自然增溫對南亞熱帶森林土壤微生物群落與有機碳代謝功能基因的影響
2022-02-12 08:38:32胡明慧趙建琪張慧玲褚國偉張德強
生態學報 2022年1期
胡明慧,趙建琪,王 玄,熊 鑫,張慧玲,褚國偉,孟 澤,張德強,*
1 中國科學院華南植物園, 廣州 510650 2 普洱學院, 普洱 665000 3 中國科學院大學, 北京 100049
自工業革命以來,大氣二氧化碳(CO2)濃度不斷攀升,隨之引起的全球變暖也已經成為不爭的事實[1]。IPCC報告指出,目前全球氣溫較工業革命前已經增加了1℃,預計在2030—2052年間全球氣溫將升高1.5℃[2]。反過來,氣溫上升又加速了土壤碳的排放[3]。土壤作為陸地生態系統最大的碳庫,其碳儲量超過了植被和大氣碳庫的總和,土壤碳庫的細微變化足以引起大氣中溫室氣體濃度的巨大波動[4]。因此,準確理解土壤碳循環過程對增溫的響應與適應機制,對合理預測未來全球變化背景下全球碳平衡格局的動態變化具有重要意義。
大量的研究表明,模擬增溫往往能提高土壤的呼吸速率,但其增加幅度因生態系統類型、增溫方式和幅度、增溫季節以及持續時間的不同而存在巨大差異[5]。也有研究指出,雖然增溫可以在短期內刺激土壤碳的排放,但隨著時間的延長這種促進作用會慢慢減弱,表現出一定的適應性[6];但對這種適應性的解釋,目前尚未達成共識[7]。根據熱動力學原理,化學結構不同的土壤有機碳各組分對溫度的敏感性也不同[8]。一般認為惰性組分較活性組分更難分解,對增溫的響應也更敏感[9—10]。然而,越來越多的證據顯示,土壤有機碳對增溫的敏感性不僅僅取決于其化學組成,礦物顆粒的物理化學保護、微生物的生理特性和群落組成可能發揮著更為重要的作用[11—12]。Allison等[13]認為土壤碳循環過程對增溫的響應主要依賴于土壤微生物的組成和活性,并提出在對氣候變化過程進行預測時必須考慮土壤微生物的反饋作用。
土壤微生物以土壤有機碳為主要碳源進行物質代謝,并通過分泌酶參與土壤有機碳的解吸附、解聚、溶解等分解過程[11]。增溫可以通過影響土壤微生物的生理功能,甚至改變微生物群落組成,從而刺激或抑制土壤碳的排放[14]。目前,關于增溫背景下土壤微生物在土壤碳循環過程中的作用已有大量研究。Zhang等[15]研究發現,增溫3年后土壤微生物生物量并沒有顯著變化,但真菌在群落中所占的比例較細菌有明顯提高,降低了土壤碳的排放(真菌的碳同化率較細菌更高)。但Frey等[16]在美國哈佛森林的增溫實驗結果表明,長期增溫(12年)后土壤微生物中真菌的豐度降低了。也有研究發現氣溫上升提高了真菌的活性,增強了微生物對土壤有機碳的分解代謝[17]。Bokhorst等[18]則發現增溫導致的土壤濕度降低抑制了土壤微生物的活性,從而減弱了微生物對土壤有機碳的代謝作用。另外,其他的一些研究指出,增溫對土壤微生物群落幾乎沒有影響[19—20]。顯然,因氣候條件、植被類型、土壤性質等的不同,不同研究中土壤微生物對增溫的響應與適應機制存在著差異。
受實驗條件的限制,目前大部分的模擬增溫實驗都是在室內開展的,其研究結果能否適用于野外自然狀況還難以定論。為數不多的野外研究也多集中于溫帶地區。與溫帶相比,熱帶和亞熱帶地區溫度的變化幅度較小,適合生物生存的閾值區間也相對較窄,其生態系統的結構和功能可能更容易受到氣候變化(特別是溫度變化)的影響[21]。因此,我們選擇在南亞熱帶地區的鼎湖山森林,將模擬的生態系統(含植物和土壤)從高海拔整體移位至低海拔地區形成自然增溫的效果。借助磷脂脂肪酸法(PLFAs)和宏基因組測序等手段,探究土壤微生物對增溫的響應與適應機制,研究結果將有助于合理預測全球變暖背景下南亞熱帶森林土壤碳庫的動態變化。
1 材料與方法
1.1 試驗地概況
研究樣地位于鼎湖山國家級自然保護區。保護區地屬廣東省肇慶市鼎湖區(23°09′N—23°11′N, 112°30′E—112°33′E),面積約1133 hm2。全年平均氣溫為20.9℃,最高氣溫達到38℃,最低氣溫為-0.2℃,年降雨量1927 mm,4—9月為主要降雨季節,降雨量約占全年的80%,干季(10—次年3月)降雨量占全年降雨量的20%。年平均蒸發量1115 mm,年平均相對濕度80.3%。鼎湖山自然保護區屬于低山丘陵地貌,構造線呈西南—東北走向,最高峰“雞籠山”海拔達到1000.3 m,坡度平均為30°—45°[22]。
不同海拔引起小氣候的變化,使得不同海拔梯度上的森林類型不同。海拔梯度自下而上分布主要森林類型分別有季風常綠闊葉林、針闊葉混交林和山地常綠闊葉林。季風常綠闊葉林優勢物種為木荷(Schimasuperba)、紅枝蒲桃(Syzygiumrehderianum)、短序潤楠(Machilusbreviflora)、紅錐(Castanopsishystrix)、海南紅豆(Ormosiapinnata)、九節(Psychotriaasiatica); 針闊葉混交林優勢物種為木荷(Schimasuperba)、紅枝蒲桃(Syzygiumrehderianum)、短序潤楠(Machilusbreviflora)、馬尾松(Pinusmassoniana)、紅錐(Castanopsishystrix)、山血丹(Ardisialindleyana);山地常綠闊葉林優勢物種為木荷(Schimasuperba)、紅枝蒲桃(Syzygiumrehderianum)、短序潤楠(Machilusbreviflora)、鼠刺(Iteachinensis)、密花樹(Myrsineseguinii)、山血丹(Ardisialindleyana)等[23]。
1.2 實驗設計
1.2.1樣地設置
增溫樣地建于2013年,在海拔600 m的山地常綠闊葉林和海拔30 m的季風常綠闊葉林,選擇空曠地帶,分別建造3個開頂箱(open-top chamber, OTC)。每個OTC方形坑長3 m、寬3 m、內徑深0.8 m,地下部分周圍用磚頭砌墻,墻高于地面0.2 m,以防止水和周圍土壤元素的橫向和垂直移動。OTC的頂部和底部各有一個出水孔,用來收集地表徑流的土壤滲透水。待修建完成,往OTC方形坑內填入土壤。土壤按照對應的層次(0—20、20—40、40—70 cm)收集,每個OTC方形坑內土壤量為6.3 m3。6個OTC內土壤均采自于山地常綠闊葉林的山地黃壤,所有土壤均按照對應的層次填埋于坑內。在海拔600 m的山地常綠闊葉林內選取年齡、基莖和樹高一致的優勢樹種苗木,在林內統一位置進行馴化后,移植到6個OTC內[22]。
本實驗通過降低海拔梯度來模擬增溫。共設置兩組OTC (OTC1、OTC2),每組設置3個重復。設置在海拔600 m的OTC1為對照樣地,設置在海拔30 m的OTC2為增溫樣地(3.0±1.0)℃。每個OTC樣地內種植植物包括木荷(Schimasuperba)、紅枝蒲桃(Syzygiumrehderianum)、短序潤楠(Machilusbreviflora)、鼠刺(Iteachinensis)、密花樹(Myrsineseguinii)、山血丹(Ardisialindleyana)[23]。
1.2.2樣品采集
分別于2017年12月(干季)和2018年6月(濕季)在上述OTC樣地進行兩次土壤樣品的采集。采用五點混合取樣法,每次采樣時選取較為分散的5個點混合成一個樣品,取樣前去除表層土壤的腐殖質、凋落物等雜質,每個OTC用內徑5 cm的土鉆取表層土0—10 cm,每個OTC采集3個土壤樣品。采集的土壤樣品立即放入裝有干冰的保溫箱保存,盡快帶回實驗室進行分析處理。挑除土壤中肉眼可見的根系和石礫后過2 mm篩。隨后將土壤樣品分為三份,一份放置陰涼處風干用于測定土壤基本理化性質;一份放置于冰箱內4℃保存,用于土壤微生物生物量以及群落結構組成的測定;一份放置于冰箱內-80℃保存,用于后續宏基因組測定(將同一海拔梯度的3個OTC樣地樣品混合成一份樣品用于宏基因組測序)。
1.3 樣品測定
1.3.1土壤理化性質
土壤溫、濕度數據來自OTC樣地觀測設備的溫、濕度傳感器(傳感器每隔一小時記錄一次土壤溫、濕度)。采用重鉻酸鉀氧化-硫酸亞鐵滴定法測定土壤有機碳;半微量凱氏定氮法測定土壤全氮,采用氫氟酸—高氯酸消煮—鉬銻抗比色法測定土壤全磷;土壤pH值的測定采用電極電位法,水土比為2.5:1;土壤硝態氮的測定采用紫外分光光度法;土壤銨態氮的測定采用靛酚藍比色法。上述理化性質的測定方法參照魯如坤[24]和鮑士旦[25]。
1.3.2土壤微生物群落組成測定
采用磷脂脂肪酸法(PLFAs)測定土壤不同種群微生物量,實驗步驟參照Bossio和Scow[26]的基礎上稍作改進。稱取8 g凍干土壤于35 mL離心管中,向離心管中加入5 mL磷酸緩沖液、6 mL三氯甲烷、12 mL甲醇震蕩提取后離心,離心液加入12 mL三氯甲烷和12 mL磷酸緩沖液后靜置過夜,在30—32℃水浴中N2吹干后,用SEP柱分離純化,收集過柱的脂肪酸于試管內,在32℃水浴中N2濃縮后加入內標的正己烷溶解,最后用氣相色譜儀測定、結合MIDI軟件系統鑒定分析。微生物脂肪酸的濃度基于單個樣品中的碳內標19:0的濃度計算。用14:0 iso、14:0 anteiso、15:0 iso、15:0 anteiso、16:0 iso、17:0 iso、17:0 anteiso、18:0 iso表征革蘭氏陽性菌;用16:1 ω7c、17:1 ω8c、17:0 cyclo、17:1 ω7c 10-methyl、18:1 ω7c表征革蘭氏陰性菌;革蘭氏陽性菌和革蘭氏陰性菌豐度之和表征細菌;16:1 ω5c、18:3 ω3c、18:2 ω6c、18:1 ω9c表征真菌;16:1 ω5c、18:2 w6c表征叢枝菌根真菌;16:0 10-methyl、17:0 10-methyl、18:0 10-methyl表征放線菌[27]。
1.3.3宏基因組測序分析
使用PowerSoil DNA Isolation Kit(MoBio Laboratories, Carlsbad, CA)試劑盒,提取土壤樣品中的基因組DNA。Qubit對DNA濃度進行精確定量后,采用1%瓊脂糖凝膠電泳,檢測樣品DNA質量。檢測合格的DNA樣品用Covaris S220超聲波破碎儀將基因組 DNA 隨機打斷成300 bp左右的小片段,使用Agencourt AMPure XP試劑盒純化打斷產物。將純化后的打斷產物使用NEBNext Ultra II DNA Library Prep Kit試劑盒進行文庫構建,具體步驟及試劑參考試劑盒說明書。文庫構建完成后,先使用Qubit2.0進行初步定量,稀釋文庫至2 ng/ul。隨后使用Agilent 2100對文庫的插入片段進行檢測,插入片段符合預期后,使用qPCR方法對文庫的有效濃度進行準確定量(文庫有效濃度>3 nM),以保證文庫質量。質檢合格的文庫在Illumina HiSeq4000平臺測序。
1.4 數據分析
采用配對樣本t檢驗來分析增溫對土壤溫度和濕度的影響;采用單因素方差分析法,檢驗溫度升高對干濕季土壤基本理化因子以及微生物生物量的影響。微生物群落結構響應溫度升高的變化用主成分分析(PCA)檢驗。顯著性水平設為P<0.05。運用軟件Excel 2016,SPSS 21.0,Sigmaplot 14.0,R 3.6.3對上述數據進行分析及作圖處理。
宏基因組數據的處理,使用fastp(v0.20.0)軟件對序列(reads) 3′端和5′端的接頭(adapter)序列進行質量剪切,保留高質量的雙端序列(pair-end reads)和單端序列(single-end reads)。所得優化序列用拼接軟件MEGAHIT(1.1.2)進行拼接組裝,過濾結果中300 bp以下的片段。使用MetaGene對拼接結果中的序列重疊群(contigs)進行開放閱讀框(ORF)預測,用CD-HIT(v4.6.1)對所有預測的基因序列進行聚類并構建非冗余基因集。使用SOAPaligner(v2.21)將測序數據與非冗余基因集進行比對,統計基因在對應樣品中的豐度信息。使用Diamond(v0.8.35)將預測得到的非冗余基因集與KEGG數據庫進行比對,采用Pathway對應的碳水化合物代謝(Carbohydrate metabolism)計算土壤有機碳代謝基因豐度;采用EC對應的基因豐度總和計算淀粉酶(Amylase)、纖維素酶(Cellulase)、半纖維素酶(Hemicellulase)、幾丁質酶(Chitinase)的基因豐度。
2 結果
2.1 土壤溫濕度對增溫的響應
增溫處理顯著改變了土壤溫濕度,旱季增溫效果更明顯。圖1可見,2016—2018年間,增溫處理下,土壤月平均溫度在0—10 cm土層增加2.48℃,增溫效果顯著(P<0.01);土壤濕度在0—10 cm土層下降23.93%,增溫處理對土壤濕度影響顯著(P<0.01)。其中,在2017年10月至2018年3月(干季),增溫處理使0—10 cm土層土壤溫度上升2.81℃ (P<0.05),土壤濕度下降29.31% (P<0.01);2018年4—9月(濕季),增溫處理使0—10 cm土層土壤溫度上升2.24℃ (P<0.05),土壤濕度下降27.74% (P<0.01)。采樣當月(12月)增溫處理使0—10 cm土層土壤溫度上升4.12℃ (P<0.01),濕度下降36.15% (P<0.01);6月增溫處理使0—10 cm土層土壤溫度上升1.47℃ (P<0.05),土壤濕度下降19.52% (P<0.05)。
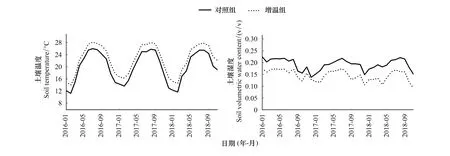
圖1 對照組和增溫組0—10cm的月平均溫度和濕度Fig.1 Monthly mean temperature and moisture of the control and the warming treatment in the dry and wet season (from January 2016 to November 2018)
2.2 土壤理化性質對增溫的響應
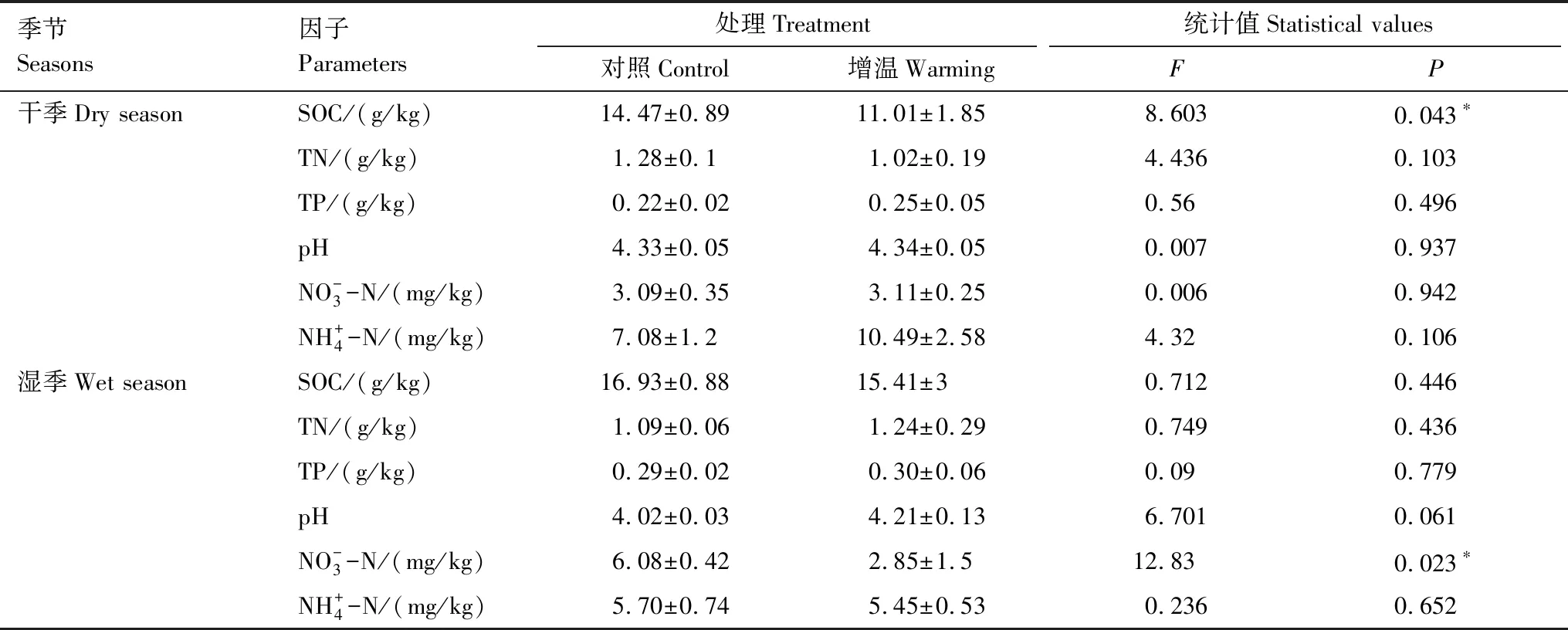
表1 增溫對干濕季土壤理化性質的影響
2.3 不同種群微生物生物量對增溫的響應
圖2可見,增溫對干濕季土壤總PLFAs、Bacteria、Fungi、Act、AMF、G+/G-的影響不顯著。干季總PLFAs、Bacteria、Act、AMF、G+/G-的絕對豐度隨溫度升高略有下降,Fungi的絕對豐度隨溫度增高略有上升,濕季土壤總PLFAs、Bacteria、Fungi、Act、AMF的絕對豐度隨溫度升高而上升,G+/G-隨溫度升高而下降。
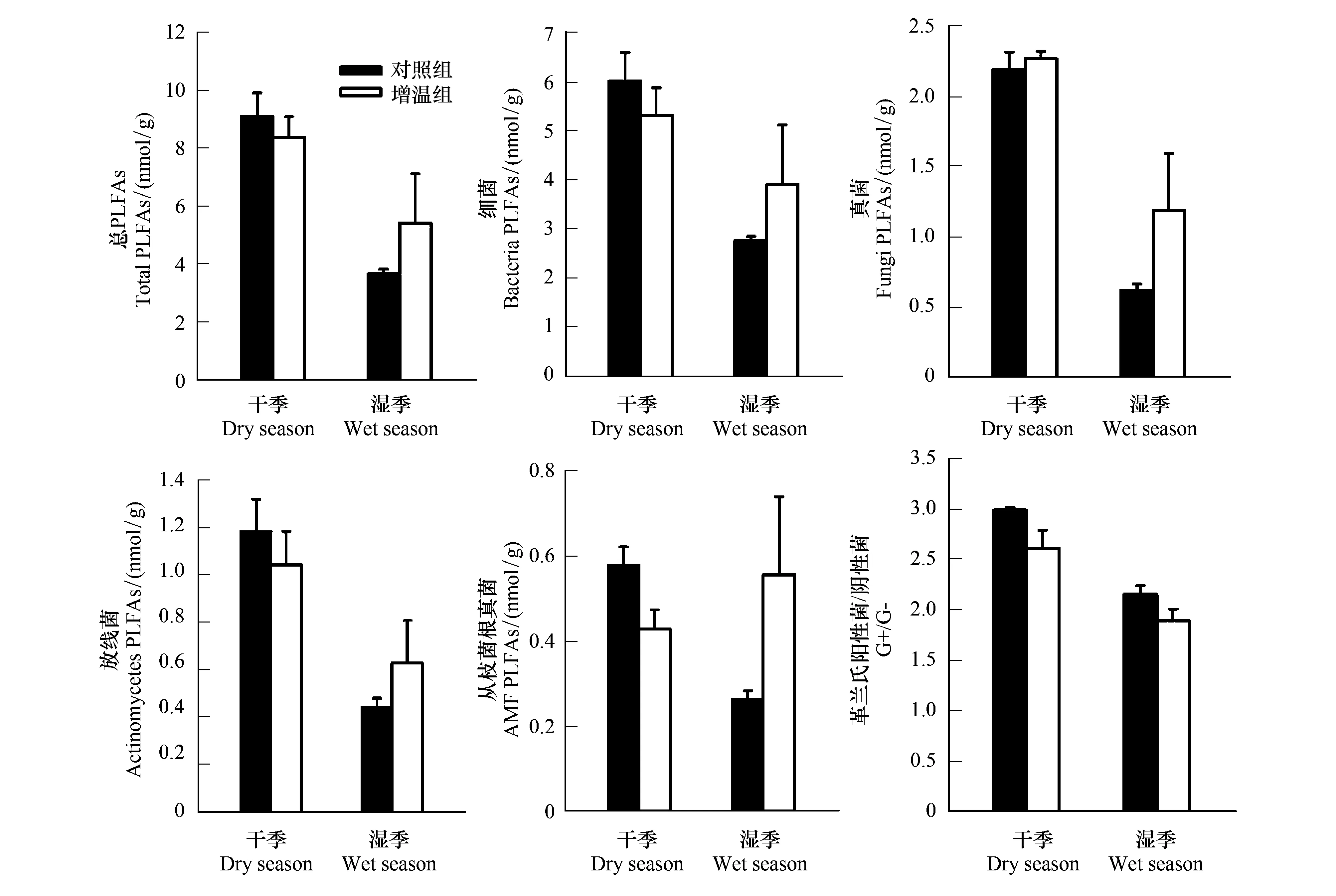
圖2 增溫對干濕季微生物磷脂脂肪酸含量影響Fig.2 Effects of warming on microbial phospholipid fatty acids (PLFAs) content in the dry and wet seasons
如圖3可見,在干季和濕季中,細菌相對豐度降低,真菌相對豐度上升,真菌細菌比值(Fungi/Bacteria)增加,且細菌、真菌、真菌細菌比值在濕季中變化顯著。
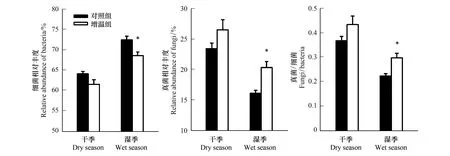
圖3 增溫對干濕季微生物相對豐度影響Fig.3 Effects of warming on microbial relative abundance in the dry and wet seasons
由圖4可見,在干季時,主成分軸分別解釋了群落結構變異的74.2%和18.5%,總解釋度為92.7%,增溫和對照組差異不顯著(P>0.05)。在濕季時,主成分軸分別解釋了群落結構變異的68.8%和27.2%,總解釋度為96%,增溫和對照組差異顯著(P<0.05)。
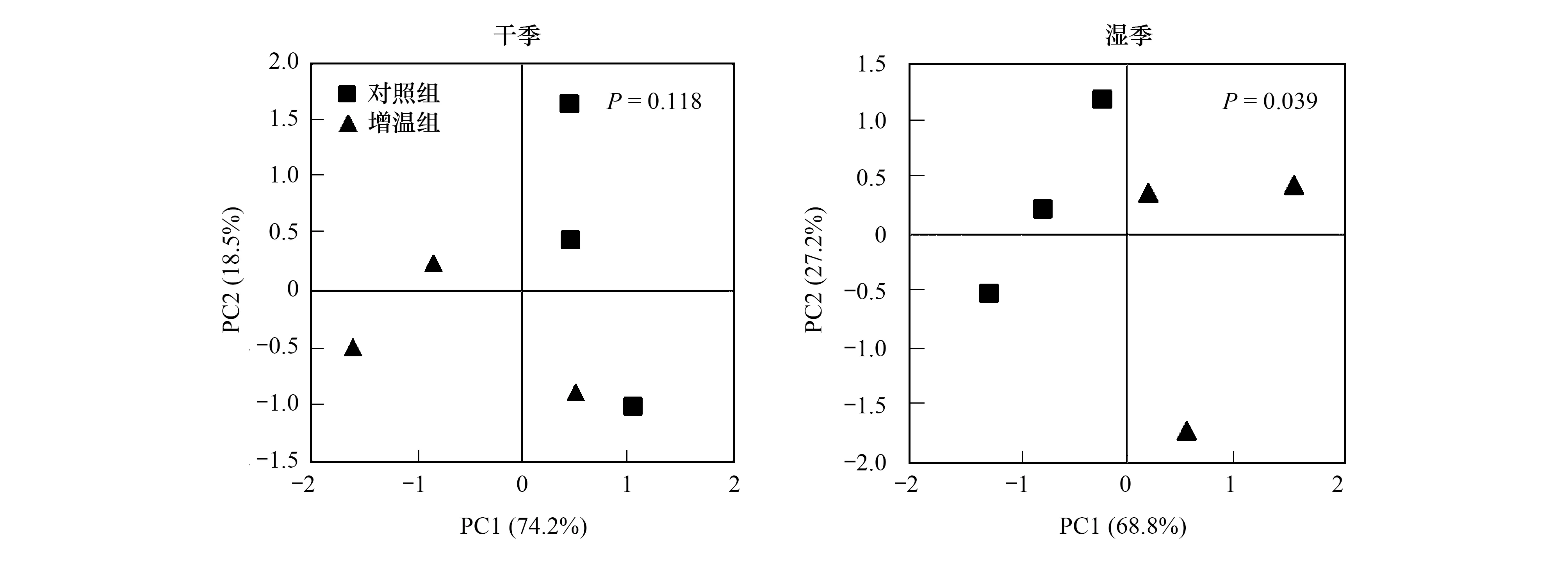
圖4 干濕季土壤微生物群落結構的主成分分析Fig.4 Principal component analysis of soil microbial community structure in the dry and wet seasons
RDA分析結果表明(圖5),土壤濕度為影響干季和濕季土壤微生物群落結構的主要因素。在干季中,第1軸和第2軸分別解釋土壤微生物群落結構變化的91.3%和6.3%,土壤濕度能解釋群落結構變異的度為50.2%(P>0.05);濕季中,第1軸和第2軸分別解釋土壤微生物群落結構變化的92.5%和6.5%,土壤溫度能解釋群落結構變異的度為79.2%(P<0.05)。
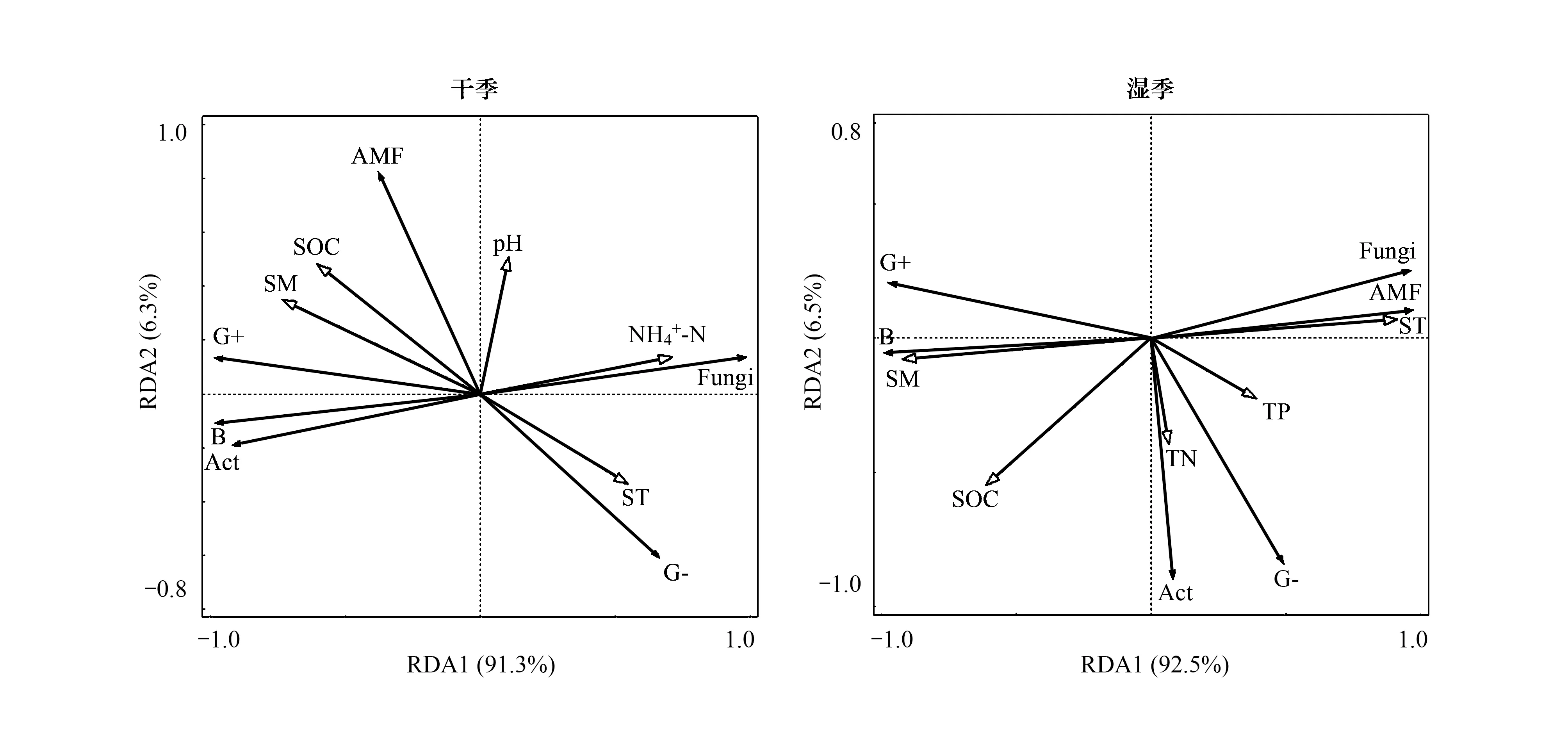
圖5 干濕季微生物PLFAs數據與環境因子RDA分析Fig.5 Redundancy analysis of PLFAs of soil microbial and environmental parametersin the dry and wet seasonsB:細菌 Bacteria;Fungi:真菌;G+:革蘭氏陽性菌 Gram-positive bacterial;G-:革蘭氏陰性菌 Gram-negative bacterial;ACT:放線菌 Actinomycetes;AMF:叢枝菌根真菌 Arbuscular mycorrhizal fungi;ST:土壤溫度 Soil temperature;SM:土壤濕度 Soil moisture;SOC:土壤有機碳 Soil organic carbon;TN:土壤全氮 Total soil nitrogen;TP:土壤全磷 Total soil 銨態氮 Ammonium 硝態氮 Nitrate nitrogen
2.4 增溫對碳代謝功能基因的影響
土壤碳循環功能基因分析結果表明(圖6),在干季和濕季中,增溫處理下淀粉酶、纖維素酶、半纖維素酶、幾丁質酶基因豐度高于對照組。干季中增溫處理使土壤總有機碳代謝基因豐度略有降低,濕季中增溫處理使土壤總有機碳代謝基因豐度上升。在干季中,與對照組相比,增溫處理下淀粉酶、纖維素酶、半纖維素酶、幾丁質酶基因豐度分別上升了3.18%、8.78%、8.99%、12.84%,總有機碳代謝基因豐度下降了0.14%。在濕季中,增溫處理使土壤淀粉酶、纖維素酶、半纖維素酶、幾丁質酶基因豐度增加,與對照組相比,增溫處理下淀粉酶、纖維素酶、半纖維素酶、幾丁質酶等基因豐度分別上升了9.64%、7.04%、3.51%、0.52%,土壤總有機碳代謝基因豐度上升了9.92%。在干季和濕季中,纖維素酶基因豐度占總有機碳代謝基因豐度比例最高,分別為1.35%和1.47%。
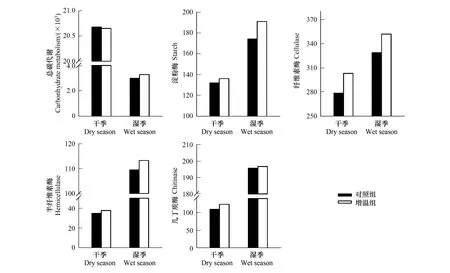
圖6 增溫對干濕季土壤有機碳代謝功能基因絕對豐度影響Fig.6 Effects of warming on absolute abundance soil carbon metabolism function genes in dry and wet seasons縱坐標表示基因注釋到ko (直系同源蛋白)的數目
3 討論
3.1 增溫對土壤不同種群微生物量的影響
本實驗中,干季增溫抑制了細菌的生長,促進了真菌的生長,RDA分析結果顯示這種變化與土壤濕度有關。本實驗干季增溫使土壤含水量顯著下降了24.99%,采樣當月(12月)更是下降了36.15% (圖1),顯然,干季增溫加劇了土壤水分的限制,影響了細菌的生長,Sowerby等研究了橫跨南北歐的四種歐石楠叢生的生態系統土壤微生物和酶活性,結果表明干旱(水分限制)和氣溫上升降低了土壤微生物活性[28]。與細菌相比,真菌對溫度升高引起的水分脅迫有更強的抵抗力[15],而含水量的降低使土壤通透性增加,有利于更依賴氧氣生長的真菌[29],由于它們的絲狀性質,對較高的土壤溫度更有耐受性[30],這在本實驗增溫條件下土壤真菌生物量增加這一結果得到驗證。盡管增溫提高了真菌生物量,但其在土壤微生物占比較低,最終導致干季增溫降低了土壤微生物總PLFAs。
濕季增溫促進了土壤幾乎所有微生物的生長(圖2),微生物群落內真菌/細菌比值顯著增加,RDA分析結果顯示這種變化與土壤溫度和濕度有關。本實驗濕季對照處理OTC樣地土壤(10 cm)平均溫度約23.62℃,平均含水量約20.50% (圖1),是微生物極為適宜的生長環境,增溫促進了微生物的生長,這與許多研究結果相一致[15,31]。濕季增溫雖然使土壤含水量顯著下降了27.74%,但仍保持有15%的含水量(圖1),顯然,在這樣的水分含量條件,溫度增加對土壤微生物的影響遠大于水分含量的下降(圖5)。在本實驗中,無論是在增溫條件下還是對照樣地,土壤微生物生物量均是旱季高于濕季(圖2),這與植物生長對養分的需求密切相關。有研究表明,溫度升高可能通過促進作物生長而加劇根系與微生物之間的養分競爭,利于作物對養分的吸收,導致土壤中微生物生活所需的養分匱乏,進而限制微生物的生長[32]。本實驗地的濕季(4—9月)正是植物生長最旺盛的季節,對養分需求更大,加劇了植物根系生長與微生物對養分需求的競爭。本實驗前期研究也表明,增溫顯著增加了土壤自養呼吸(根呼吸)對土壤總呼吸的貢獻率,而且濕季顯著高于旱季[33],這一結論用本實驗的結果得到了合理的解釋。也有研究結果表明增溫使真菌與細菌PLFAs比值降低[34],或無明顯變化[35],說明土壤微生物對增溫的響應規律存在不確定性,仍有待更多的實驗研究加以揭示。
3.2 增溫對土壤碳代謝基因影響
已有研究表明土壤胞外酶活性可由微生物相關功能基因豐度得到反映[36]。本實驗結果顯示干季和濕季土壤中纖維素酶基因豐度隨溫度升高而上升,這說明增溫提升了土壤纖維素酶活性,這是因為土壤真菌豐度的增加,促進了纖維素酶分泌,加速了土壤難降解有機質的分解。Feng等[37]在美國俄克拉荷馬州中部進行的12年增溫實驗表明,增溫顯著增加了土壤纖維素降解的微生物功能基因的豐度,這與我們研究結果相同。
干季增溫降低了土壤微生物生物量,抑制了土壤微生物生長,土壤有機碳代謝基因豐度下降,而實驗結果表明土壤有機碳含量隨增溫處理顯著降低。這是因為增溫加劇了干季土壤含水量的限制作用(干季土壤濕度顯著下降24.99%),影響了分解過程中的生物化學反應以及土壤微生物群落結構,間接降低土壤微生物的分解作用[38],減少了通過微生物分解作用歸還到土壤的有機碳數量。在濕季的研究中,土壤總有機碳代謝基因豐度隨溫度升高而增加,微生物總PLFAs量也有所上升。這是因為濕季土壤溫度和含水量較高,有利于微生物的生長,增溫進一步促進了微生物的活性。此外,濕季是植物的生長季節,增溫促進植物的生長,植物根系更趨活躍,根系分泌物的增加有利于土壤微生物的繁殖。微生物活性的提高加速了有機質的分解,增加了土壤有機碳的淋溶和歸還[39],進一步豐富了土壤微生物的碳源,使得細菌和真菌豐富度隨溫度升高而增加,土壤有機碳代謝能力增強。MacDonald等[40]研究結果表明,土壤溫度上升會加速土壤微生物呼吸速率和溶解的有機碳淋溶,從而導致土壤中有機碳的損失。Nottingham等[41]通過對秘魯熱帶森林進行5年的增溫實驗發現溫度每升高1℃,土壤有機碳含量下降4%,土壤微生物群落結構發生變化。在土壤含水量較大時,溫度對土壤有機碳分解速率的提升作用較為明顯[42],加劇了土壤有機碳的流失。
4 結論
本研究依托于鼎湖山南亞熱帶山地常綠闊葉林,將模擬的生態系統(含植物和土壤)從高海拔整體移位至低海拔地區形成自然增溫的效果。借助磷脂脂肪酸法(PLFAs)和宏基因組測序等手段,探究土壤微生物對增溫的響應與適應機制。主要結論如下:增溫處理顯著增加了干濕季的土壤溫度,顯著降低了干濕季的土壤濕度,顯著降低了干季土壤有機碳含量與濕季土壤硝態氮含量,對其他土壤理化因子作用不顯著;增溫處理改變干濕季土壤微生物生物量及其群落結構,土壤銨態氮是影響干季土壤微生物群落變化的關鍵因子,溫度是影響濕季土壤微生物群落變化的關鍵因子;增溫抑制了干季山地常綠闊葉林土壤有機碳代謝基因豐度,增強了濕季山地常綠闊葉林土壤有機碳代謝基因豐度。氣候變暖通過改變土壤微生物群落結構及其有機碳代謝功能基因豐度,影響微生物對土壤碳庫的礦化作用,最終影響南亞熱帶山地常綠闊葉林土壤有機碳代謝過程。
