Clostridium butyricum蔗糖磷酸化酶的酶學性質及其功能研究*
2022-02-09 12:51:48譚姚姚何賀賀王宏萌黃日波杜麗琴
廣西科學 2022年6期
譚姚姚,何賀賀,王宏萌,梁 蒙,林 宇,黃日波,杜麗琴**
(1.廣西大學生命科學與技術學院,廣西微生物與酶工程技術研究中心,廣西南寧 530005;2亞熱帶農業生物資源保護與利用國家重點實驗室,廣西南寧 530005)
糖苷類化合物通常是指由單糖或多糖的半縮醛羥基與配體兩部分縮合而成的含糖衍生物,配體可以是糖類,也可以是非糖類物質——蛋白質、脂肪醇等[1]。糖苷類化合物不僅可為生物體提供能量、調節植物的生長,而且在生產和生活中使用也非常廣泛[2]。如在醫藥行業當中,環烯醚萜苷類化合物、芳香碳糖苷化合物、三萜苷類化合物等糖苷類化合物因具有極大的藥理性價值,被用于治療營養不良、中風等多種癥狀[3,4]。在化妝品行業,α-熊果苷是一種十分有效的美白成分,而甘油糖基化后生成的2-O-(α-D-glucopyranosyl) -sn-glycero是一種很好的保濕劑(Glycoin?)[5,6]。此外,烷基糖苷作為一類可降解、毒性低的優質非離子型表面活性劑被大家所熟知[7]。因此,具有極大應用價值的糖苷類化合物引起了人們的研究熱潮。除了直接從自然界獲取外,人們還可以使用多種方法來合成糖苷類化合物,如化學法、生物催化法等,通過對糖類或非糖類化合物糖基化修飾來形成新的糖苷鍵,從而合成糖苷類化合物。
蔗糖磷酸化酶主要存在于微生物體內,屬于糖基水解酶GH13家族。除了能夠水解蔗糖外,蔗糖磷酸化酶還具有催化糖苷鍵轉移的作用[8,9],因此它可以作為一種生物催化劑,將底物合成糖苷類化合物[10]。Wang等[11]利用蔗糖磷酸化酶合成曲二糖,使得益生元曲二糖得到了大規模的生產;對減輕過敏性皮炎癥狀有益的低聚糖——蜜二糖也可以通過蔗糖磷酸化酶來催化合成[5];經過蔗糖磷酸化酶的作用,可以將苯二酚分子催化加上葡萄糖基,形成具有美白作用的熊果苷[12]。Gudiminchi等[13]利用蔗糖磷酸化酶對維生素C進行葡萄糖糖基化,使其不穩定性得到很大程度的改善。此外,蔗糖磷酸化酶還可對兒茶素[14]、白藜蘆醇[15]、甜菊醇[16]、槲皮素[17]等進行轉糖苷催化。因此,蔗糖磷酸化酶在轉糖苷催化方面有較大的應用價值。目前,未有關于丁酸梭菌Clostridiumbutyricum中蔗糖磷酸化酶的相關報道,本研究對其酶學性質和轉糖苷功能進行探討,并進一步討論不同來源的蔗糖磷酸化酶的差異性,以挖掘出具有利用價值的蔗糖磷酸化酶,提高其在工業應用中的潛力。
1 材料與方法
1.1 材料
1.1.1 菌株與質粒
丁酸梭菌C.butyricumDSM10702、表達載體pQE30、感受態細胞Escherichia.coliXL1-blue等均由本實驗室保存。
1.1.2 主要試劑和儀器
試劑:PrimerSTARTMHS DNA聚合酶、T4DNA 連接酶、dNTP、λ/Hind Ⅲ DNA marker、限制性內切酶BamH Ⅰ、PstⅠ等均購買自Takara(寶)生物股份有限公司;膠回收/PCR純化試劑盒、質粒DNA提取試劑盒以及純化試劑盒均購自Fluxion Biosciences公司;蔗糖購自生工生物工程(上海)股份有限公司;葡萄糖-1-磷酸(G-1-P)、兒茶素等購買自上海源葉生物科技有限公司。
儀器:PCR儀(T-Gradient Thermoblock、MJ Reserch PTC-2000,蘇州亞凡生物技術有限公司),離心機(Eppendorf Centrifuge 5415D,上海智巖科學儀器有限公司),高效液相色譜儀(Agilent G1314F,杭州瑞析科技有限公司),隔水式恒溫培養箱(Binder,上海齊欣科學儀器有限公司),恒溫振蕩搖床(ZHWY-211B,上海知楚儀器有限公司)。
1.2 方法
1.2.1 蔗糖磷酸化酶基因的克隆
通過對高通量全基因組從頭測序丁酸梭菌的結果分析,查找糖基水解酶13家族中被注釋為蔗糖磷酸化酶的核苷酸序列,并對丁酸梭菌的蔗糖磷酸化酶基因所編碼的蛋白質進行結構組件分析,充分了解其結構,然后利用軟件Vector NTI Advance 11.5進行引物設計,所設計的引物序列如下(其中下劃線表示酶切位點):
cbsp-F:5′-CTCGGATCCGATAAGATAAAA-
AATGAAATAATG-3′;
cbsp-R:5′-CTACTGCAGCTCAATAGTAAA-
ATCATAACTCTT-3′。
PCR程序:98℃ 3 min;98℃ 15 s,56.5℃ 10 s,72℃ 110 s,30個循環;72℃ 10 min。將PCR擴增所得的DNA片段及提取的質粒pQE30分別經BamH Ⅰ和Hind Ⅲ雙酶切處理后,在T4DNA 連接酶下進行連接,然后轉化到E.coliXL1-blue中。轉化子送生工生物工程(上海)股份有限公司進行測序,將構建成功的重組質粒命名為pQE30-cbsp。
1.2.2 重組酶Cbsp的生物信息學分析
在http://smart.embl-heidelberg.de/網站對蔗糖磷酸化酶基因cbsp編碼的氨基酸序列進行SMART Main page分析,在http://www.cbs.dtu.dk/services/SignalP/網站對重組酶Cbsp進行SignalP 4.1信號肽分析。
1.2.3 重組酶Cbsp的誘導表達及純化
將測序正確的重組質粒轉入E.coliXL1-blue感受態細胞中,于37℃、220 r·min-1培養至OD600為0.4-0.6時,加入終濃度為0.5 mmol·L-1的 IPTG,在20℃、180 r·min-1條件下誘導22 h,離心收集菌體。超聲儀破胞后經鎳親和層析(Ni-NTA)純化重組酶Cbsp,并進行變性聚丙烯酰胺凝膠電泳(SDS-PAGE)分析。
1.2.4 重組酶Cbsp的酶學性質分析
酶活力測定方法[18]:取170 μL的磷酸-檸檬酸緩沖液、20 μL 10%的蔗糖溶液,放置在最適溫度水浴鍋中水浴2-3 min后,加入10 μL適當稀釋的酶液,精確反應20 min,然后加入400 μL DNS終止反應,沸水浴5 min顯色。冷卻至室溫后,取200 μL混合液置于96孔酶標板中,在540 nm波長處讀取樣品的吸光值。對照為用10 μL滅活后的酶液替代原反應體系的酶液。酶活單位(U/mL)=(葡萄糖含量×103×稀釋倍數)/(198.17×20×0.01),其中:198.17為葡萄糖分子量,20為反應時間(min),0.01為粗酶液體積(mL)。
(1) pH值對重組酶的影響。
在常溫條件下,測定不同pH值(3.0,3.5,4.0,4.5,5.0,5.5,6.0,6.5,7.0,7.5,8.0)的磷酸氫二鈉-檸檬酸緩沖液對重組酶酶活力的影響,得到重組酶的最適pH值。將重組酶保存在不同pH值(3.0,3.5,4.0,4.5,5.0,5.5,6.0,6.5,7.0,7.5,8.0的磷酸氫二鈉-檸檬酸;8.0,8.5,9.0的硼酸-硼砂;9.0,9.5,10.0,10.5的甘氨酸-氫氧化鈉)的緩沖液中,于4℃冰箱放置12 h;然后,根據酶活力測定方法,將經不同pH值緩沖液處理過的酶液加入反應體系中,測定重組酶的pH穩定性。每組3個平行,計算不同pH值下重組酶的酶活力。相對酶活力是指以最高酶活力為100%,計算出其他條件下酶活力的相對比例。
(2)溫度對重組酶的影響。
在最適pH值條件下,測定不同反應溫度(25-65℃,間隔5℃)對重組酶酶活力的影響,得到重組酶的最適溫度。將重組酶放置在不同溫度(4℃、35℃、40℃、45℃和50℃)下,測定隨時間變化重組酶的酶活力,即重組酶的熱穩定性。每組3個平行,計算不同溫度條件下重組酶的酶活力。
(3)Km和Vmax的測定。
在最適pH值和最適溫度條件下,測定以不同濃度(1.5-234.0 mmol·L-1)蔗糖為底物時重組酶的酶活力,使用軟件GraphPad Prism 5通過非線性回歸分析擬合出重組酶的Km、Vmax。
(4)化學試劑對重組酶的影響。
在最適反應條件下,分別測定N,N-二甲基甲酰胺(DMF)、二甲基亞砜(DMSO)、乙腈、甲醇、乙醇、異丙醇、正丁醇、乙二醇、1,2-丙二醇、1,3-丙二醇、2,3-丁二醇等醇類(各試劑濃度設為5%、10%、15%、20%、25%、30%,V∶V)對重組酶酶活力的影響,同時還測定0.05%(W∶V,下同)SDS、1%(V∶V,下同)Triton X-100、5%(V∶V,下同)Tween-80、0.01 mol·L-1咪唑、4 mol·L-1脲、1%(V∶V,下同)巰基乙醇等蛋白質還原劑和變性劑對重組酶酶活力的影響。以不添加化學試劑的反應作為對照,每組3個平行,計算不同濃度化學試劑存在時重組酶的酶活力。
1.2.5 轉糖苷功能的測定
(1)以糖類和糖醇類為受體。
在最適反應條件下,分別以終濃度15% (V∶V,下同)的G-1-P和8% (W∶W,下同)的蔗糖為糖基供體,以終濃度10% (V∶V)的D-/L-阿拉伯糖、L-山梨糖、D-半乳糖、L-鼠李糖、D-甘露糖、D-果糖、D-木糖、D-甘露糖、纖維二糖、海藻糖、麥芽糖、甘露醇和木糖醇為糖基受體,加入適量的純酶反應12 h后煮沸10 min終止反應,通過HPLC檢測產物。
檢測條件:儀器設備為Agilent 1260系列液相色譜儀、示差折光檢測器,色譜柱為Alltima Aminoz氨基柱(4.6 μm×250 mm,5 μm),流動相為70% (V∶V)乙腈,等度洗脫,進樣量10 μL,柱溫25°C,流速1 mL·min-1。
(2)以醇類為受體。
①在最適反應條件下,分別以終濃度15%的G-1-P和8%的蔗糖為糖基供體,以終濃度5%(V∶V)的甲醇、乙醇、異丙醇、正丙醇、1,2-丙二醇、1,3-丙二醇、2,3-丁二醇、乙二醇和丙三醇為糖基受體,加入適量的純酶反應12 h后煮沸10 min終止反應,通過HPLC檢測產物。
檢測條件:儀器設備為Agilent 1260系列液相色譜儀、示差折光檢測器,色譜柱為Alltima Aminoz氨基柱(4.6 μm×250 mm,5 μm),流動相為83% (V∶V)乙腈,等度洗脫,進樣量10 μL,柱溫25℃,流速1 mL·min-1。
②在最適反應條件下,分別以終濃度15% 的G-1-P和8%的蔗糖為糖基供體,以200 μL正己醇、正辛醇、正壬醇和正癸醇為糖基受體,加入適量的純酶反應12 h后煮沸10 min終止反應,通過HPLC檢測產物。
檢測條件:儀器設備為蒸發光散射檢測器,色譜柱為C18柱,流動相為75% (V∶V)甲醇,等度洗脫,進樣量10 μL,流速0.8 mL·min-1。
(3)以烷基糖苷類物質為受體。
在最適反應條件下,分別以終濃度15%的G-1-P和8%的蔗糖為糖基供體,以終濃度1%的己基-β-D-葡萄糖苷、庚基-β-D-葡萄糖苷、N-辛基-β-D-葡萄糖苷、壬基-β-D-葡萄糖苷和0.5%的癸基-β-D-葡萄糖苷為糖基受體,加入適量的純酶反應12 h后煮沸10 min終止反應,通過HPLC檢測產物。
檢測條件:儀器設備為蒸發光散射檢測器,色譜柱為C18柱,流動相為75% (V∶V)甲醇,等度洗脫,進樣量10 μL,流速1 mL·min-1。
(4)以L-抗壞血酸和兒茶素類物質為受體。
在最適反應條件下,分別以終濃度15%的G-1-P和8%的蔗糖為糖基供體,以終濃度120 mmol·L-1L-抗壞血酸和終濃度35 mmol·L-1兒茶素為糖基受體,加入適量的純酶反應12 h后煮沸10 min終止反應,通過HPLC檢測產物。
L-抗壞血酸檢測條件:儀器設備為Agilent 1260系列液相色譜儀、紫外檢測器,色譜柱為C18柱,流動相為98% (V∶V)甲醇,等度洗脫,進樣量10 μL,柱溫30℃,流速1 mL/min,檢測波長250 nm。
兒茶素檢測條件:儀器設備為Agilent 1260系列液相色譜儀、紫外檢測器,色譜柱為C18柱,流動相為95% (V∶V)的甲醇,等度洗脫,進樣量10 μL,柱溫35°C,流速1 mL/min,檢測波長254 nm。
蔗糖磷酸化酶的糖苷轉化率=(實驗組底物減少量/對照組底物的量)×100%。
2 結果與分析
2.1 生物信息學分析及基因的克隆
經SMART Main page分析發現,其N端23-382位氨基酸為α-淀粉酶結構域[圖1(a)]。結合SignalP 4.1分析,結果顯示重組酶Cbsp不含信號肽[圖1(b)]。以丁酸梭菌的基因組DNA為模板,利用引物cbsp-F、cbsp-R進行PCR擴增目的基因,得到一條大小約為1.5 kb的特異性條帶,測序正確后將其命名為cbsp[圖1(c)]。
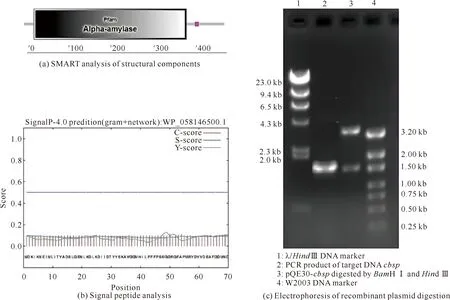
圖1 蔗糖磷酸化酶生物信息學分析及重組質粒酶切電泳圖
2.2 重組菌株的誘導表達與純化
如圖2所示,含有重組質粒的菌株和重組酶Cbsp純化物在約55 kDa大小處均有明顯的蛋白質特征條帶,與理論分子質量一致,可進行下一步測定。
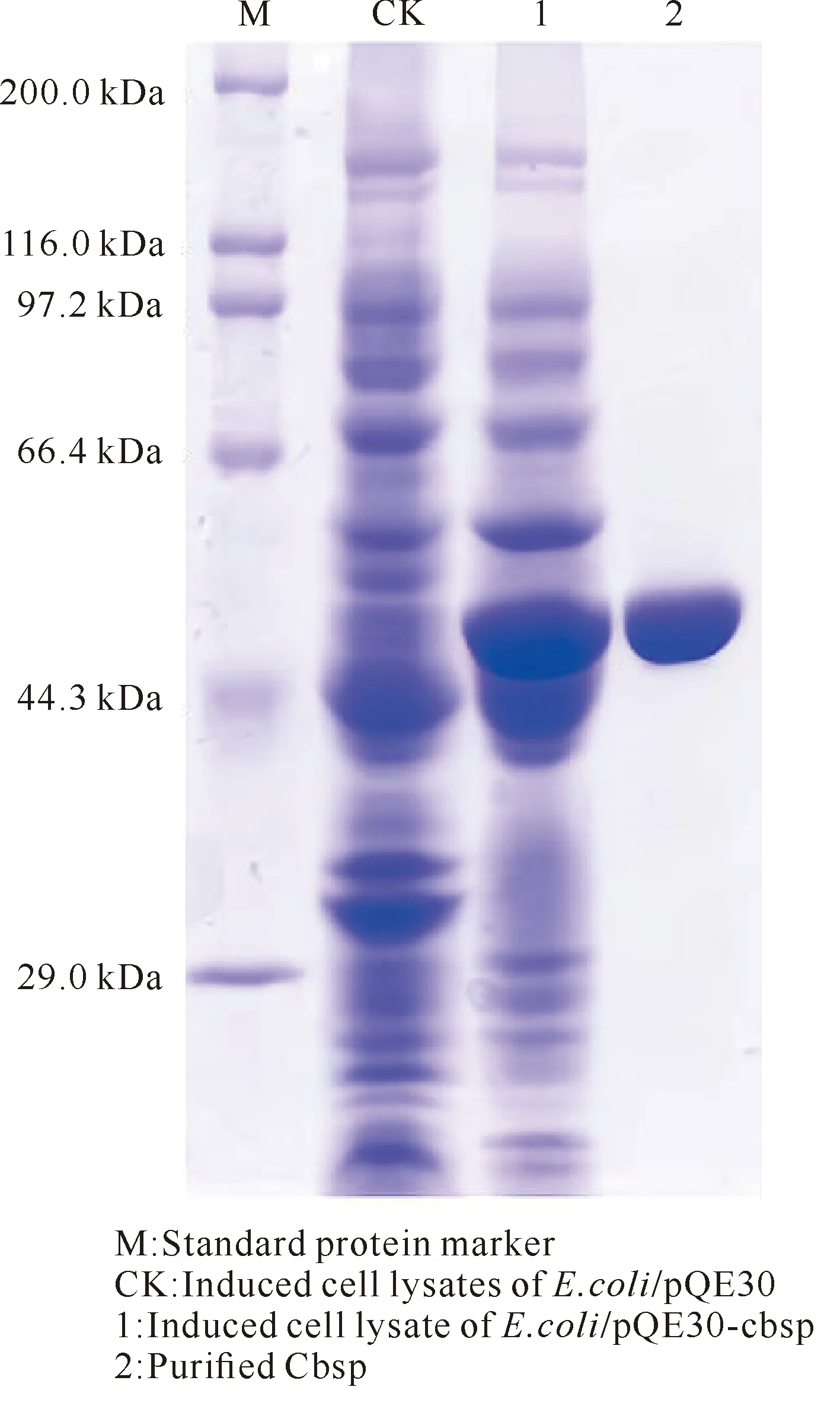
圖2 Cbsp的SDS-PAGE分析
2.3 重組酶Cbsp的酶學性質分析
2.3.1 溫度和pH值對酶活力的影響
如圖3(a)所示,Cbsp的最適pH值為7.0,在 pH值為6.0-7.5時可以保持60%以上的相對酶活力,當pH值大于7.5時,Cbsp酶活力急劇下降;當pH值達到8.0時,Cbsp僅剩20%左右的相對酶活力。如圖3(b)所示,Cbsp的最適溫度為45℃,在35-50℃時能保持40%以上的相對酶活力,當溫度大于55℃時則完全失活,說明Cbsp熱穩定性不佳,會對后續相關功能的研究造成一定的影響。如圖3(c)(d)所示,在35℃、40℃和4℃條件下保存36 h,Cbsp的相對酶活力仍然可以保持在80%以上,在45℃條件下保存42 h后,酶活力完全喪失,半衰期為210.4 min;在50℃保存0.5 h相對酶活力僅殘余20%,1 h后酶活力完全失去,半衰期為9.57 min。如圖3(d)所示,Cbsp在pH值為6.5-9.0時酶活力較為穩定,能夠保持80%以上的相對酶活力,表現出較好的堿耐受性。
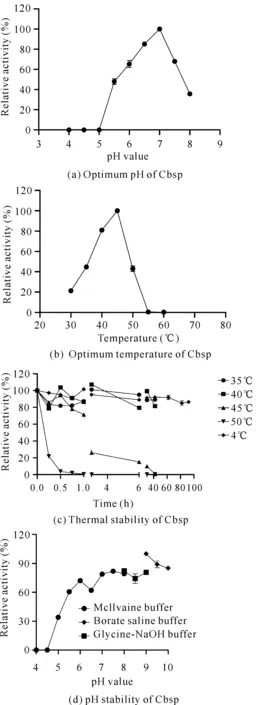
圖3 pH值和溫度對Cbsp的影響
2.3.2 重組酶Cbsp的Km和Vmax
利用軟件GraphPad Prism 5,通過雙倒數作圖法擬合出Cbsp的Km為(62.4±4.749) mmol·L-1,Vmax為(957±23.29) μmol·mg-1·min-1,如圖4所示。
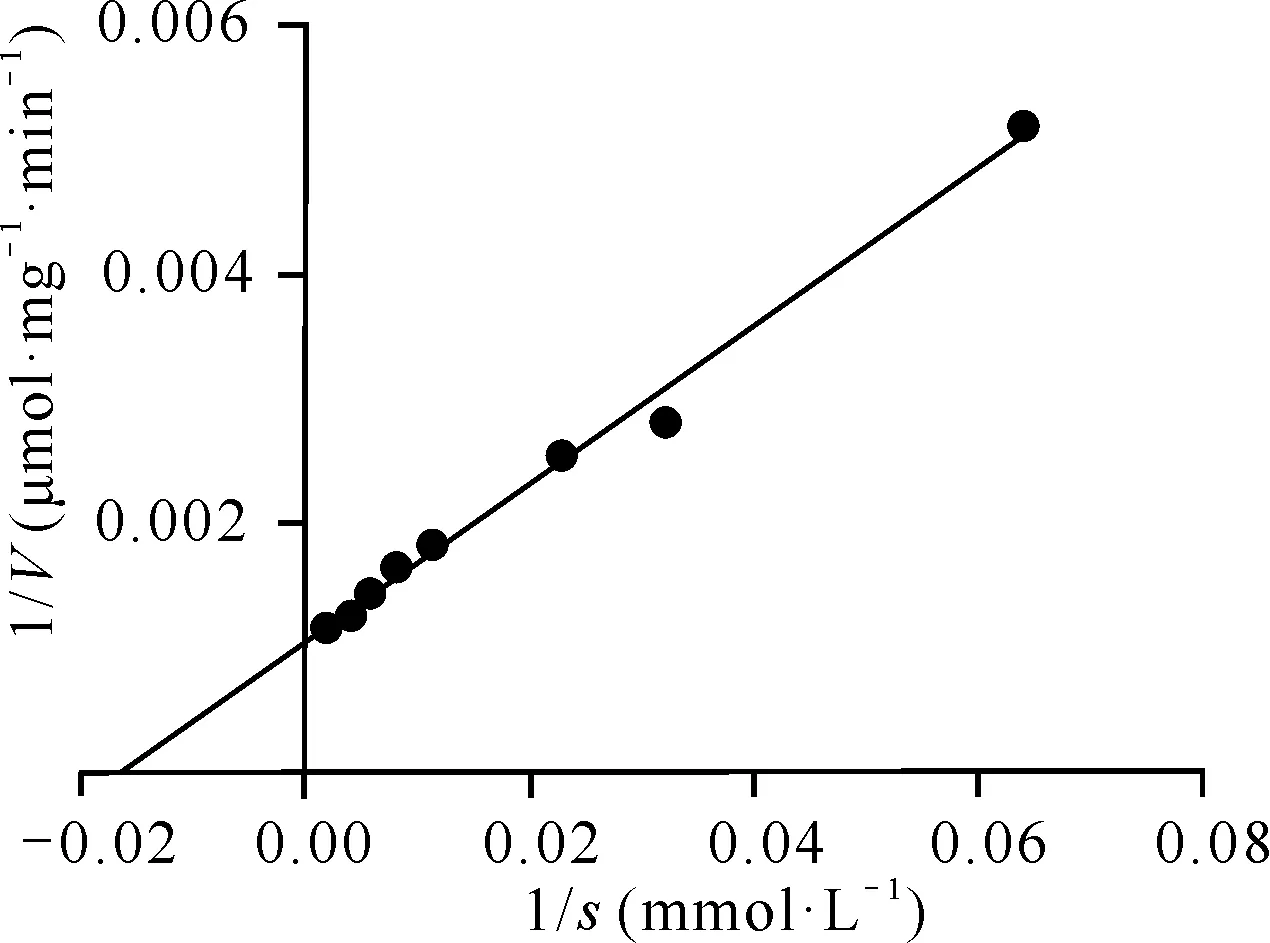
圖4 Cbsp的Km和Vmax
2.3.3 化學試劑對酶活力的影響
如圖5:(a)-(c)所示,除了低濃度的DMSO和甲醇對酶活力有輕微的促進作用外,其他一元醇、二元醇和有機試劑都不同程度地抑制酶活力,并且抑制強度隨有機試劑濃度的增大而增強。如圖5(d)所示,1% Triton X-100、5% Tween-80和20 mmol·L-1DTT 對于酶活力具有促進作用,其中20 mmol·L-1DTT可明顯提高酶活力;0.05% SDS和4 mol·L-1脲則使酶完全失去活力。由此可知,后續研究可以通過試劑來增強重組酶的酶活力,促進底物的溶解,進而提高其轉化效率。
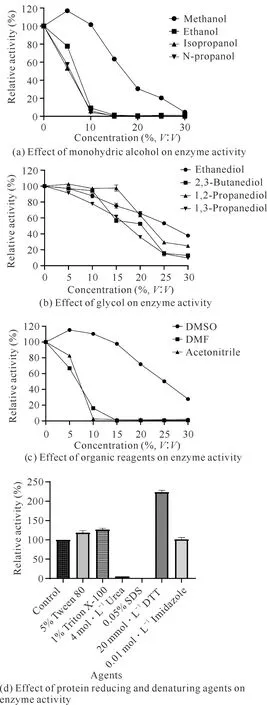
圖5 化學試劑對于Cbsp的影響
2.4 轉糖苷功能的測定
以終濃度15%的G-1-P為糖基供體時,Cbsp對D-阿拉伯糖、L-阿拉伯糖、D-果糖、D-半乳糖、D-甘露糖、L-山梨糖、木糖醇、1,2-丙二醇、1,3-丙二醇、2,3-丁二醇、乙二醇、丙三醇、L-抗壞血酸、(-)-兒茶素和己基-β-D-吡喃葡萄糖苷都具有轉糖苷活性;以終濃度8%的蔗糖為供體時,Cbsp僅對己基-β-D-吡喃葡萄糖苷和(-)-兒茶素有轉糖苷活性。其中以終濃度8%的蔗糖為糖基供體時,Cbsp對終濃度1%的己基-β-D-吡喃葡萄糖苷具有較高的轉糖苷能力,轉糖苷產物檢測結果如圖6所示。從圖6可知,以終濃度8%的蔗糖為糖基供體時,Cbsp對己基-β-D-吡喃葡萄糖苷的轉糖苷效率為42.0%[圖6(a)箭頭所指為轉糖苷產物]。而當以終濃度15%的G-1-P為糖基供體時,己基-β-D-吡喃葡萄糖苷的轉化率為5.5%[圖6(b)箭頭所指為轉糖苷產物],遠遠低于以蔗糖為糖基供體時的轉化效率。另外,Cbsp只有在以蔗糖為糖基供體時才對(-)-兒茶素具有轉糖苷活性,其結果如圖7所示。從圖7可以看出Cbsp催化(-)-兒茶素轉糖苷生成了兒茶素葡萄糖苷[圖7(a)箭頭所指],經計算其轉化率約為6.8%。
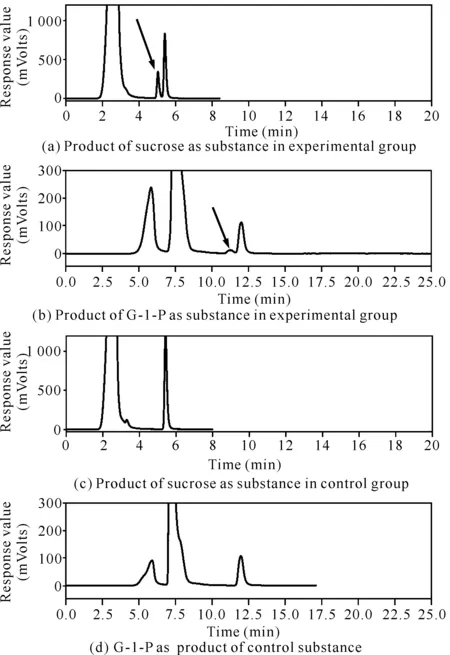
圖6 以己基-β-D-吡喃葡萄糖苷為受體時Cbsp的轉糖苷產物分析

圖7 以-(-)兒茶素為受體時Cbsp的轉糖苷產物分析
3 討論
如今,隨著蔗糖磷酸化酶的應用愈加廣泛,對于其性能的研究需求也越來越大。何賀賀等[19]研究發現來自腸膜明串珠菌Leuconostocmesenteroides的蔗糖磷酸化酶的最適反應溫度為37℃,來源于BacillusmegateriumNCIB 8508的蔗糖磷酸化酶最適反應溫度為50℃。來自Bifidobacteriumlongum的蔗糖磷酸化酶在20-50℃時,相對酶活性為98%以上;在60-90℃時,相對酶活性為70%以上[18]。本研究中的重組酶Cbsp最適反應溫度是45℃,在40℃以下保存36 h時酶只有80%的相對活性,一旦溫度上升到50℃且只保存1 h酶便會完全失活。這些研究不僅表明了不同的來源會造成酶活力的差異,而且還說明了大多數的蔗糖磷酸化酶都不是嗜熱型酶。
本研究中的Cbsp與Kitao等[20]報道的來自腸膜明串珠菌的蔗糖磷酸化酶相比,都對部分單糖及糖醇類具有受體特異性,有利于對人體健康十分有幫助的益生元低聚糖如曲二糖等的合成,這些益生元通過蔗糖磷酸化酶轉糖苷功能來催化合成,極大地降低了工業成本,帶來非常大的經濟效益。另外,本研究中的重組酶Cbsp還對烷基糖苷類物質己基-β-D-吡喃葡萄糖苷具有42.0%的轉糖苷效率,能夠有效地發生轉糖苷反應,得到的產物可以作為一類具有低刺激度、可生物降解、毒性低、無污染等優良性狀的新型的非離子表面活性劑,在食品、洗滌劑、農藥和醫療等領域展現出廣闊的應用前景[21]。
本研究的重組酶Cbsp雖對(-)-兒茶素具有轉糖苷活性,但轉化率僅有6.8%,而且部分受體底物無法發生轉糖苷反應,可能是由于受體親和力太低,無法與反應系統中的水分子等其他物質競爭結合位點。De Winter等[22]利用液態雙相體系進行糖基化轉換,大大提高了轉糖苷效率,證明了親和力低會影響糖基化反應。因此,水相系統、非水相系統和微水相系統等不同的反應系統對轉糖苷反應有明顯的影響。下一步可改善反應體系,以提高轉糖苷活性。
4 結論
本研究克隆了來源于丁酸梭菌DSM10702中的蔗糖磷酸化酶基因cbsp,并在感受態細胞E.coliXL1-blue中成功實現了異源表達。重組酶蛋白分子質量大小約為55 kDa。以蔗糖為底物時,蔗糖磷酸化酶最適反應溫度為45℃,最適pH值為7.0,在35-45℃、pH值6.0-9.0條件下能保持酶活力穩定;Km為(62.4±4.749) mmol·L-1,Vmax為(957±23.29) μmol·mg-1·min-1。當G-1-P為糖基供體時,Cbsp對于大部分單糖及糖醇類受體表現出轉糖苷活性,對(-)-兒茶素、己基-β-D-吡喃葡萄糖苷和L-抗壞血酸表現出一定程度的轉糖苷活性。對來自丁酸梭菌的蔗糖磷酸化酶的酶學性質的研究,為蔗糖磷酸化酶相關數據庫增添了新的內容,但相比之下,重組酶Cbsp相對較弱的熱穩定性十分不利于工業應用,未來可加強這方面的研究改造。
