大倉鼠背毛和腹毛紅外光譜差異
2022-02-24 06:09:06楊文建周思宇金志民
野生動物學報 2022年1期
楊文建 周思宇 金志民
(牡丹江師范學院生命科學與技術學院,牡丹江,157011)
大倉鼠(Tscherskiatriton)隸屬于嚙齒目(Rodentia),倉鼠科(Cricetidae),大倉鼠屬,是一種農田害鼠,是多種疾病的傳播媒介和宿主[1]。當前,對大倉鼠的鑒定主要是依據形態學,對鼠類被毛的研究較少[2]。野生動物毛發的主要成分是角蛋白[3],受大倉鼠的進化、分布、行為和環境等的影響,角蛋白的成分會發生一定變化,使用紅外光譜進行鑒定分析具有可行性。傅里葉紅外光譜(FT-IR)是一種發展迅速的檢測技術,能夠以連續波長的紅外光為光源,照射樣品引起分子振動和在轉動能級之間躍遷,進而獲得分子的振轉光譜。通過分析光譜的變化,可以獲得細胞內生物分子結構、構型、構像和數量變化的信息[4]。吳桂芳等[5]對偶蹄目(Artiodactyla)中綿羊的山羊絨和細支綿羊毛進行了紅外光譜分析。也有學者對食肉目(Carnivora)動物中的貉(Nyctereutesprocyonoides)、狼(Canislupus)、豹貓(Prionailurusbengalensis)、金貓(Pardofelistemminckii)、大靈貓(Viverrazibetha)、小靈貓(Viverriculaindica)、豺(Cuonalpinus)、花面貍(Pagumalarvata)、云豹(Neofelisnebulosa)、藏狐(Vulpesferrilata)、水貂(Neovisonvison)、紫貂(Marteszibellina)等進行了被毛紅外光譜分析和對比[6-11]。針對嚙齒目動物中的灰鼠(Sciurusvulgaris)和麝鼠(Ondatrazibethicus)的被毛,也有研究人員進行了紅外光譜分析[12]。目前,國內利用紅外光譜進行嚙齒目動物被毛鑒別的研究還處于探索和積累階段。
嚙齒動物是多種鸮形目(Strigiformes)鳥類喜愛的食物之一[13-14],但利用食團分析法對鸮形目鳥類進行食性分析時,鑒別嚙齒動物種類的主要依據是頭骨和牙齒,而對在食團中占比較大的被毛材料利用較少。被毛這種耐浸泡、耐腐蝕的材料,能夠較好地保存物種的特征性成分[15]。若能夠有效利用食團中殘留的被毛,根據其特征進行種類鑒定,對于鸮形目鳥類的食性分析具有一定意義[16]。本研究對大倉鼠背毛和腹毛的紅外圖譜和二階導數譜進行對比,分析其光譜特征,旨在為鸮形目鳥類食團中殘留的被毛鑒定提供參考,同時豐富嚙齒動物被毛結構的研究內容,為綜合運用被毛特征識別與鑒定動物種類提供科學依據。
1 研究區概況
二道林場位于牡丹江與支流二道河子的交界處,距離柴河鎮約80 km,調查區域周圍海拔為200~400 m。柴河林區(44°47′45″—45°37′30″N,128°59′30″—129°54′30″E)位于黑龍江省東部海林市境內,屬長白山山地張廣才嶺北部,為長白山與小興安嶺過渡山系[17]。
2 材料與方法
2.1 試驗材料
2.2 試驗方法
將拔取的被毛置于無水乙醇中清洗,用超聲波清洗器清洗10 min,倒掉無水乙醇,重復清洗4次,將樣品置于烘箱中烘干8 h后取出備用。
加入液氮將被毛研磨至粉末狀,得到大倉鼠背毛粉末1.436 g,腹毛粉末0.444 g,分別取1~2 mg在瑪瑙研缽中研磨成細粉末,與干燥的溴化鉀粉末(約100 mg)均勻混合。將樣品裝入模具內,在壓片機上壓制成片。然后,放入傅里葉變換紅外光譜儀樣品光路中,掃描波數為400~4 000 cm-1,開啟自動分析系統,得到分析數據。重復壓片分析5次。
2.3 數據處理
使用Excel軟件對試驗數據進行預處理,處理后的數據使用Origin 2018軟件進行繪圖和分析。以掃描波數為橫坐標,透過率為縱坐標繪圖,透過率是紅外光透過樣品的光強和紅外光透過背景(空光路)的光強的比值,即紅外光透過樣品物質的比例,能夠直觀地看出樣品對紅外光的吸收情況。采取定性分析,相應波長的特征峰是對比判斷的主要依據,譜線高低僅能代表所制壓片薄厚不同導致的待測物濃度差異,不能作為成分分析依據,因此對400~4 000 cm-1的紅外光譜圖進行分析,確定其指紋區是在400~1 330 cm-1,指紋區內的紅外吸收光譜復雜,能反映分子結構的細微差異[18]。指紋區中600~900 cm-1變化明顯,故選取這一范圍內的譜圖進行特征識別。
3 結果與分析
3.1 背毛和腹毛紅外光譜差異
由大倉鼠背毛和腹毛樣品的紅外圖譜(圖1)可知,在400~4 000 cm-1內,大倉鼠背毛和腹毛的紅外圖譜有著極高的相似性,均在622、1 562、1 637 cm-1附近有明顯的峰型和基本一致的吸收峰,在3 415、3 470 cm-1均有明顯的吸收峰,形成W形圖譜。背毛在1 439 cm-1、腹毛在1 426 cm-1處各有1個峰型基本一致的吸收峰,但腹毛的峰強比背毛的峰強大。

圖1 大倉鼠背毛和腹毛紅外光譜
3.2 背腹毛紅外譜圖放大差異
為了更好地比較大倉鼠背毛和腹毛的特點,對600~900 cm-1內的紅外光譜進行放大比較(圖2)。由圖2可見,在600~900 cm-1內,大倉鼠背毛和腹毛的紅外圖譜基本一致,均在721、782 cm-1有明顯的峰型一致的吸收峰形成,在779、846 cm-1附近有吸收峰出現。但背毛和腹毛分別在848、845 cm-1有明顯的峰型一致的吸收峰出現,在700~727 cm-1附近背毛和腹毛譜線較為平滑,未形成明顯的吸收峰。
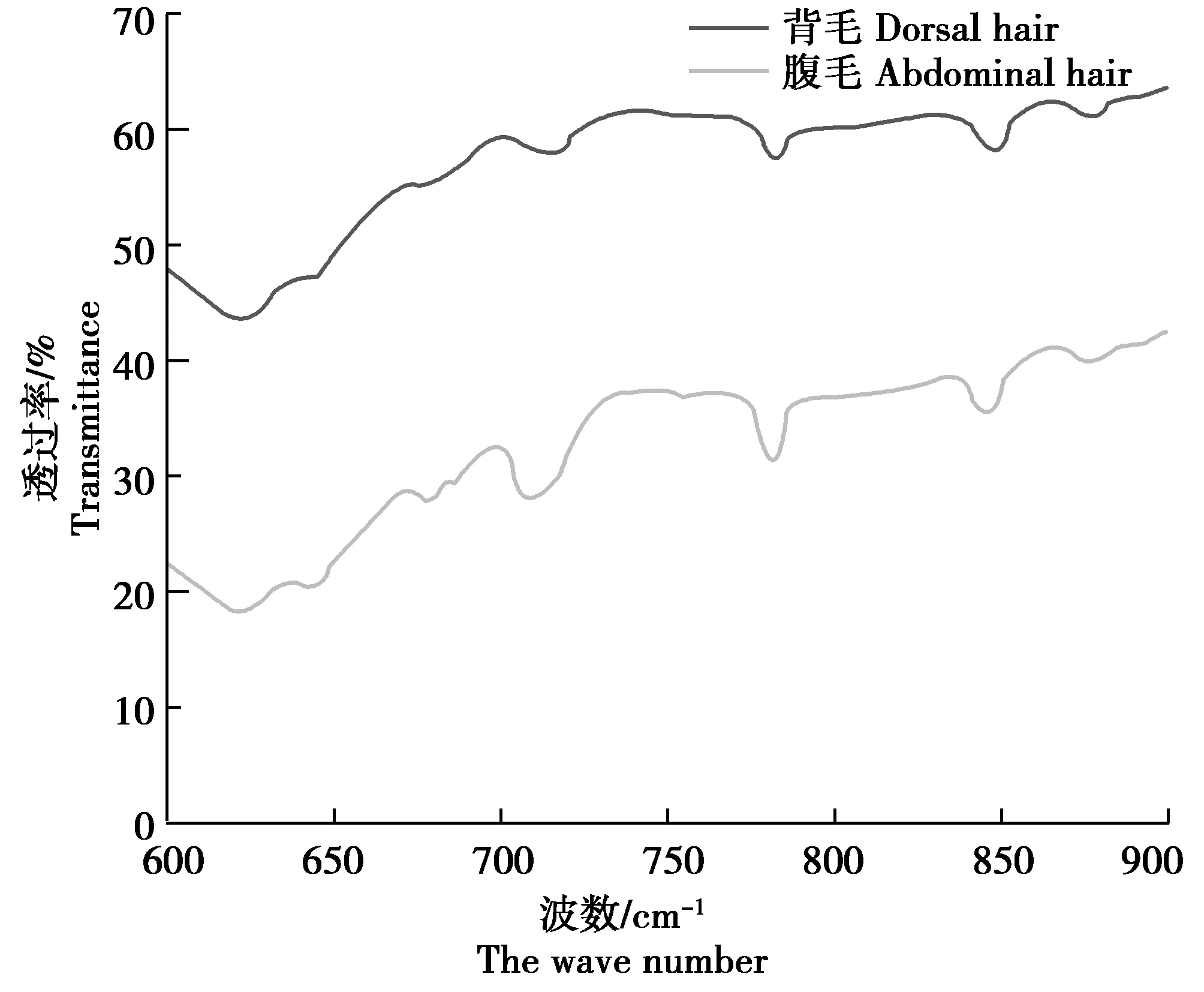
圖2 大倉鼠背毛和腹毛紅外放大光譜
綜上可見,大倉鼠背毛和腹毛的吸收峰基本一致,2個部位的被毛組成物質基本一致,是大倉鼠物種特征的一部分;但是背毛和腹毛之間還存在著一定差異,腹毛的特征性吸收峰、峰強和峰面積均大于背毛,這可能是同一動物背毛和腹毛組成蛋白的含量或者蛋白側鏈的種類不同導致的差異。
3.3 背毛和腹毛的二階導數譜
二階導數可以有效加強紅外光譜的分辨率,為了更好地比較大倉鼠背毛和腹毛之間的差異,分別對2種毛600~900 cm-1的二階導數進行了比較(圖3)。由圖3可見,波數在600~900 cm-1的二階導數圖譜可以反映出更多在紅外圖譜中無法觀察到的特點。腹毛的吸收峰峰強均較大,在786、776 cm-1腹毛的峰強最大,也最容易觀察到;在703、683、854 cm-1,2種毛均有明顯的吸收峰。雖然大倉鼠背毛和腹毛的二階導數圖譜吸收峰的峰型和波數基本一致,但也存在一些差異,在621~645 cm-1腹毛有4個明顯相似的單峰,形成2個W形圖譜,背毛僅在632~645 cm-1有2個明顯相似的單峰,形成1個W形圖譜;背毛和腹毛分別在652~655、650~655 cm-1形成1個單峰;在700 cm-1背毛形成1個單峰,腹毛未形成明顯的峰;在707~718 cm-1,背毛有2個明顯的單峰,形成一個W形圖譜,腹毛有1個明顯的單峰;腹毛在826 cm-1處有1個明顯的吸收峰出現,背毛在822、826、830 cm-1處有吸收峰出現,所以,腹毛形成1個M形圖譜,背毛形成2個M形圖譜。

圖3 大倉鼠背毛和腹毛二階導數譜
在二階導數譜中,大倉鼠背毛和腹毛之間的微小差異更加明顯,一些在紅外光譜中不明顯的差異通過二階導數譜中可以觀察到。二階導數譜進一步證明了大倉鼠背毛和腹毛基本一致,但具有一定差異,是對紅外光譜圖分析結果的有效補充。
4 討論
目前國內外對于動物被毛的識別鑒定主要是在微觀層面上對被毛的鱗片、髓質和橫斷面等進行觀察比較[19]。有學者系統論述了被毛種屬鑒定的原則、方法和注意事項等,并且根據其總結的被毛長度、外觀特征、被毛各部分的鱗片形態、髓質形態、被毛不同部位的橫斷面形態和髓質橫斷面形態和數值指標等,對西歐地區的73種哺乳動物(Mammalia)進行了分類,證實了毛的形態結構在分類應用中的可行性與可靠性[20]。有學者利用掃描電鏡和光學顯微鏡進行觀察分析,發現背部、腹部、臀部和頸部被毛的鱗片類型和排列順序均有一定的差異,例如大靈貓的背毛不具有方瓣型鱗片,而腹毛具有一定量的方瓣型鱗片[21]。侯森林[22]對赤狐(Vulpesvulpes)的背毛和腹毛進行了顯微形態學特征研究,發現赤狐背毛主要鱗片類型為雜波型,腹毛的主要鱗片類型為長瓣型,背毛的髓質指數較腹毛的大。被毛的髓質指數越大,髓質越發達,能夠容納的靜止空氣也越多,使得背毛保溫的能力較腹毛更強。被毛在結構和功能上是相互關聯的,隨著其生理功能的變化,被毛的結構也會有所變化,進而引起形態的變化[23]。有學者對絨山羊四肢不同部位的被毛進行對比分析發現,被毛從四肢近端到遠端,髓質指數呈梯度遞減趨勢,表明山羊四肢離地表越近,體溫越低,與環境的溫差越小,越有利于保持體溫。相應地,體表溫度低則被毛保溫功能相對減弱,髓質發達程度也相對趨于減少[24]。通過已有研究判斷,依據微觀狀態下鱗片的類型、排列順序、髓質指數等,動物被毛可以作為種類鑒別的有效途徑。
本研究表明,通過傅里葉紅外變換光譜和二階導數譜可以有效地顯現大倉鼠被毛的特征,如不同部位的被毛均在622、782、846 cm-1形成明顯的吸收峰,故可以運用到動物種類的鑒定之中,為鸮形目鳥類的食性分析提供參考。而譜圖中出現的差異,如在707~718 cm-1,背毛有2個明顯的單峰,形成1個W形圖譜,腹毛只有1個明顯的單峰,則有效地證明了大倉鼠不同部位的被毛由于受到遺傳進化、生態環境和生活習慣等因素的影響,組分會發生一定的變化,進一步說明了紅外光譜法分析動物被毛的靈敏性和準確性。由于被毛成分的差異,紅外光譜對動物被毛的分析數據具有極高的鑒定性,能夠有效鑒別不同種類的動物[25],因此利用紅外光譜對被毛進行對比分析的方法能夠有效地補充和完善被毛研究的試驗思路。但是,目前利用紅外光譜對動物被毛進行分析鑒定的研究較少,通過紅外光譜對未知動物被毛進行鑒定的參考數據尚不充分,還需進行大量研究來構建動物被毛紅外光譜譜圖庫,為動物被毛的鑒定提供有效參考和依據。
