煙草根莖部病原真菌的分離與鑒定
2022-03-08 03:32:10李義婷蔡劉體張長青尹軍良汪漢成
中國煙草科學 2022年6期
關鍵詞:煙草
劉 茜,李義婷,蔡劉體,張長青,尹軍良*,汪漢成*
煙草根莖部病原真菌的分離與鑒定
劉 茜1,2,李義婷1,蔡劉體2,張長青1,尹軍良1*,汪漢成2*
(1.長江大學農學院,湖北 荊州 434025;2.貴州省煙草科學研究院,貴陽 550081)
為系統了解煙草根莖部主要病原真菌,采集根莖部發黑腐爛病株進行室內分離與鑒定,共分離到病原真菌64株。根據菌落、菌絲和孢子形態特征觀察以及多基因位點系統發育分析對病原菌進行分類鑒定,并在煙草葉、莖和根部進行病原菌致病力測定。結果表明,分離得到的病原菌包括:尖孢鐮刀菌()、接骨木鐮刀菌(.)、共享鐮刀菌(.)、蘇拉威鐮刀菌(.)、柑橘鐮刀菌(.)、番薯鐮刀菌(.)、條紋孢殼屬()、小孢根霉()和枝狀枝孢菌()。致病力測定表明,這些病原真菌對煙草都有致病力,其中接骨木鐮刀菌、尖孢鐮刀菌、共享鐮刀菌和的致病力較強,蘇拉威鐮刀菌、番薯鐮刀菌、柑橘鐮刀菌、小孢根霉和枝狀枝孢菌致病力稍弱。病原真菌的分離與鑒定結果將為科學、合理、有效地防治煙草根莖部病害提供依據。
根莖部病害;鐮刀菌屬;條紋孢殼屬;根霉屬;枝孢屬
煙草是重要的經濟作物,然而,煙草的產量和品質受到病害的嚴重威脅,年損失高達7億元[1-2]。目前,由于煙草品種較為單一,且連年連片種植,導致煙草根莖部病害愈發嚴重,煙葉的產量和品質受到嚴重影響[3]。由鐮刀菌侵染煙草引起的根腐病在美國、韓國、加拿大、澳大利亞、南非等產煙國普遍發生。近兩年鐮刀菌根腐病在我國煙區(云南、河南、貴州、湖北、安徽等地)的發病范圍和發病程度呈上升趨勢,病情嚴重的煙田發病率高達30%以上[4-6],往往成為地區毀滅性的病害[7]。由于對病原菌的認識不清楚,導致用藥缺乏針對性,防效不佳,過度用藥則造成環境污染和病原菌抗藥性增強,給病害防治工作增加了難度。目前,有關根莖部主要病原菌種類鑒定、菌株致病力的差異等均缺乏系統的研究[8]。而明確病原菌的種類及致病力是科學有效防治煙草根莖部病害的基礎。因此,本試驗采集根莖部腐爛發黑的煙草病株,對其病原真菌進行了分離;通過形態特征、分子系統發育分析以及人工接種后的發病癥狀,明確這些病原真菌的種類和致病力,為煙草根莖部病害的防控提供參考。
1 材料與方法
1.1 試驗材料
1.1.1 病樣采集 2020年8月在貴州省正安縣四聯村(28° 57' N,107° 28' E)和鳳岡縣熊坪村(27° 72' N,107° 71' E)、廣西上林縣東禮村(23° 43' N,108° 61' E)采集煙田根莖部發黑腐爛的病株,分別獲得32、28和77個樣品。
1.1.2 煙草材料 品種為云煙87(貴州煙草科學研究院提供),于溫室種植(22 ℃,相對濕度>85%,16 h光照/8 h黑暗)。長至8葉期時,在葉部(從上至下第3~4葉位)、莖部和根部進行致病力測定。
1.2 試驗方法
1.2.1 病原真菌的分離 參照李小杰等[3]的方法,將發病根莖用自來水沖洗干凈,瀝干水后,先后用75%的酒精棉球、無菌水和15%的次氯酸鈉消毒,挑取長出的菌絲尖端轉移至PDA培養基上培養,純化好的單菌落編號保存。
1.2.2 病原真菌的形態學觀察 參照何世芳等[1]的方法,將直徑為6 mm的菌餅轉接至新鮮的PDA培養基(90 mm)中央,25 ℃恒溫黑暗培養。培養7 d后,觀察菌落形態、顏色、質地等,拍照并用十字交叉法測量菌落直徑。挑取培養菌絲和分生孢子制成臨時水玻片,在微分干涉顯微鏡(NiKON DS-Ri2)下觀察菌絲和分生孢子的形態。
1.2.3 病原真菌的系統發育分析 刮取菌絲至1.5 mL離心管中,參照劉海峰[9]的方法,快速提取基因組DNA,并以之為模板進行PCR擴增,獲得ITS(ITS4,ITS5)[10]、(728F,986R;EF1,EF2)[11-12]、(512F,783R)[11]和(5F2,7CR)[13]的目的片段。PCR反應體系和反應程序參考HE等[14]的方法。PCR產物送至擎科基因生物科技有限公司測序。使用Phydit拼接序列,上傳至NCBI(https://www.ncbi.nlm.nih.gov/)進行BLAST比對分析,下載同源性高的菌株信息,利用MEGA7軟件,構建Neighbor-Joining(NJ)系統發育樹(Bootstrap 1000)[15],進一步鑒定分析。
1.2.4 病原真菌的致病力測定 進行煙草葉部、莖部和根部的致病力試驗,葉部和莖部接種參考韓鳳等[16]方法,根部接種參考邱睿等[5]方法,接種后放在溫室中培養,7 d后統計病斑大小。重復3次,每次重復2株。對發病煙苗采用組織分離法重新分離,鏡檢觀察分離物,完成柯赫氏法則驗證。對照試驗接種無菌的PDA培養基塊。
2 結 果
2.1 病原真菌分離
根莖部病樣進行病原真菌分離,共獲得64株菌株。根據菌株菌落、菌絲及分生孢子的形態特征,初步將64株菌株鑒定為鐮刀菌屬(spp.)57株,條紋孢殼屬(spp.)2株,根霉屬(spp.)3株和枝孢屬(spp.)2株[17-20]。
2.2 鐮刀菌屬病原菌分類鑒定
根據鐮刀菌屬病原菌菌株的菌落、菌絲、分生孢子形態(圖1),初步將57個鐮刀菌菌株分為6組。利用ITS和引物進行PCR擴增后,獲得鐮刀菌屬代表菌株的基因片段,有效長度分別為453~533 bp、656~704 bp。得到的序列在NCBI上比對,結果表明有19株接骨木鐮刀菌、1株番薯鐮刀菌、6株蘇拉威鐮刀菌、6株柑橘鐮刀菌、17株尖孢鐮刀菌和8株共享鐮刀菌。每個菌種隨機選取一株,進行系統的形態特征描述、系統發育分析和致病力測定。
菌株GZAX 501(接骨木鐮刀菌)7 d菌落直徑可達90 mm,菌落圓形規則,白色氣生菌絲呈絮狀,瓊脂中的色素為黃色(圖1A1);菌絲分枝少,有隔膜(圖1B1)。CLA培養基上大型分生孢子呈無色鐮刀形,大小均勻,頂端細胞較尖,足形基底細胞,3~5隔,多3隔,大小為(20.0~58.5)μm × (3.5~12) μm(圖1C1)。
菌株GZAX 207(番薯鐮刀菌)7 d菌落直徑62 mm,白色菌落近圓形,邊緣波紋狀或整齊,氣生菌絲絨狀至棉絮狀(圖1A2);菌絲分枝少,有隔膜(圖1B2)。大型分生孢子細鐮刀狀,背腹彎曲,4~6隔,多5隔,有明顯的足狀基細胞和逐漸變細的頂端細胞,(32.0~67.0) μm × (3.5~8.5) μm(圖1C2)。
菌株GZAX 211(蘇拉威鐮刀菌)7 d菌落直徑85 mm,菌落圓形規則,菌絲放射性生長,中間菌絲呈棉絮狀(圖1A3);菌絲分枝多,有隔膜(圖1B3)。大型分生孢子呈紡錘形,2~6隔,多5隔,(23.0~93.0) μm × (4.0~11.5) μm(圖1C3)。
菌株GZAX 408(柑橘鐮刀菌)7 d菌落直徑58 mm,白色菌落圓形規則,菌絲呈棉絮狀至絨狀,(圖1A4);菌絲細,有分枝和隔膜(圖1B4)。大型分生孢子呈鐮刀或紡錘形,2~7隔,多5隔;(23.5~93.0) μm × (4.5~11.5) μm。小型分生孢子橢圓至卵圓形,量極少(圖1C4)。

注:A1~A6,菌株GZAX 501、GZAX 207、GZAX 211、GZAX 408、GZAX 106、GZAX 212在PDA培養基上的菌落特征(左正面,右反面);B1~B6,PDA培養基上的菌絲形態;C1~C6,CLA培養基上的孢子形態;相應菌株在煙草葉部(D1~D6、E1~E6)、莖部(F1~F6)、根部(G1~G6)上的致病力。
菌株GZAX 106(尖孢鐮刀菌)7 d菌落直徑87 mm,菌落呈圓形,白色菌絲體較致密呈絮狀,在瓊脂中產生深品紅色色素(圖1A5);菌絲分枝多,有隔膜(圖1B5)。大型分生孢子幾乎筆直,以3隔為主,(30.5~68.5) μm × (4.5~10.0) μm。
菌株GZAX 212(共享鐮刀菌)7 d菌落直徑78 mm,菌落呈近圓形,邊緣波紋狀,菌絲茂盛呈絮狀至絨狀,2~3 d時菌落在PDA上呈粉色(圖1A6);菌絲分枝多,有隔膜(圖1B6)。大型分生孢子呈細鐮刀狀或紡錘形,多3隔,有明顯的足狀基細胞和逐漸變細的頂端細胞,(27.0~95.5) μm × (4.5~9.0) μm;小型分生孢子橢圓至卵圓形,無隔膜,(4.5~11.5) μm × (1.5~6.5) μm(圖1C6)。
根據ITS、聯合構建的系統發育樹結果表明(圖2),菌株GZAX 501、GZAX 207、GZAX 211、GZAX 408、GZAX 106、GZAX 212分別與.、.、.、.、.、.聚于同一最小分支。
2.3 條紋孢殼屬病原菌分類鑒定
擴增條紋孢殼屬病原菌的片段,獲得序列有效長度為545~551 bp的序列,在NCBI上進行同源性比對,結果表明分離得到的條紋孢殼屬病原菌為(同源性為100%)。選取菌株GZAX 110進行系統的形態特征描述、系統發育分析和致病力測定。
菌株GZAX 110 生長7 d菌落直徑90 mm,菌絲茂盛呈棉絮狀,菌絲體為灰色,后加深至灰黑色(圖3B);菌絲分枝少,有隔膜(圖3C)。分生孢子為卵圓形或橢球形,部分兩端圓鈍、中間略向內凹陷,部分一端逐漸變細、另一端圓鈍,(21.0~56.5)μm × (13.0~30.0) μm(圖3D)。根據基因位點構建的系統發育樹結果和形態學特征分析表明,菌株GZAX 110與位于同一最小分支(圖3A-D)。
2.4 根霉屬病原菌分類鑒定
擴增根霉屬病原菌的ITS片段,獲得序列有效長度為630~643 bp,NCBI同源性比對(同源性為100%)結果表明根霉屬病原菌為小孢根霉(.)。選取菌株GZAX 112進行系統的形態特征描述、系統發育分析和致病力測定。
菌株GZAX 112的氣生菌絲為白色蓬松(圖4B);無隔菌絲,具有分枝(圖4C)。分生孢子近球形,(5.5~11.0) μm × (3.5~8.0) μm(圖4D)。系統發育分析和形態學特征結果表明,菌株GZAX 112為小孢根霉(圖4A-D)。
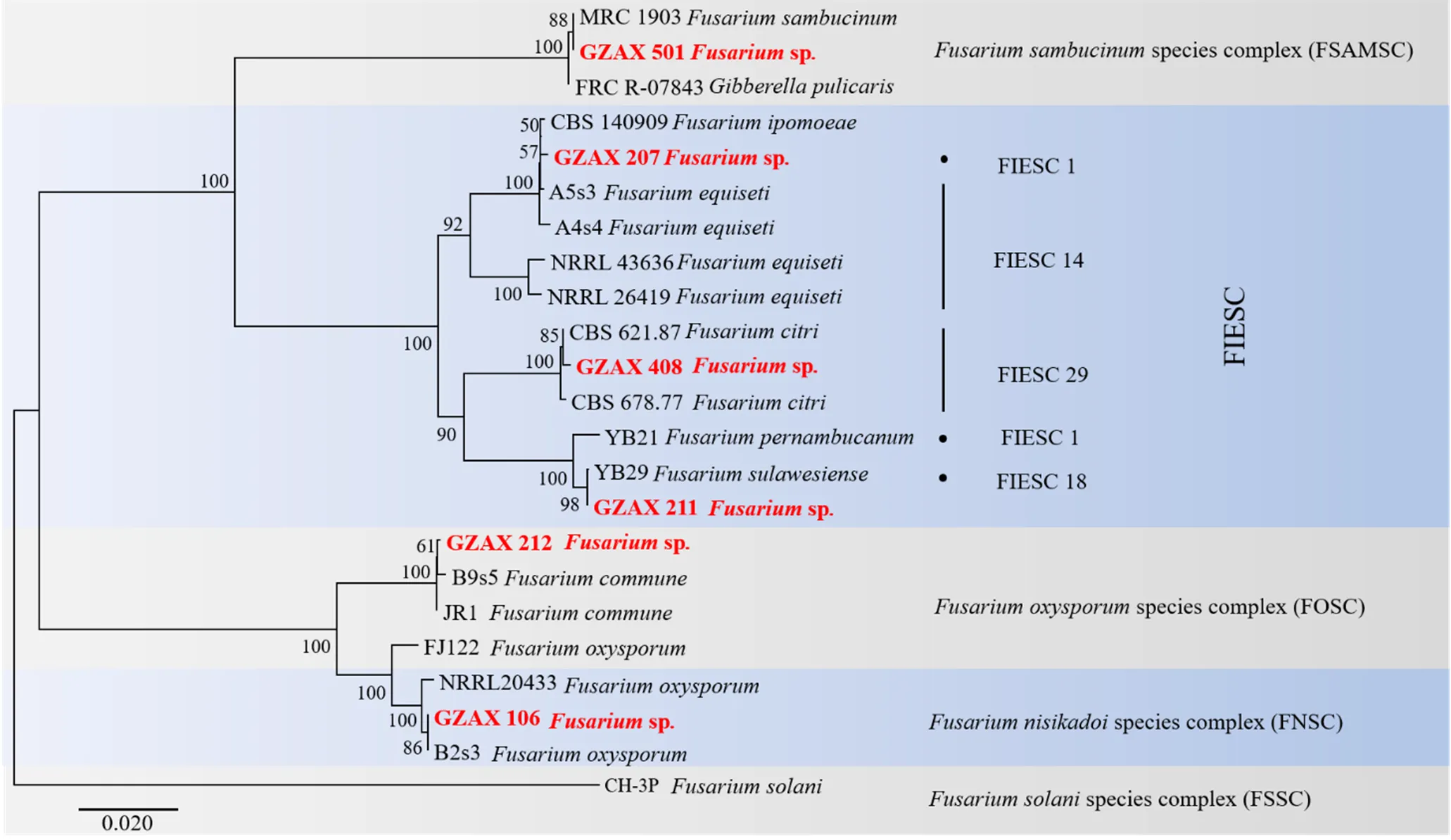
圖2 基于鐮刀菌屬病原菌EF1-α、RPB2序列構建的系統發育樹

注:A,基于條紋孢殼屬病原菌EF1-α序列構建的系統發育樹;B,PDA培養基上培養7 天的菌落特征(左正面,右反面);C,PDA培養基上的菌絲形態;D,PCA培養基上的孢子形態;菌株GZAX 110在煙草根部(E)、葉部(F-G)、莖部(H)上的致病力。

注:A,基于根霉屬病原菌ITS序列構建的系統發育樹;B,PDA培養基上培養7 天的菌落特征(左正面,右反面);C,PDA培養基上的菌絲形態;D,PCA培養基上的孢子形態;菌株GZAX 112在煙草根部(E)、葉部(F-G)、莖部(H)上的致病力。
2.5 枝孢屬病原菌分類鑒定
擴增枝孢屬病原菌的ITS和片段,得到的序列有效長度分別為468~475 bp、228~231 bp,NCBI同源性比對(同源性分別為100%,98.70%)結果表明枝孢屬病原菌為枝狀枝孢菌(.)。選取菌株GZAX 113進行系統的形態特征觀察、系統發育分析和致病力測定。
菌株GZAX 113生長7 d菌落直徑37 mm,菌落形態圓形,邊緣整齊,菌絲體匍匐生長,顏色為草綠色,邊緣為黃色(圖5B);菌絲具有隔膜和分枝(圖5C)。分生孢子多橢球形,兩端略尖,少數腎形,(5.5~26.0) μm × (3.5~7.5) μm(圖5D)。ITS、聯合構建的系統發育樹和形態學特征結果表明,菌株GZAX 113為枝狀枝孢菌(圖5A-D)。

注:A,基于枝孢屬病原菌ITS、ACT序列構建的系統發育樹;B,PDA培養基上培養7天的菌落特征(左反面,右正面);C,PDA培養基上的菌絲形態;D,PCA培養基上的孢子形態;E-H,菌株GZAX 113在煙草根部(E)、葉部(F-G)、莖部(H)上的致病力。
2.6 病原真菌的分離情況與致病力
分離得到的病原菌中接骨木鐮刀菌有19株,占比29.7%;番薯鐮刀菌有1株,占比1.56%;蘇拉威鐮刀菌6株,占比9.4%;柑橘鐮刀菌6株,占比9.4%;尖孢鐮刀菌17株,占比26.6%;共享鐮刀菌8株,占比12.5%;條紋孢殼屬2株,占比3.1%;小孢根霉3株,占比4.7%;枝狀枝孢菌2株,占比3.1%。四聯村的致病病原菌是接骨木鐮刀菌(占比15.6%)、蘇拉威鐮刀菌(18.8%)、柑橘鐮刀菌(18.8%)、尖孢鐮刀菌(31.3%)、小孢根霉(9.4%)和枝狀枝孢菌(6.25%);熊坪村的是接骨木鐮刀菌(25%)、番薯鐮刀菌(6.25%)、尖孢鐮刀菌(31.25%)、共享鐮刀菌(25%)和條紋孢殼屬(12.5%),東禮村的是尖孢鐮刀菌(33.3%)和共享鐮刀菌(66.7%)。
菌株對葉部的致病力試驗表明,菌株GZAX 501(接骨木鐮刀菌)、GZAX 106(尖孢鐮刀菌)、GZAX 212(共享鐮刀菌)、GZAX 110(條紋孢殼屬)致病力較強(圖1D1、D5、D6,圖3F),最強的是GZAX 106(尖孢鐮刀菌),菌株GZAX 110(條紋孢殼屬)不僅可在葉部形成灰黑色病斑,病斑周圍形成黃褐色的暈圈,而且葉部黃化嚴重。菌株GZAX 207(番薯鐮刀菌)、菌株GZAX 211(蘇拉威鐮刀菌)、GZAX 408(柑橘鐮刀菌)、GZAX 112(小孢根霉)、GZAX 113(枝狀枝孢菌)致病力較弱(圖1D2、D3、D4,圖4F,圖5F),不會形成深褐色病斑,但會致使葉部黃化萎蔫。對照未發病。
菌株對莖部的致病力試驗表明,接骨木鐮刀菌、條紋孢殼屬致病力最強(圖1F1,圖3H),莖部在接種菌塊后會整株死亡。番薯鐮刀菌、蘇拉威鐮刀菌、柑橘鐮刀菌、枝狀枝孢菌的致病力較弱(圖1F2、F3、F4,圖5H),只產生較小的病斑。菌株尖孢鐮刀菌、柑橘鐮刀菌、小孢根霉致病力最弱,接種后對莖部幾乎不產生影響(圖1F5、F4,圖4H)。對照未發病。
菌株對根部致病力試驗表明,蘇拉威鐮刀菌、柑橘鐮刀菌、尖孢鐮刀菌、條紋孢殼屬致病力較強(圖1 G3、G4、G5,圖3E),接骨木鐮刀菌、枝狀枝孢菌的致病力較弱(圖1G1、圖5E),番薯鐮刀菌、共享鐮刀菌、小孢根霉的致病力最弱(圖1G2、G6,圖4E),接種病原菌后病株葉部發現不同程度的萎蔫黃化,須根減少。對照未發病。
綜上,統計了各菌株對葉部、莖部和根部的致病力(表1),發現菌株對葉部和莖部的致病力相似,但共享鐮刀菌對葉部致病力強,而對莖部致病力較弱(圖2D6、E6、F6)。煙草葉部和莖部在劃傷和未劃傷情況下致病力略有差異,劃傷致病力普遍強一些。

表1 病原菌的致病力分析
注:“+++”表示致病力強,病斑面積大于30 mm;“++”表示致病力較強,病斑面積10~30 mm;“+”表示致病力弱,病斑面積小于10 mm。
Note: “+++” indicates strong pathogenicity and the diameter of lesion is greater than 30 mm; “++” indicates that moderate pathogenicity and the diameter is 10-30 mm; "+" indicates weak pathogenicity and the diameter is less than 10 mm.
3 討 論
鐮刀菌屬(sp.)真菌是引起植物根腐病的主要致病菌,其種類繁多,地理分布廣[21]。其中尖孢鐮刀菌(.)是引發煙草鐮刀菌根腐病的優勢病原菌群,會導致葉部發黃萎蔫下垂、枯死[2,22]。陳高航[22]發現接骨木鐮刀菌(.)是煙草根腐病的非致病菌。吳安忠等[23]報道共享鐮刀菌(.)為煙草根腐病的主要病原物。本研究結果表明尖孢鐮刀菌和共享鐮刀菌對葉部致病力較強,這與前人研究結果一致;但本研究分離得到的接骨木鐮刀菌對葉部及莖部致病力都較強,與陳高航[22]的研究結果不一致,說明該菌可能存在致病力分化,需要進一步研究。在前人研究中并未出現番薯鐮刀菌(.)、蘇拉威鐮刀菌(.)和柑橘鐮刀菌(.),本研究發現番薯鐮刀菌、蘇拉威鐮刀菌和柑橘鐮刀菌致病力較輕,可見其不是主要病原菌。本研究分離得到的優勢真菌屬為鐮刀菌屬,而鐮刀菌屬(sp.)真菌是引起植物根腐病的主要致病菌[23],因此在煙草生產中需把根腐鐮刀菌作為防治對象。
李小林[24]發現根霉屬是成熟期煙草根際優勢真菌之一;劉宏玉等[25]在分離煙草根、莖和葉內生真菌種類組成時發現,枝孢屬在根莖葉均有分布;但目前并不清楚根霉屬與枝孢屬在煙草上的致病力強弱。條紋孢殼屬并未在以往的研究結果中出現。本研究發現條紋孢殼屬對煙草致病力強,在致病力檢測時,其他菌株均未導致煙草葉部黃化,但.不僅使葉部產生褐色病斑,并且使整個葉部黃化。本研究還發現菌株的種類和采樣地點也有一定關系,尖孢鐮刀菌在四聯村、熊坪村和東禮村的田間發病植株中分離得到,故該菌株不存在地理隔離,在三地普遍引發病害;番薯鐮刀菌和條紋孢殼屬僅在熊坪村的病株中分離得到,蘇拉威鐮刀菌、小孢根霉和枝狀枝孢菌僅在四聯村分離得到,這5株病原菌有地理特異性,推測可能存在地理隔離。有研究表明鐮刀真菌在復合侵染中具有重要的地位與優勢,2種以上病原菌混合接種植株致病力效果比單獨一種侵染發病率高、致病力強,潛育期短[26]。在本研究中,接骨木鐮刀菌,尖孢鐮刀菌,共享鐮刀菌和條紋孢殼屬是煙草根腐病的主要病原真菌。但環境條件導致地域間病害種類及其發生情況不同,因此,為了更符合煙草生產的實際情況,本研究分離得到的多種菌株復合侵染對病原真菌致病力的強弱、侵染速度的快慢及不同條件對不同菌種侵染情況的影響等問題還需進一步研究。
4 結 論
形態特征、分子鑒定和致病力分析表明,發病植株根莖部分離得到的64株病原菌,包括鐮刀菌屬6種,條紋孢殼屬、根霉屬和枝孢屬各1種,鐮刀菌屬是優勢菌群。接骨木鐮刀菌和條紋孢殼屬病原致病力較強。首次從煙草上分離到番薯鐮刀菌、蘇拉威鐮刀菌和柑橘鐮刀菌,致病力較輕,為次要病原菌。在防治根莖部病害時把鐮刀菌屬作為重點防控對象。同時也需監測條紋孢殼屬、根霉屬和枝孢屬的發生與流行情況,對有效控制煙區病害的發生具有重要作用。
[1] 何世芳,桑維鈞,曹毅,等. 煙草彎孢菌葉斑病病原菌鑒定及生物學特性研究[J]. 中國煙草科學,2021,42(5):50-56.
HE S F, SANG W J, CAO Y, et al. Identification and biological characterization ofcausing the tobacco leaf spot disease [J]. Chinese Tobacco Science, 2021, 42(5): 50-56.
[2] 陳志敏. 福建省煙草根莖病害診斷及防治藥劑篩選[D]. 福州:福建農林大學,2009.
CHEN Z M. Diagnosis and screening chemicals to control of tobacco root and stem diseases in Fujian province[D]. Fuzhou: Fujian Agriculture and Forestry University, 2009.
[3] 李小杰,李琦,劉暢,等. 河南煙區煙草根莖類病害調查及病原鑒定[J]. 煙草科技,2022,55(1):41-47.
LI X J, LI Q, LIU C, et al. Pathogen identification of tobacco rhizomatic diseases in tobacco growing areas of Henan province[J]. Tobacco Science & Technology, 2022, 55(1): 41-47.
[4] 邱睿,白靜科,李成軍,等. 河南煙草鐮刀菌的分子鑒定及致病性分析[J]. 中國煙草學報,2018,24(2):129 -134.
QIU R, BAI J K, LI C J, et al. Molecular identification and pathogenicity analysis of tobaccospp. in Henan[J]. Acta Tabacaria Sinica, 2018, 24(2): 129-134.
[5] 陳瑞泰,朱賢朝,王智發,等. 全國16個主產煙省(區)煙草侵染性病害調研報告[J]. 中國煙草科學,1997,1(14):1-7.
CHEN R T, ZHU X Z, WANG Z F, et al. A report of investigating and studying tobacco infectious diseases of 16 main tobacco producing provinces (regions) in China[J]. Chinese tobacco science, 1997, 1(14): 1-7.
[6] 田艷艷,王偉杰,苗圃,等. 河南煙草鐮刀菌的初步分子鑒定[J]. 煙草科技,2014,47(11):89-92.
TIAN Y Y, WANG W J, MIAO P, et al. Preliminary molecular identification ofinfecting tobacco in Henan[J]. Tobacco Sciense & Technology, 2014, 47(11): 89-92.
[7] 魏國勝,周恒,朱杰,等. 土壤PH值對煙草根莖部病害的影響[J]. 江蘇農業科學,2011,39(1):140-143.
WEI G S, ZHOU H, ZHU J, et al. Effects of soil PH on tobacco root and stem diseases[J]. Jiangsu Agricultural Sciences, 2011, 39(1): 140-143.
[8] 苗圃. 河南省煙草真菌性根莖病害鑒定及黑脛病菌生理小種鑒定[D]. 洛陽:河南科技大學,2013.
MIAO P. The Identification of the pathogen of tobacco root and stem fungal diseases and the physiological race ofVar. nicotianae in Henan[D]. Luoyang: Henan University of Science and Technology, 2013.
[9] 劉海峰. 茄科植物鏈格孢菌大孢子種分類研究及菌株YZU 161111全基因組測序[D]. 荊州:長江大學,2020.
LIU H F. Taxonomy of large-sporedfrom Solanaceae and whole genome sequencing of strain YZU 161111[D]. Jingzhou: Yangtze University, 2020.
[10] WHITE T, BRUNS T, LEE S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[J]. PCR protocols: a guide to methods and applications, 1990, 18(1): 315-322.
[11] CARBONE I, KOHN L M. A method for designing primer sets for speciation studies in filamentous ascomycetes[J]. Mycologia, 1999, 91(3): 553-556.
[12] O’DONNELL K, KISTLER H C, CIGELNIK E, et al. Multiple evolutionary origins of 423 the fungus causing Panama disease of banana: Concordant evidence from nuclear and 424 mitochondrial gene genealogies[J]. Proceedings of the National Academy of Sciences, 1998, 95(5): 2044-2049.
[13] REEB V, LUTZONI F, ROUX C. Contribution ofto multilocus phylogenetic studies of the(, Fungi) with special emphasis on the lichen-formingand evolution of polyspory[J]. Molecular phylogenetics and Evolution 2004, 32(3): 1036-1060.
[14] HE L, CHENG H, HTUN A A, et al. Phylogeny and taxonomy of two new(:) species in section Gypsophilae from China[J]. Mycological Progress, 2021, 20(4): 355-363.
[15] CHENG H, ZHAO L, WEI X, et al.species causing leaf spot on hemp () in Northern China[J]. European Journal of Plant Pathology, 2022, 162(4): 957-970.
[16] 韓鳳,李巧玲,韓如剛,等. 渝產多花黃精根腐病病原菌的分離與鑒定[J]. 分子植物育種,2019,18(11):3693-3698.
HAN F, LI Q L, HAN R G, et al. Isolation and identification of pathogen of root rot disease of polygonum produced in Chongqing[J]. Molecular Plant Breeding, 2019, 18(11): 3693-3698.
[17] XIA J W, SANDOVAL-DENIS M, CROUS P W, et al. Numbers to names-restyling the-species complex[J]. Persoonia-Molecular Phylogeny and Evolution of Fungi, 2019, 43(1): 186-221.
[18] NETTO M S B, ASSUN??O I P, LIMA G S A, et al. Species ofassociated with papaya stem-end rot in Brazil[J]. Fungal Diversity, 2014, 67(1): 127-141.
[19] BENSCH K, GROENEWALD J Z, BRAUN U, et al. Common but different: the expanding realm of[J]. Studies in Mycology, 2015, 82(1): 23-74.
[20] 盧燦華,蘇家恩,蓋曉彤,等. 烤煙霉變病原菌鑒定及其生防菌篩選[J]. 中國煙草科學,2022,43(2):45-51.
LU C H, SU J E, GAI X T, et al. Causal pathogen identification and antagonistic bacteria screening of tobacco leaf mold[J]. Chinese Tobacco Science, 2022, 43(2): 45-51.
[21] 高芬,任小霞,王夢亮,等. 中草藥根腐病及其微生物防治研究進展[J]. 中國中藥雜志,2015,40(21):4122-4126.
GAO F, REN X X, WANG M L, et al. Research progress in root rot diseases of Chinese herbal medicine and control strategy by antagonistic microorganisms[J]. China journal of Chinese materia medica, 2015, 40(21): 4122-4126.
[22] 陳高航. 煙草根腐病病原鑒定及其生物學特性觀察[D]. 武漢:華中農業大學, 2013.
CHEN G H. The Identification of tobacco root rot pathogen and its biological characteristics[D]. Wuhan: College of Plant Science and Technology of Huazhong Agricultural University, 2013.
[23] 吳安忠,程崖芝,巫升鑫,等. 煙草鐮刀菌根腐病的病原鑒定[J]. 中國煙草學報,2018,24(2):135-140.
WU A Z, CHENG Y Z, WU S X, et al. Identification of tobaccoroot rot pathogen[J]. Acta Tabacaria Sinica, 2018, 24(2): 135-140.
[24] 李小林. 成熟期煙草根際效應研究[D]. 雅安:四川農業大學,2013.
LI X L. Theeffect of mature stage tocacco ()[D]. Ya’an: Sichuan Agricultural University, 2013.
[25] 劉宏玉. 煙草內生真菌多樣性及促生和抗重金屬菌株的篩選[D]. 杭州:浙江大學,2014.
LIU H Y. Diversity of endophytic fungi in tobacco ang screening on strains with growth-promoting and heavy metal resistant[D]. Hangzhou: Zhejiang University, 2014.
[26] 沈清清,劉芳,胡彥. 藥用植物根腐病病原菌研究進展[J]. 北方園藝,2014,38(11):187-190.
SHEN Q Q, LIU F, HU Y. Research progress of pathogens of root rot disease on medicinal plants[J]. Northern Horticulture, 2014, 38(11): 187-190.
Isolation and Identification of Pathogenic Fungi from the Rhizomes of Tobacco
LIU Xi1,2, LI Yiting1, CAI Liuti2, ZHANG Changqing1, YIN Junliang1*, WANG Hancheng2*
(1. College of Agriculture, Yangtze University, Jingzhou 434025, Hubei, China; 2. Guizhou Academy of Tobacco Science, Guiyang 550081, China)
In order to systematically understand the main pathogenic fungi in the rhizosphere of tobacco, diseased rhizosphere samples were collected from three tobacco planting areas. Then pathogen isolation and identification were performed in the laboratory, and 64 strains of pathogenic fungi were obtained. According to the morphological characteristics of colonies, mycelium, and spores, as well as the phylogenetic analysis of multiple gene loci, the strains were identified at the species level; and their pathogenicity were tested in leaf, stem, and root of tobacco. The results showed that the isolated pathogens included,.,....,,, and. Pathogenicity assays indicated that these pathogens had certain pathogenicity to tobacco. The pathogenicity of,andwere strong. The pathogenicity of.,..,.and.were weak. The classification and identification results of the pathogenic fungi will provide a foundation for the scientific, reasonable, and effective control of tobacco rhizosphere diseases.
root and stem diseases;sp.;sp.;sp.;sp.
10.13496/j.issn.1007-5119.2022.06.007
S435.72
A
1007-5119(2022)06-0045-08
中國煙草總公司貴州省公司科技項目(201914、2020XM03);中國煙草總公司科技項目[110202101048(LS-08)、110202001035(LS-04)];貴州省“百層次”創新型人才項目{黔科合平臺人才-GCC[2022]028-1}
劉 茜(1998-),女,主要從事煙草病原真菌學研究。E- mail:1419650896@qq.com
,E-mail:尹軍良,yinjunliang@yangtzeu.edn.cn;汪漢成,xiaobaiyang126@hotmail.com
2022-03-29
2022-10-05
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
