蠋蝽成蟲對兩種煙草害蟲卵及3齡幼蟲的捕食功能反應
2022-02-10 23:33:07楊燦母銀林汪潔黃純楊張長華喻會平陳祥盛
植物保護 2022年1期
楊燦 母銀林 汪潔 黃純楊 張長華 喻會平 陳祥盛
摘要
為探究蠋蝽對兩種煙草害蟲煙青蟲和斜紋夜蛾的捕食能力,在室內可控條件下開展了蠋蝽雌雄成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食試驗,利用HollingⅡ功能模型擬合蠋蝽雌雄成蟲對煙青蟲和斜紋夜蛾的捕食功能反應。結果表明,蠋蝽雌成蟲對煙青蟲和斜紋夜蛾的瞬時攻擊率為:煙青蟲3齡幼蟲(1.056)>斜紋夜蛾卵(1.055)>煙青蟲卵(1.035)>斜紋夜蛾3齡幼蟲(0.973);控害效應高低為:煙青蟲卵(152.21)>斜紋夜蛾卵(124.18)>煙青蟲3齡幼蟲(15.09)>斜紋夜蛾3齡幼蟲(9.63);蠋蝽雄成蟲對煙青蟲和斜紋夜蛾的瞬時攻擊率為:煙青蟲卵(1.060)>斜紋夜蛾卵(1.056)>煙青蟲3齡幼蟲(0.825)>斜紋夜蛾3齡幼蟲(0.760);控害效應高低順序為:斜紋夜蛾卵(111.16)>煙青蟲卵(101.92)>煙青蟲3齡幼蟲(8.87)>斜紋夜蛾3齡幼蟲(7.04)。隨著煙青蟲和斜紋夜蛾密度的增加,蠋蝽成蟲的搜尋效應會逐漸下降。研究結果表明蠋蝽雌雄成蟲對煙青蟲和斜紋夜蛾具有良好的防控能力,因此,可結合害蟲預測預報,在煙青蟲和斜紋夜蛾產卵高峰期和3齡幼蟲發生前期大量釋放蠋蝽成蟲以降低兩種害蟲的種群數量,減少其對煙草的危害。
??關鍵詞
蠋蝽;煙青蟲;斜紋夜蛾;生物防控;捕食功能反應
中圖分類號:
S476
文獻標識碼:A
DOI:10.16688/j.zwbh.2020678
Predatory functional responses of Arma chinensis (Fallou) to the eggs and 3rdinstar larvae of two tobacco pests
YANG Can1,2,MU Yinlin1,2,WANG Jie1,2,HUANG Chunyang3,
ZHANG Changhua3,YU Huiping4,CHEN Xiangsheng1,2*
(1. Institute of Entomology, Guizhou University, Guiyang550025, China; 2. Provincial Special Key Laboratory
for Development and Utilization of Insect Resources, Guizhou University, Guiyang550025, China;
3. Zunyi Branch Company of Guizhou Tobacco Company, Zunyi563300, China; 4. Technology
Center, Bijie Branch Company of Guizhou Tobacco Company, Bijie551700, China)
Abstract
To explore the predatory potential of Arma chinensis against Heliothis assulta and Spodoptera litura, the predation of the eggs and 3rdinstar larvae of H. assulta and S.litura by A.chinensis adults was measured under controlled conditions in the laboratory. The predatory functional responses of A.chinensis adults fitted the Holling Ⅱ function model. The instantaneous attack rates of A.chinensis female adults to the pests from high to low were: the 3rdinstar larvae of H.assulta (1.056), eggs of S.litura (1.055), eggs of H.assulta (1.035), the 3rdinstar larvae of S.litura (0.973). The predation capacities from high to low were: eggs of H.assulta (152.21), eggs of S.litura (124.18), 3rdinstar larvae of? H.assulta (15.09) and 3rdinstar larvae of S.litura (9.63). The instantaneous attack rates of A.chinensis male adults to the pests from high to low were: eggs of H.assulta (1.060), eggs of S.litura (1.056), 3rdinstar larvae of H.assulta (0.825), 3rdinstar larvae of S.litura (0.760). The predation capacities from high to low were: eggs of S.litura (111.16), eggs of H.assulta (101.92), 3rdinstar larvae of H.assulta (8.87), and 3rdinstar larvae of S.litura (7.04). With increasing densities of H.assulta and S.litura, the searching efficiency of A.chinensis adults decreased gradually. This study suggested that A.chinensis adults had a good control ability against H.assulta and S.litura. Combined with pest forecast and prediction, releasing a large number of A.chinensis adults during the oviposition peak period and occurrence prophase of 3rdinstar H.assulta and S.litura can help reduce the pest population size and decrease its damage to tobacco.
Key words
Arma chinensis;Heliothis assulta;Spodoptera litura;biological? control;predatory functional response
煙草是一種經濟效益非常顯著的經濟作物,我國是世界上最大的烤煙產區之一[1]。斜紋夜蛾Spodoptera litura (Fabricius)和煙青蟲Heliothis assulta (Guenée) 是為害煙草的兩種重要害蟲,每年給煙草生產造成較大的經濟損失[2 -4]。
煙青蟲是一種重要的寡食性農業害蟲,主要為害煙草、辣椒等茄科植物[5]。在煙田,幼蟲取食煙草心芽及嫩葉,偶爾蛀食煙莖,使煙株整株死亡[6],特別是3齡后幼蟲進入暴食期,還具有轉移為害的特性。資料顯示,各大煙區每年因該蟲造成的煙草產量損失達5%~10%, 在大發生年份可超過15%[3,7- 9]。斜紋夜蛾是一種全世界范圍分布并且具有間歇性暴發特點的廣食性農業害蟲,可為害100多科389種植物[10 -11]。斜紋夜蛾主要以幼蟲為害寄主植物。在煙葉旺長期 采收期幼蟲啃食中、下部葉片及葉柄,進入3齡后因暴食性可將煙葉吃光只剩稈,若氣候與食物適宜,能在短時間內造成不可逆的嚴重危害[12]。目前,“以蟲治蟲”的生物防治方法已逐漸成為煙草生產中害蟲防治的主要趨勢,蠋蝽等捕食性天敵昆蟲在煙草害蟲防治中的利用逐步為人們所重視。
蠋蝽Arma chinensis (Fallou)隸屬于半翅目Hemiptera,蝽科Pentatomidae,益蝽亞科Asopinae,是一種捕食性天敵昆蟲,可以捕食鱗翅目、鞘翅目、膜翅目及半翅目等多個目的害蟲[13 -15]。而在飼養技術[16]以及人工飼料[17 -19]等方面的研究使得蠋蝽已經可以實現規模化擴繁。目前全國已有多個基地可持續為生物防治提供蠋蝽。研究表明蠋蝽對斜紋夜蛾幼蟲具有良好的防控作用[12, 20 -21],而其對斜紋夜蛾卵及對煙青蟲的捕食情況目前未見報道。
斜紋夜蛾與煙青蟲的3齡期是一個暴發與轉移為害的關鍵期,因此本試驗幼蟲蟲態選擇為3齡,通過擬合蠋蝽雌雄成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食功能反應,探索其搜尋效應,以填補蠋蝽對煙青蟲的捕食參數的空白,了解蠋蝽對兩種煙草害蟲的捕食能力,以期為利用蠋蝽防治這兩種煙草害蟲提供參考。
1材料與方法
1.1供試蟲源
蠋蝽由貴州省甕安縣天敵昆蟲繁育基地提供;斜紋夜蛾和煙青蟲采自貴州省甕安縣天文鎮煙田(107.47°E,27.32°N),用煙草上二部以上去頂的煙花與煙葉進行擴繁備用。蠋蝽、煙青蟲和斜紋夜蛾都飼養于人工氣候箱(ARMA580),飼養條件:溫度為(26±0.5)℃,光周期為L∥D=16 h∥8 h,濕度(RH)為(75±5)%。
1.2試驗方法
試驗前將羽化48 h以內的蠋蝽雌雄成蟲放置于捕食盒中(長21.2 cm、寬14.3 cm、高9 cm),每盒放置1頭蠋蝽和一塊8 cm2的水浸脫脂棉,同時將煙青蟲或斜紋夜蛾幼蟲放入同等規格的獵物盒中,卵則放入培養皿(直徑8 cm、高1.2 cm)中,后將培養皿放入捕食盒內,待蠋蝽饑餓24 h后轉移到獵物盒中。以幼蟲作為獵物時,試驗盒中放入一片煙葉避免其自相殘殺。煙青蟲和斜紋夜蛾卵密度設置為:20、40、60、80、100粒/盒,3齡幼蟲密度設置為:5、10、15、20、25頭/盒。24 h后檢查獵物死亡情況,用鑷子撥動幼蟲,若其無反應、不活動則判定為死亡,若卵變為空殼則判定為被取食。每個處理5次重復。試驗在溫度(26±0.5)℃,光周期L∥D=16 h∥8 h,RH(70±5)%的條件下進行。
1.3數據分析
捕食功能反應方程:Na=aNTr/(1+aThN)[22],搜尋效應:S=aTr/(1+aThN)[23]。控害效應:a/Th。
式中Na:捕食獵物數,a:捕食者對獵物的瞬時攻擊率,N:獵物密度,Tr: 試驗持續時間(試驗設置為24 h即Tr=1 d),Th:處理獵物時間(捕食者處理一頭獵物所需要的時間),S:搜尋效應。
所得數據先用Excel 2010記錄與處理,再利用SPSS 19.0擬合功能反應方程,最后用Graphpad Prism 8.3.0作圖。
2結果與分析
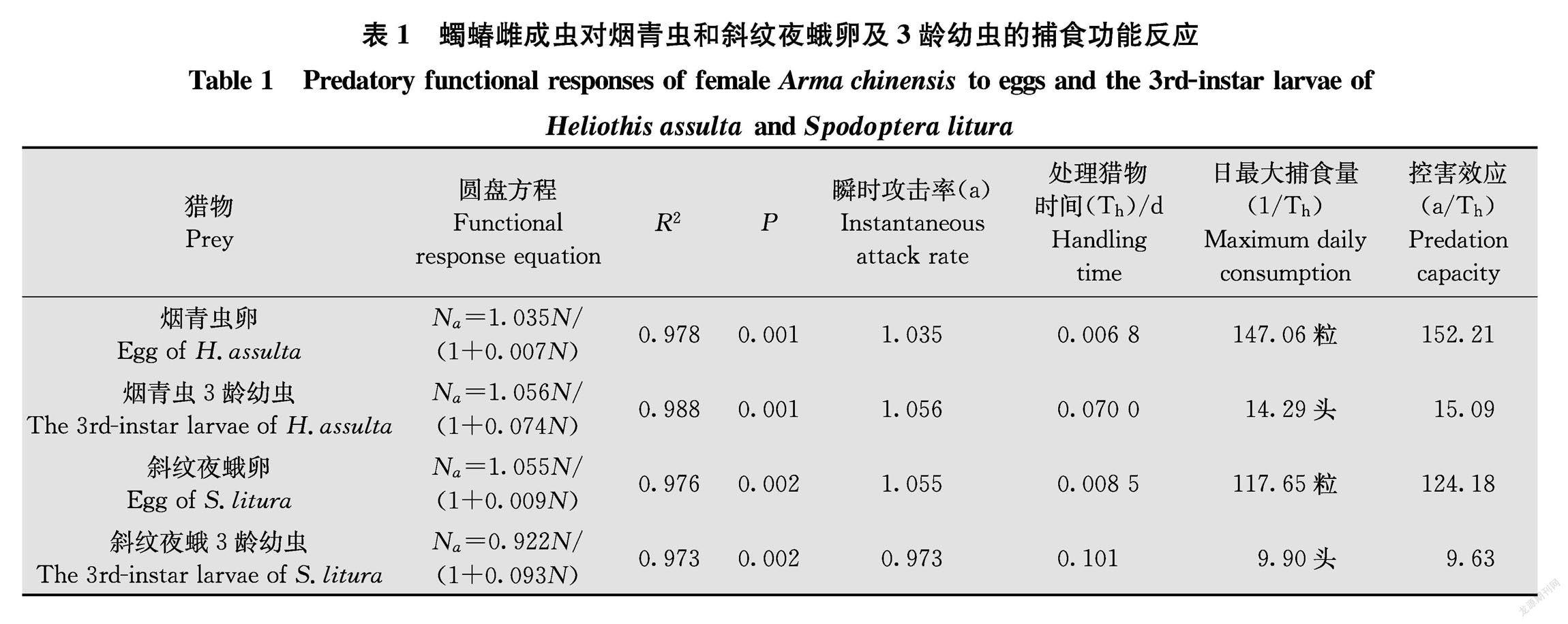
2.1蠋蝽雌成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食功能反應
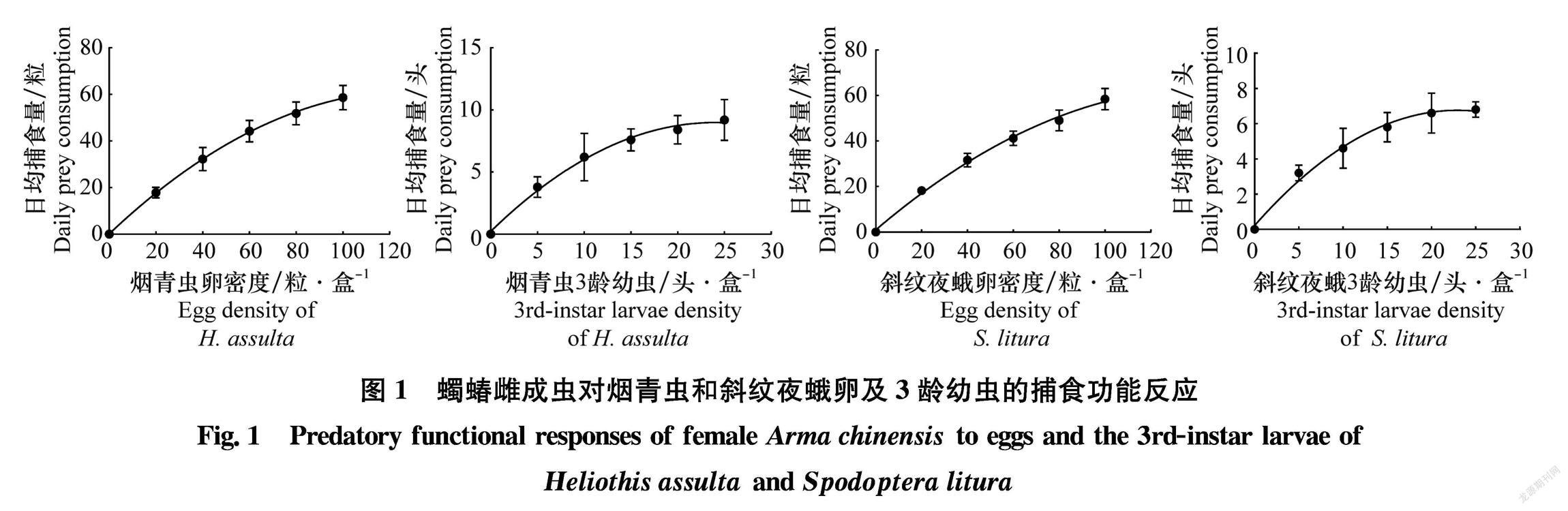
蠋蝽雌成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食功能反應均符合HollingⅡ模型(圖1),且P值都遠小于0.01(表1),隨著獵物密度的增加,蠋蝽雌成蟲的捕食量先上升后逐漸趨于穩定。雌成蟲對煙青蟲和斜紋夜蛾瞬時攻擊率表現為:煙青蟲3齡幼蟲(1.056)>斜紋夜蛾卵(1.055)>煙青蟲卵(1.035)>斜紋夜蛾3齡幼蟲(0.973);日最大捕食量為:煙青蟲卵147.06粒,煙青蟲3齡幼蟲14.29頭,斜紋夜蛾卵117.65粒,斜紋夜蛾3齡幼蟲9.90頭;控害效應為:煙青蟲卵(152.21)>斜紋夜蛾卵(124.18)>煙青蟲3齡幼蟲(15.09)>斜紋夜蛾3齡幼蟲(9.63)。其擬合度為:煙青蟲3齡幼蟲(R2=0.988)>煙青蟲卵(R2=0.978)>斜紋夜蛾卵(R2=0.976)>斜紋夜蛾3齡幼蟲(R2=0.973)。
2.2蠋蝽雌成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的搜尋效應
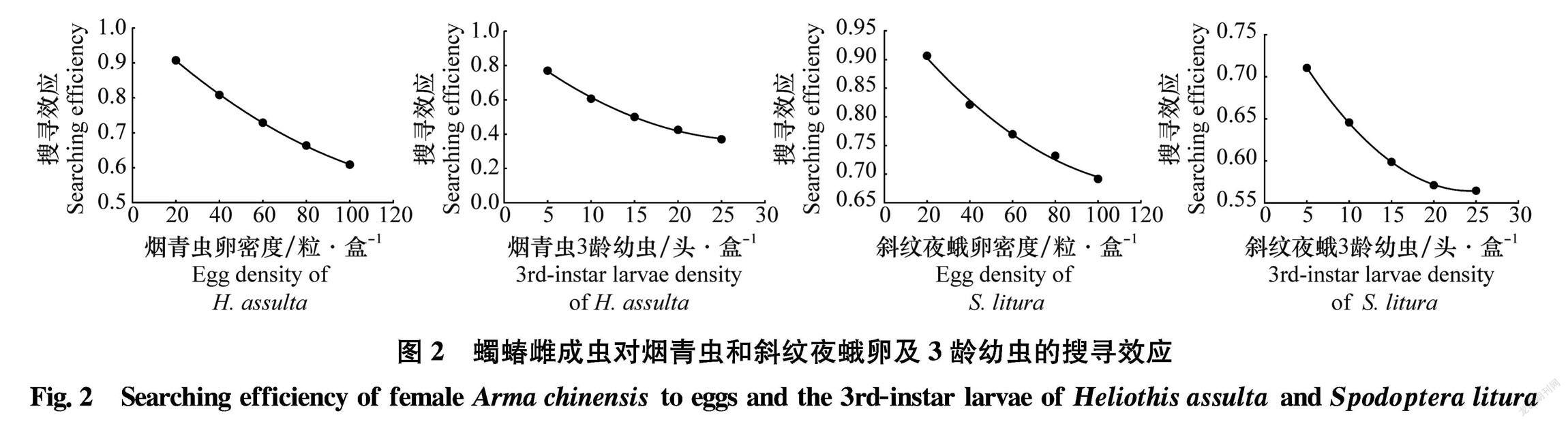
蠋蝽雌成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的 搜尋效應均隨著獵物密度的增加而不斷降低(圖2)。 初始時,蠋蝽雌成蟲對害蟲的搜尋效應最高,順序表現為:煙青蟲卵(0.908)> 斜紋夜蛾卵(0.906)> 煙青蟲3齡幼蟲(0.771)>斜紋夜蛾3齡幼蟲(0.711),當獵物密度達到最大時,蠋蝽雌成蟲對其搜尋效應都降到最低值,順序表現為:斜紋夜蛾卵(0.692)> 煙青蟲卵(0.609)>斜紋夜蛾3齡幼蟲(0.565)>煙青蟲3齡幼蟲(0.371)。
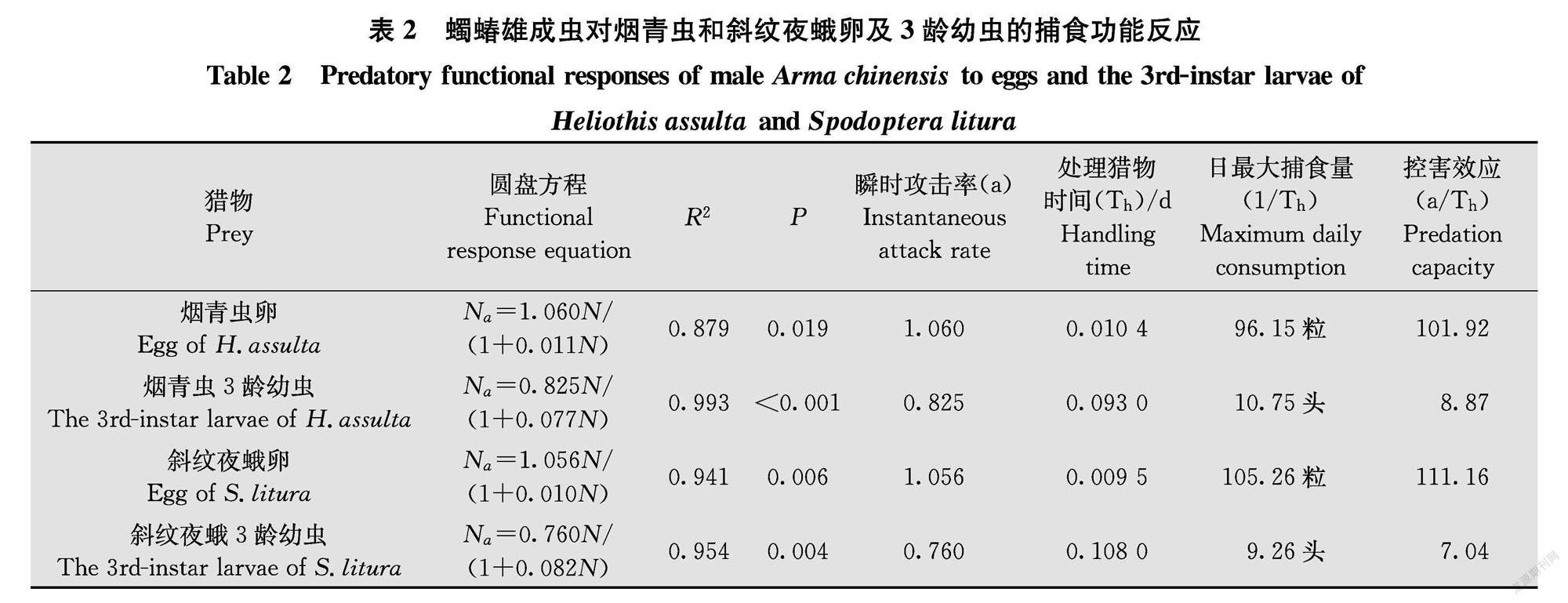
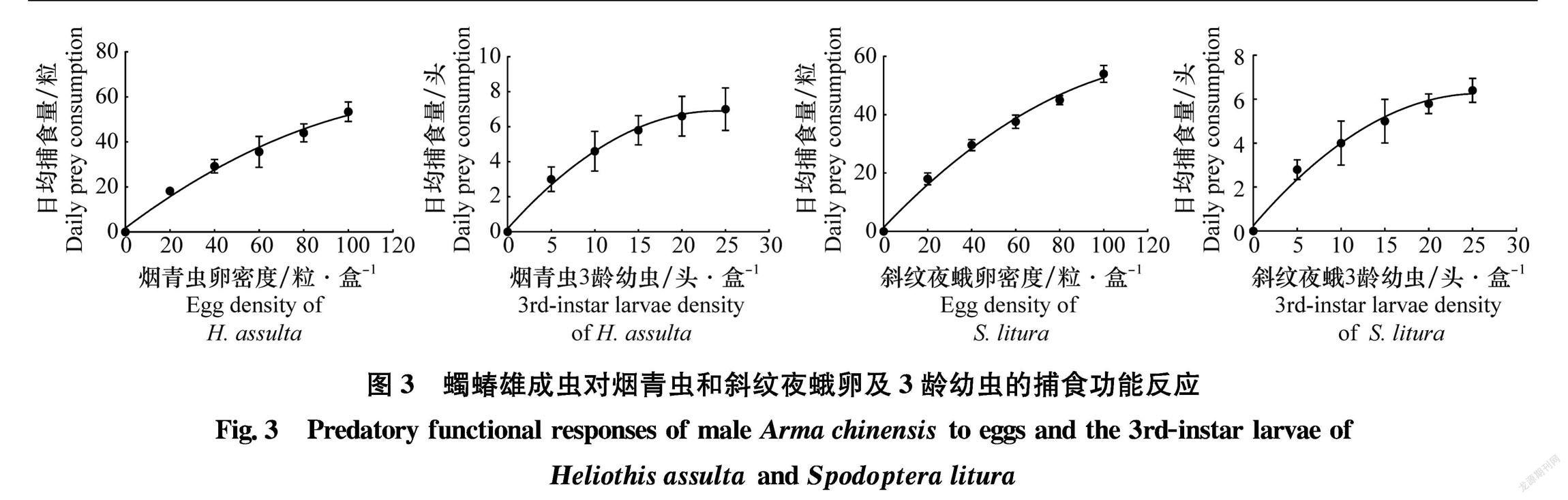
2.3蠋蝽雄成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食功能反應
蠋蝽雄成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食功能反應均符合HollingⅡ模型(圖3),且P值都遠小于0.05(表2),隨著獵物密度的增加,蠋蝽雄成蟲的捕食量同樣呈先上升后逐漸趨于穩定的趨勢。蠋蝽雄成蟲對煙青蟲和斜紋夜蛾瞬時攻擊率表現為:煙青蟲卵(1.060)>斜紋夜蛾卵(1.056)>煙青蟲3齡幼蟲(0.825)>斜紋夜蛾3齡幼蟲(0.760);日最大捕食量為:煙青蟲卵96.15粒,煙青蟲3齡幼蟲10.75頭,斜紋夜蛾卵105.26粒,斜紋夜蛾3齡幼蟲9.26頭;控害效應為:斜紋夜蛾卵(111.16)>煙青蟲卵(101.92)>煙青蟲3齡幼蟲(8.87)>斜紋夜蛾3齡幼蟲(7.04)。其擬合度表現為煙青蟲3齡幼蟲(R2=0.993)>斜紋夜蛾3齡幼蟲(R2=0.954)>斜紋夜蛾卵(R2=0.941)>煙青蟲卵(R2=0.879)。
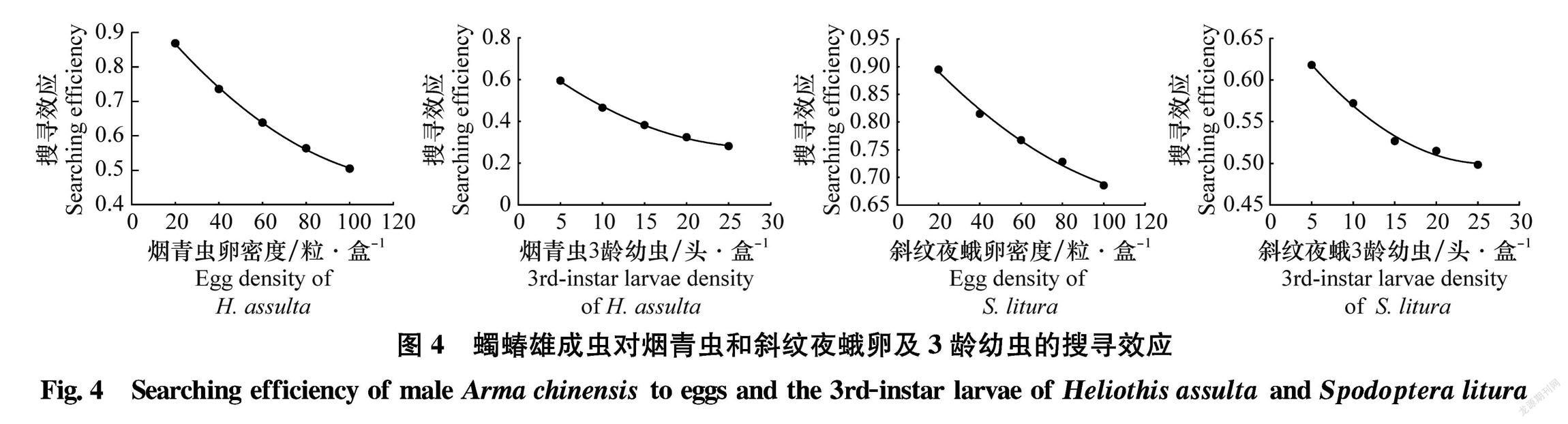
2.4蠋蝽雄成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的搜尋效應
蠋蝽雄成蟲對煙青蟲和斜紋夜蛾的搜尋效應同樣隨著獵物密度的增加而不斷降低(圖4)。初始時,蠋蝽雄成蟲對兩種害蟲的搜尋效應最高,順序表現為:斜紋夜蛾卵(0.895)>煙青蟲卵(0.869) > 斜紋夜蛾3齡幼蟲(0.618)>煙青蟲3齡幼蟲(0.596),當煙青蟲和斜紋夜蛾密度達到最大時,蠋蝽雄成蟲對其搜尋效應都降到最低值,其順序表現為:斜紋夜蛾卵(0.686)> 煙青蟲卵(0.505)>斜紋夜蛾3齡幼蟲(0.498)>煙青蟲3齡幼蟲(0.282)。?
3討論
捕食功能反應是測試天敵昆蟲能否捕食害蟲以及對害蟲控害能力的重要依據。本研究開展了蠋蝽雌雄成蟲對煙青蟲和斜紋夜蛾卵及3齡幼蟲的捕食功能反應試驗,明確了蠋蝽分別對兩種煙草害蟲的捕食能力,為進一步利用蠋蝽防治斜紋夜蛾或煙青蟲提供參考數據。
試驗結果顯示,蠋蝽雌雄成蟲對煙青蟲或斜紋夜蛾卵與3齡幼蟲都具有捕食能力,且符合HollingⅡ功能模型,這與其他捕食性天敵對獵物的捕食功能模型一致[24 -27]。其中,除了蠋蝽雌成蟲對煙青蟲3齡幼蟲的瞬時攻擊率高于煙青蟲卵外,蠋蝽雌成蟲捕食斜紋夜蛾,蠋蝽雄成蟲捕食斜紋夜蛾和捕食煙青蟲中,均表現為蠋蝽對獵物卵的瞬時攻擊率高于其3齡幼蟲。可能由于卵是靜止蟲態,所以被蠋蝽優先取食,這與天敵麗草蛉Chrysopa formosa Brauer捕食草地貪夜蛾低齡幼蟲與卵的研究結果相似[28]。 此外,蠋蝽雌成蟲對煙青蟲或斜紋夜蛾的捕食量均高于雄成蟲,說明雌成蟲對煙青蟲或斜紋夜蛾具有更強的捕食能力,這與體型較大的捕食者會因交配產卵等需求而取食更多獵物的觀點相吻合[29]。
搜尋效應是天敵在捕食獵物過程中的行為反應。本試驗中,蠋蝽成蟲對煙青蟲和斜紋夜蛾的搜尋效應均隨著獵物密度的增加而降低,這與蠋蝽捕食其他獵物的研究結果相一致[12, 21, 30]。另外,捕食功能反應曲線和搜尋效應曲線顯示,當煙青蟲和斜紋夜蛾卵與3齡幼蟲密度逐漸增加時,蠋蝽雌雄成蟲對其捕食速率緩慢降低,而搜尋效率則由最高值降到最低值,這說明在煙青蟲和斜紋夜蛾種群密度較低時,蠋蝽能夠發揮更高效的控害作用。
研究結果表明,蠋蝽成蟲對煙青蟲或斜紋夜蛾卵與幼蟲均具有一定的捕食能力,其中蠋蝽雌雄成蟲對煙青蟲和斜紋夜蛾卵的控害效應最好,在兩種害蟲卵及3齡幼蟲密度較低時防控效果較好。實際應用時可結合害蟲預測預報,在煙青蟲和斜紋夜蛾進入為害初期和產卵高峰期時大量釋放蠋蝽成蟲以抑制兩種煙草害蟲種群數量,降低為害。
本研究結果在限定的空間中獲得,一定程度上反映出蠋蝽對煙青蟲和斜紋夜蛾卵和3齡幼蟲的捕食能力,但在大田中,受環境因子的影響,蠋蝽對獵物的捕食效能可能會有所下降[20, 31]。 因此,蠋蝽在田間對兩種煙草害蟲的實際控害能力尚有待進一步研究。此外,由于捕食者對獵物的控制能力會受到捕食者對獵物捕食選擇性的直接影響[32],在煙青蟲和斜紋夜蛾幼蟲發生時期重疊時[33],蠋蝽是否對同時存在的兩種害蟲幼蟲具有選擇性,也值得進一步研究。
參考文獻
[1]吳紅波. 生物防治在我國煙草病蟲害防治上的應用[J]. 貴州農業科學, 2006, 34(S1): 103 -105.
[2]章士美, 邢健生, 胡文萍, 等. 六種多食性昆蟲食性的比較觀察[J]. 昆蟲學報, 1956, 6(1): 107 -122.
[3]惠豐立, 諸學英. 煙草夜蛾的主要生物學特性及生物防治[J]. 湖北農業學報, 2003(2): 61- 62.
[4]姚峰, 廖偉, 陳芝波, 等. 烤煙煙青蟲防治研究進展[J]. 安徽農業科學, 2015, 43(26): 110 -112.
[5]宋南, 李為爭, 董鈞鋒, 等. 煙青蟲的分類地位和起源地辨析[J]. 應用昆蟲學報, 2020, 57(3): 735 -743.
[6]黃國聯, 匡傳富, 譚琳, 等. 煙草煙青蟲生態防治研究進展[J]. 農業開發與裝備, 2015(9): 37 -38.
[7]王恩沛, 陶謙, 宋宏偉. 應用人工合成煙青蟲Heliothis assulta Guenée性信息素大面積誘蛾的研究[J]. 中國煙草學報, 1994, 2(2): 41 -45.
[8]趙兵, 田福海, 許濤, 等. 煙青蟲發生規律及測報因子研究[J]. 中國煙草, 1995(4): 34 -36.
[9]謝立群, 楊效文, 張效羲. 煙青蟲主要生物學特性及防治方法的研究現狀[J]. 煙草科技, 1997(2): 46 -47.
[10] 秦厚國, 汪篤棟, 丁建, 等. 斜紋夜蛾寄主植物名錄[J]. 江西農業學報, 2006, 18(5): 51 -58.
[11] AHMAD Y U, GHAFFAR A, RAFIQ M. Host plants of leaf worm, Spodoptera litura (Fabricius) (Lepidoptera: noctuidae) in Pakistan [J]. Asian Journal of Agriculture and Biology, 2013, 1(1): 23 -28.
[12]? 唐藝婷, 王孟卿, 李玉艷, 等. 蠋蝽對斜紋夜蛾幼蟲的捕食作用[J]. 中國煙草科學, 2020, 41(1): 62 -66.
[13] 潘淑琴, 楊顯山, 苑榮發, 等. 蠋敵捕食柳毒蛾數量測定[J]. 吉林林業科技, 1993(1): 25 26.
[14] 陳靜, 張建萍, 張建, 等. 蠋敵對雙斑長跗螢葉甲成蟲的捕食功能研究[J]. 昆蟲天敵, 2007, 29(4): 149 -154.
[15] 王文亮, 劉芹, 閆家河, 等. 美國白蛾新天敵 蠋敵捕食能力的初步觀察[J]. 山東林業科技, 2012(1): 11- 14.
[16] 潘明真, 張海平, 張長華, 等. 飼養密度和性比對蠋蝽存活和繁殖生物學特性的影響[J]. 中國生物防治學報, 2018, 34(1): 52 -58.
[17] ZOU Deyu, COUDRON T A, LIU Chenxi, et al. Nutrigenomic in Arma chinensis: transcriptome analysis of Arma chinensis fed on artificial diet and Chinese oak silkmoth Antheraea pernyi pupae [J/OL]. PLoS ONE, 2013, 8(4): e60881. DOI: 10.1371/journal.pone.0060881.
[18] ZOU Deyu, COUDRON T A, WU Huihui, et al. Performance and cost comparisons for continuous rearing of Arma chinensis(Hemiptera: Pentatomidae: Asopinae) on a zoophytogenous artificial diet and a secondary prey [J]. Journal of Economic Entomology, 2015, 108(2): 454-461.
[19] 廖平, 苗少明, 許若男, 等. 新型蠋蝽若蟲液體人工飼料效果評價[J]. 中國生物防治報, 2019, 35(1): 9 -14.
[20] 高強, 王迪, 張文慧, 等. 蠋蝽對斜紋夜蛾幼蟲的捕食作用研究[J]. 中國煙草科學, 2019, 40(6): 55- 59.
[21] 楊志浩, 孟玲, 李保平. 蟲齡對蠋蝽捕食斜紋夜蛾幼蟲行為參數的影響[J]. 生態學雜志, 2019, 38(11): 3376- 3381.
[22] HOLLING C S. Some characteristics of simple types of predation? and parasitism [J]. The Canadian Entomologist, 1959, 9l(7): 385 -398.
[23] 丁巖欽. 昆蟲數學生態學[M]. 北京: 科學出版社, 1994: 257 258, 303 304.
[24] 曹雯星, 張韜, 楊歡, 等. 大草蛉對草地貪夜蛾低齡幼蟲的捕食功能評價[J]. 植物保護學報, 2020, 47(40): 839- 844.
[25]? 侯崢嶸, 孫貝貝, 劉先建, 等. 大紅犀獵蝽對草地貪夜蛾3齡幼蟲捕食功能反應[J]. 植物保護學報, 2020, 47(4): 852- 858.
[26] 孫貝貝, 侯崢嶸, 董民, 等. 東亞小花蝽對草地貪夜蛾1齡幼蟲的捕食作用[J]. 植物保護學報, 2020, 47(4): 845 -851.
[27]? 李萍, 李玉艷, 向梅, 等. 大草蛉幼蟲對草地貪夜蛾低齡幼蟲的捕食能力評價[J]. 中國生物防治學報, 2020, 36(4): 513 -519.
[28] 李玉艷, 王孟卿, 張瑩瑩, 等. 麗草蛉幼蟲對草地貪夜蛾卵及低齡幼蟲的捕食能力評價[J]. 植物保護, 2021, 47(5): 178 -184.
[29] 巫厚長, 程遐年, 鄒運鼎. 不同饑餓程度的龜紋瓢蟲成蟲對煙蚜的捕食作用[J]. 應用生態學報, 2000, 11(5): 749- 752.
[30] 唐藝婷, 郭義, 潘明真, 等. 蠋蝽對小菜蛾幼蟲的捕食作用[J]. 植物保護, 2020, 46(4): 155- 160.
[31] 張曉軍, 張健, 孫守慧. 蠋蝽對榆紫葉甲的捕食作用[J]. 中國森林病蟲, 2016, 35(1): 13 -15.
[32] 吳圣勇. 栗真綏螨生物學及實驗種群生態學研究[D]. 北京: 中國農業科學院, 2011.
[33] 楊柳. 湖南煙區主要煙草害蟲發生動態監測及抗性機制研究[D]. 長沙:湖南農業大學, 2014.
收稿日期:2020 -12 -17修訂日期:2021 -03- 05
基金項目:
貴州省煙草公司遵義市公司科技項目(遵煙計[2018]11號);中國煙草總公司貴州省公司重大科技專項(中煙黔科[2019]10號)
* 通信作者
Email:chenxs3218@ 163.com
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學物理·高中(2016年12期)2017-04-22 11:53:03
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
聲屏世界(2014年6期)2014-02-28 15:18:09
中國煙草學報(2012年5期)2012-04-12 06:21:18
