[4],水稻(OryzasativaL.)中有100多個基因[6],小麥(TriticumaestivumL.)中有119個基因[7],小白菜(BrassicacampestrisL.)中有46個基因[4]。WRKYs被認為是參與生物脅迫響應的關鍵TFs,這些實驗數據為WRKY調控非生物脅迫響應提供了證據。此外,一些TFs對干旱脅迫特別敏感。例如,金錢草(DichondrarepensL)WRKY70基因通過調控精氨酸脫羧酶基因在抗旱和腐胺合成中發揮作用。轉二穗短柄草(BrachypodiumdistachyonL.)的BdWRKY36基因對干旱脅迫的耐受性顯著增強[8]。另外2個WRKY基因(ABO3和ThWRKY4)已被證明是干旱脅迫響應中的關鍵調控基因[9]。MuWRKY3基因在花生(ArachishypogaeaL.)中的過表達是通過減少MDA、過氧化氫和超氧陰離子的積累以及增加脯氨酸含量、可溶性糖含量和抗氧化的活性顯著提高花生的耐旱性[10]。雖然一些TFs在模式植物中具有生理功能,但大多數TFs的功能尚不清楚,特別是在羽衣甘藍(Brassicaoleraceavar.acephalaDC)等非模式植物中。【本研究切入點】WRKY家族基因在植物抗旱育種中具有廣泛的應用前景,前期在擬南芥、小白菜和油菜(BrassicacampestrisL.)等十字花科中發現一些WRKY家族轉錄因子基因在植物逆境中起重要作用,但在羽衣甘藍中鮮有研究。【擬解決的關鍵問題】本研究以羽衣甘藍為材料,利用轉基因技術探究BoWRKY10基因在羽衣甘藍耐旱性中的生物學功能。探明在羽衣甘藍中超表達BoWRKY10對羽衣甘藍抗旱性的影響,明確該基因在羽衣甘藍響應干旱脅迫的生物學功能其在旱育種中的價值。
1 材料與方法
1.1 試驗材料
羽衣甘藍種質材料“葉牡丹”由黃淮學院園林中心提供。煙草NC89及pCAMBIA1301載體由黃淮學院生物與食品工程學院實驗室保存。植物在溫室中用混合營養土[m(蛭石):m(營養土)=1∶1]培養,培養溫度為24 ℃/22 ℃(白天/黑夜),濕度為65%,光照周期為光照(L):黑暗(D)=16 h:8 h。
1.2 RNA提取與載體構建
用植物總RNA分離試劑盒(TaKaRa,大連,中國)從羽衣甘藍的葉片中提取總RNA。根據試劑盒的說明,使用逆轉錄酶試劑盒(TaKaRa,大連,中國)合成第一鏈cDNA。從NCBI數據庫中獲得羽衣甘藍WRKY10基因序列,利用Primer Premier 5.0分別設計了特異的正向和反向引物(表1)。擴增的PCR產物純化后,被克隆到pMD18-T載體( TaKaRa,大連,中國)上并測序。
1.3 煙草轉化與篩選
把測序后正確的序列與含有新霉素磷酸轉移酶基因(Neomycin phosphotransferase gene, NPTII)和35S啟動子(CAMV35S)的pCAMBIA1301雙元載體相連。將pCAMBIA-BoWRKY10導入根癌農桿菌LBA4404中。根據Horsch等[11]的方法轉化煙草。將T0轉基因植株的種子收獲并接種在含有卡那霉素(100 mg/L)的MS培養基上。用qPCR引物擴增BoWRKY10基因(表1)。采用定量PCR(qPCR)技術檢測BoWRKY10在轉基因植株中的表達。
1.4 試驗設計
取適量野生型(WT)和轉基因煙草種子于無菌2 mL離心管中,70%酒精消毒30 s,無菌水洗1次,10%(v/v)H2O225 ℃消毒 10 min,無菌水洗5~6次,點播于含有15% PEG4000的 1/2 MS培養基上,黑暗1 d后,28 ℃光照培養,每隔24 h 統計發芽率;正常萌發的種子移栽到 5%、10%和15% PEG4000的1/2 MS條件14 d后,統計根長,每個株系取12株;煙草于溫室培養30 d后,15% PEG4000處理,24 h后取材料檢測 BoWRKY10和相關脅迫響應因子的表達量,1周后測定SOD、POD、CAT活性和H2O2含量,每個株系取8株。在抗旱性試驗中,每個株系取生長相似的5周齡植物30株,停止澆水14 d后復水,計算存活率。在干旱第7天時,取樣測定H2O2含量以及超氧化物歧化酶(SOD)、過氧化物酶(POD)和過氧化氫酶(CAT)活性和相對含水量(RWC)、丙二醛(MDA)、脯氨酸、可溶性糖含量。所有測定按李玲的方法[12]進行。每個實驗3個重復。

表1 引物序列及用途
1.5 實時定量PCR在煙草中的表達分析
用TRIzol試劑提取總RNA,取2 μg RNA合成cDNA。以表1所列的序列為引物,cDNA 為模板,用ABI7300實時熒光定量PCR系統(ABI,Foster City,CA,USA)進行qPCR,3次重復。反應程序為95 ℃10 s,94 ℃10 s,60 ℃25 s,40個循環。反應結束后分析熒光值變化曲線和融解曲線。每個反應3次重復,采用2-ΔΔCt算法分析結果。
1.6 統計分析
利用Excel 2016和SPSS 17.0中進行數據分析,并從3個獨立實驗中計算平均值±標準差。兩組間比較采用t檢驗,P<0.05被認為是差異顯著。
2 結果與分析
2.1 BoWRKY10基因轉化煙草及陽性植株篩選
利用Primer 5.0軟件設計帶酶切位點的特異性引物擴增目的片段,構建pBIA-35S-BoWRKY10植物表達載體。利用農桿菌介導法將該重組質粒轉化到煙草中,100 mg/mL Kan培養基上篩選出長勢較好的煙草進行PCR檢測。結果表明,在轉基因煙草中檢測到9個陽性株系。野生型煙草葉片中沒有檢測到目的條帶。qRT-PCR分析表明,9個株系BoWRKY10基因均能過表達,但OE-3、OE-4和OE-6表達量較高(圖1)。因此,以下研究選取這3個株系作為研究材料。
2.2 轉基因煙草對干旱的耐受性分析
為研究外源BoWRKY10基因表達對植物干旱脅迫響應的影響,選擇相對表達量較高的OE-3、OE-4 和OE-6轉基因煙草T2代株系進行耐旱性分析,結果(圖2-A)顯示,轉BoWRKY10基因的煙草比WT對干旱處理產生了一定的耐性。對5周齡的轉基因植株和野生植株進行干旱處理, 干旱處理14 d后,3個轉基因株系葉只有少許萎焉,而野生型煙草完全萎焉,復水后轉基因株系恢復較好,轉基因株系OE-3、OE-4 和OE-6存活率分別為60.6%、63.3%和61.0%,而野生型對照存活率僅為22.2%,轉基因株系存活率顯著高于野生型對照(圖2-B) 。
2.3 干旱脅迫下轉基因煙草種子萌發率及根長
種子在含15% PEG4000 1/2 MS培養基中萌發14 d后,3個轉基因株系主根長顯著大于野生型 (圖3-A,3-B)。在正常1/2 MS培養基上,轉基因株系OE-3、OE-4 和OE-6的發芽率和野生型對照沒有明顯差異(圖3-C);而15%PEG4000培養基中,OE-3、OE-4 和OE-6萌發率分別為65.9%、75.4%和84.3%,野生型僅為22.5%(圖3-D);與野生型對照相比,轉基因株系萌發率分別提高了43.4%、52.9%和 61.8%(圖3-D)。
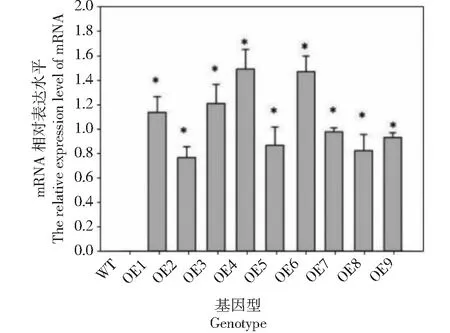
*表示與對照有顯著差異(P<0.05),下同* denotes significant differences as compared with the control group(P<0.05), the same as below圖1 轉基因煙草中BoWRKY10基因過表達分析Fig.1 Overexpression analysis of BoWRKY10 gene in transgenic tobacco
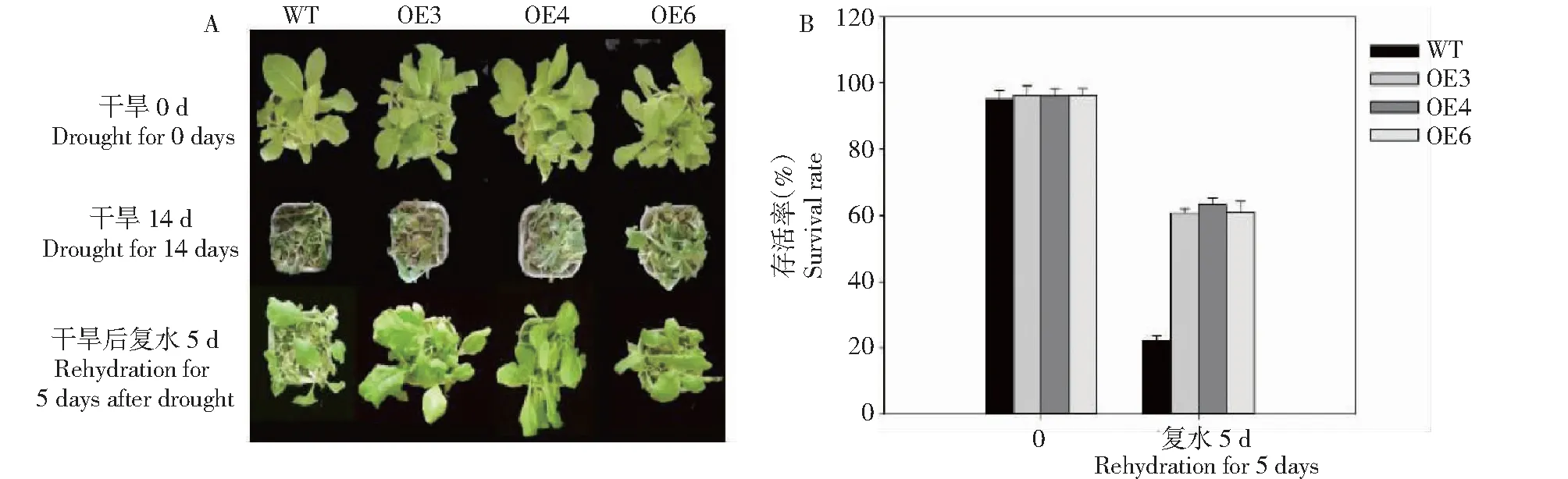
圖2 干旱脅迫下轉基因煙草的表型和存活率Fig.2 Phenotype and survival rate of transgenic tobacco under drought stress
2.4 干旱脅迫下BoWRKY10過表達對煙草生理特性的影響
轉基因植株與野生型(WT)植株在處理前脯氨酸和MDA含量差異不顯著。干旱脅迫后,轉BoWRKY10基因植株和WT植株的脯氨酸和MDA含量增加,且轉BoWRKY10基因植株的脯氨酸含量顯著高于WT植株(圖4)。相比之下,轉基因BoWRKY10株系的MDA含量顯著降低。在干旱脅迫后,BoWRKY10轉基因植株和WT植株的可溶性糖含量增加,但BoWRKY10轉基因植株的可溶性糖含量顯著高于WT植株。干旱處理7 d后,BoWRKY10過表達株系的RWC顯著高于WT。
干旱脅迫下轉基因煙草葉片中SOD、POD、CAT活性顯著高于對照,而轉基因植株中的H2O2水平較低(圖5)。此外,在MS培養基上培養2周的幼苗,移栽到含PEG4000(mol/L,15%)的MS培養基上1周,檢測結果與干旱脅迫結果相似。表明,在干旱脅迫下,BoWRKY10的表達通過增加3種重要抗氧化酶的活性影響ROS水平。
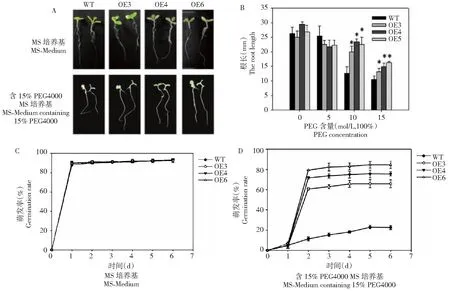
圖3 不同脅迫條件下過表達BoWRKY10基因煙草植株的根長和萌發率Fig.3 Root length and germination rate of tobacco plants overexpressing the BoWRKY10 gene under different stress conditions
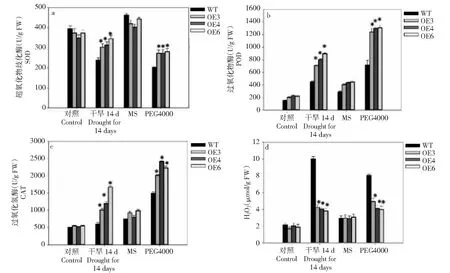
**表示與對照有極顯著差異(P<0.01),下同** denotes significant differences as compared with the control group (P<0.01), the same as below圖5 正常和干旱滲透脅迫條件下野生型(WT)和轉基因煙草中抗氧化酶活性和H2O2積累Fig.5 Three antioxidant enzyme activities and H2O2 accumulation in WT and transgenic tobacco under normal and drought/osmotic conditions
2.5 BoWRKY10調節干旱脅迫下應答基因的表達
以2周齡對照和轉基因煙草(OE-3)植株為材料,PEG4000(m/mol/L,15%)處理1周,NtSOD、NtAPX、NtCAT、NtGST、NtSPSAt和NtNCED1等6個ROS相關的脅迫響應基因進行qPCR檢測。與對照植物相比,當用PEG4000時,轉基因植物中所檢測的應答基因均顯著上調(圖6),表明BoWRKY10在煙草中的表達通過誘導一些ROS相關基因和脅迫響應基因的表達,增強了煙草的抗旱性。
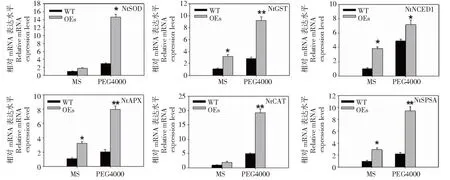
圖6 干旱處理下野生型(WT)和 BoWRKY10轉基因煙草脅迫響應相關基因表達Fig.6 Expression of stress response genes in WT and BoWRKY10 transgenic tobacco under drought conditions
3 討 論
WRKY-TFs是植物中最大的轉錄調控家族之一,具有多種功能環境脅迫下的發育和生理功能。近年來,在水稻[6]、大豆[13]、棉花[14]、短梗草[8]、玉米[15]和小麥[7]等植物中發現了大量的WRKY蛋白。隨著研究的深入,發現植物WRKY 轉錄因子對不同的脅迫環境均有應答。已有研究發現,過表達TaWRKY46增強了轉基因擬南芥植株的滲透脅迫耐受性,其轉基因擬南芥植株在含甘露醇的1/2 MS培養基中具有較高的發芽率和較長的主根[7]。番茄SlWRKY39是擬南芥AtWRKY40的同源基因,它能被鹽、干旱及ABA等相關脅迫因子誘導表達[16]。但Liu等[17]對棉花研究表明,轉基因植株中GhWRKY25的過表達降低了抗旱性。本研究表明,當把羽衣甘藍的BoWRKY10基因克隆到煙草中,其過表達增強了轉基因煙草的抗旱性;在干旱脅迫下,轉基因株系的根長、發芽率及存活率均顯著高于野生型。
在干旱脅迫下,ROS的過量產生會導致氧化損傷,通過激活活性氧清除酶進行解毒[18-19]。MDA濃度是膜脂過氧化的一個參數,反映了響應程度。因此,丙二醛含量通常被用作非生物脅迫下植物受損的參數[20]。抗氧化酶系統,如POD、SOD和CAT酶,通過將ROS保持在閾值水平來保護受脅迫的植物,這些酶的活性表明植物在非生物脅迫下的抗脅迫能力[21-22]。本研究中,與野生型煙草相比,轉基因煙草在干旱脅迫下膜損傷程度較低,表現為電解質滲漏、ROS和MDA積累減少,表明活性氧清除系統的增強能力與BoWRKY10過表達植株的抗旱性增加密切相關,即BoWRKY10通過激活ROS清除系統提升煙草抗旱能力。
在干旱脅迫下,一些脅迫相關基因表達水平上調或下調。例如,GmWRKY16的過表達使脅迫相關標記基因RD29A、RD22、KIN1和LEA14的表達顯著上調[23]。WRKY17s在其他植物中的負調控也有報道。在鹽脅迫下,轉基因擬南芥CmWRKY17中AtRD29A和ATDREB2的表達水平顯著下調[24]。此外,HaWRKY76的過表達通過調節RAB18、RD29A和RD29B的表達增強了轉基因擬南芥的耐旱性[25]。本研究表明,在干旱脅迫下,NtSOD、NtAPX、NtCAT、NtGST、NtSPSAt和NtNCED1等6個基因表達上調。推測BoWRKY10可能通過直接或間接調控干旱脅迫相關基因來調控干旱脅迫。在今后研究中,對BoWRKY10蛋白與DNA結合位點及其BoWRKY10蛋白在其他環境脅迫中功能等方面進行研究,探索其在基因調控中作用機理及其生理功能。
4 結 論
過表達羽衣甘藍BoWRKY10基因煙草顯著提高煙草在干旱脅迫下成活率、萌發率及抗氧化酶活性,增加可溶性糖含量和減少MDA含量;NtSOD、NtAPX、NtCAT、NtGST、NtSPSAt和NtNCED1等6個ROS相關的脅迫響應基因表達上調。本研究表明BoWRKY10基因與干旱脅迫相關。這對培育羽衣甘藍新品種、探索基因調控機理等具有理論與實踐意義。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
聲屏世界(2014年6期)2014-02-28 15:18:09
