大紅浙醋醋酸菌分離及其產(chǎn)酸關(guān)鍵氨基酸分析
2022-03-10 12:09:18石暉琴張彥民遲煥榮李兆飛趙國忠
中國釀造 2022年2期
石暉琴,張 工,張彥民,遲煥榮,李 沛,李兆飛,趙國忠*
(1.天津科技大學 食品科學與工程學院 省部共建食品營養(yǎng)與安全國家重點實驗室,天津 300457;2.山東巧媳婦食品集團有限公司,山東 淄博 255400;3.安琪酵母股份有限公司,湖北 宜昌 443003)
食醋在我國有著3 000多年的發(fā)展歷史,其作為調(diào)味品可以提升食物的風味和口感。大紅浙醋是一種屬于廣東特色的中國傳統(tǒng)食醋,由于復雜的微生物發(fā)酵體系,使其呈現(xiàn)清香的風味和透亮的玫瑰色。大紅浙醋主要由糖化階段、酒精發(fā)酵階段以及醋酸發(fā)酵階段發(fā)酵而成[1]。
醋酸菌作為醋酸發(fā)酵階段中的重要微生物,其屬于革蘭氏陰性的需氧菌,能夠快速并不完全氧化大量的碳水化合物和醇,產(chǎn)生有機酸作為最終產(chǎn)物[2]。有機酸決定了食醋的品質(zhì),為食醋清香而酸甜的風味作出了重要貢獻。有趣的是,醋酸菌包含很多新屬和種,每個菌株都有獨特的生長需求。醋酸菌可以利用廣泛的化合物作為氮的來源,如從簡單的無機化合物到復雜的有機化合物,甚至銨離子都可以作為唯一的氮源提供醋酸菌生長代謝。因此,研究不同種類醋酸菌生長的具體營養(yǎng)源濃度和類型是解除其生長限制的重要前提[3-4]。有研究通過單一營養(yǎng)缺乏實驗,確定了天冬氨酸、谷氨酸、脯氨酸和色氨酸是巴氏醋酸桿菌(Acetobacter pasteurianus)CICIM B7003的關(guān)鍵營養(yǎng)氨基酸[5]。還有研究報道,天冬氨酸和谷氨酸能對巴氏醋酸桿菌(Acetobacter pasteurianus)的代謝和抗酸脅迫產(chǎn)生有益影響[6]。此外,有研究發(fā)現(xiàn)氨基酸是通過平衡極端環(huán)境條件下的氧化還原電勢[7],提升糖利用率以及保護特定微生物免受競爭的侵害,以此對微生物的代謝產(chǎn)生影響[8-11]。
為了研究大紅浙醋發(fā)酵中特定醋酸菌的營養(yǎng)需求,本研究首先采用鈣透明圈法從大紅浙醋中分離篩選醋酸菌,并通過分子生物學技術(shù)對其進行菌種鑒定;然后研究10種氮源對巴氏醋酸桿菌產(chǎn)酸量的影響,并對10種氮源組成進行定性和定量分析,得出產(chǎn)酸代謝的關(guān)鍵氨基酸;最后挑出10種關(guān)鍵氨基酸進行驗證,不僅可以了解醋酸菌對氨基酸的需求,還可以通過使用干預手段直接投入關(guān)鍵氨基酸以降低食醋生產(chǎn)成本和提升食醋發(fā)酵效率。
1 材料與方法
1.1 材料與試劑
1.1.1 材料
大紅浙醋樣品:采用無菌方式從廣州致美齋食品有限公司獲取發(fā)酵不同天數(shù)的大紅浙醋樣品。
1.1.2 試劑
5種酵母抽提物(yeast extract,YE)1#YE、2#YE、3#YE、4#YE、5#YE:安琪酵母股份有限公司;動物蛋白胨、小麥蛋白胨(均為生化試劑):北京奧博星生物技術(shù)有限公司;1#大豆蛋白胨、2#大豆蛋白胨(均為生化試劑):北京鴻潤寶順科技有限公司;氫氧化鈉(分析純):天津風船化學試劑有限公司;無水乙醇(分析純):天津市江天化工股份有限公司;普通凝膠脫氧核糖核酸(deoxyribonucleic acid,DNA)回收試劑盒:北京天根生化試劑公司;牛骨蛋白胨(生化試劑)、革蘭氏染色試劑盒、脯氨酸、甘氨酸、丙氨酸、天冬氨酸、谷氨酸、色氨酸、精氨酸、賴氨酸、絲氨酸、纈氨酸(均為分析純):北京索萊寶科技有限公司。
1.1.3 培養(yǎng)基
分離純化培養(yǎng)基[12]:1%1#YE,1%葡萄糖,2%瓊脂粉,2%CaCO3,0.01%硫酸鎂,自然pH,121 ℃滅菌20 min,使用前加入4%(V/V)無水乙醇。
液體發(fā)酵培養(yǎng)基[12]:1%不同種類氮源,1%葡萄糖,0.01%硫酸鎂,自然pH,121 ℃滅菌20 min,使用前加入4%(V/V)無水乙醇。
斜面保藏培養(yǎng)基[12]:1%1#YE,1%葡萄糖,2%瓊脂粉,2%CaCO3,自然pH,121 ℃滅菌20 min。
1.2 儀器與設(shè)備
DY04-13-44-00高壓蒸汽滅菌鍋:上海東亞壓力容器有限公司;MTZ18H009離心機:美國貝克曼庫爾特有限公司;ZHJH-C1115C超凈工作臺:上海智城儀器有限公司;FE28-Standard pH計:梅特勒-托利多有限公司;ZWY-100H恒溫培養(yǎng)箱:上海智城儀器有限公司;UV-1800紫外分光光度計:上海美譜達儀器有限公司;T100聚合酶鏈式反應(yīng)(polymerase chain reaction,PCR)儀、Y-600電泳儀:美國Bio-Rad Laboratories公司;1260 infinityⅡ高效液相色譜(high performance liquid chromatography,HPLC)儀:美國安捷倫公司;Kjeltec 8400全自動凱氏定氮儀:丹麥FOSS公司。
1.3 方法
1.3.1 醋酸菌的分離與篩選
在無菌環(huán)境下稱取25 mL大紅浙醋樣品加入225.0 mL無菌生理鹽水中,常溫下振蕩混勻,梯度稀釋,吸取不同稀釋度的樣品100 μL涂布于分離純化培養(yǎng)基上,在30 ℃培養(yǎng)箱中培養(yǎng)2 d。挑取培養(yǎng)基內(nèi)的單個菌落進行三區(qū)劃線,純化至無雜菌時采用斜面保藏培養(yǎng)基保藏備用[13]。挑取純化好的菌落在載玻片上革蘭氏染色,進行光學顯微鏡觀察[14]。
從固體培養(yǎng)基內(nèi)挑選透明圈較大的菌株接種于液體發(fā)酵培養(yǎng)基(氮源為1#YE),30 ℃、180 r/min條件下培養(yǎng)24 h。取5 mL菌液,4 ℃、8 000 r/min條件下離心,取上清,用1.0 mol/L NaOH調(diào)節(jié)上清液pH為7.0,滴加幾滴5%FeCl3搖勻,隨后在酒精燈火焰上將試管加熱至沸騰,觀察試管內(nèi)是否生成紅褐色沉淀,如產(chǎn)生紅褐色沉淀即為產(chǎn)醋酸菌株[12]。
1.3.2 分子生物學鑒定
采用DNA提取試劑盒提取醋酸菌基因組DNA,以其為模板,采用引物27F(5'-AGAGTTTGATCCTGGCTCAG-3')和1492R(5'-GGTTACCTTGTTACGACTT-3')對分離菌株的16S rDNA基因序列進行PCR擴增。PCR擴增條件:94 ℃預變性2 min;94 ℃變性45 s、55 ℃復性50 s、72 ℃延伸2 min,30個循環(huán),72 ℃修復延伸10 min,于4 ℃終止反應(yīng)。PCR擴增體系:引物27F、1492R各2 μL,模板2 μL,反應(yīng)混合物20 μL,重蒸水(ddH2O)14 μL。PCR擴增產(chǎn)物經(jīng)0.5%瓊脂糖凝膠電泳檢測,檢測合格后委托金唯智生物科技公司進行測序。將測序結(jié)果提交至美國國立生物技術(shù)信息中心(national center for biotechnology information,NCBI)的GenBank數(shù)據(jù)庫中,采用基本局部比對搜索工具(basic local alignment search tool,BLAST)與已知序列進行同源比對,利用MEGA 6軟件中的鄰接(neighbor-joining,NJ)法構(gòu)建系統(tǒng)發(fā)育樹[15-16]。
1.3.3 最佳氮源濃度的確定
在液體發(fā)酵培養(yǎng)基中加入0.5%、1.0%、1.5%、2.0%、5.0%的1#YE作為氮源,按3%(V/V)的接種量接入分離菌株,檢測不同發(fā)酵時間的生物量(OD600nm值)和產(chǎn)酸量[17]。
1.3.4 不同氮源總氮含量和組成成分分析
采用凱氏定氮法測定10種氮源的總氮含量[18]。采用高效液相色譜法對氮源游離氨基酸和非游離氨基酸進行定性和定量測定[19-20]。
1.3.5 不同氮源對分離菌株生物量和產(chǎn)酸量的影響
為了保持不同氮源總氮量恒定,液體發(fā)酵培養(yǎng)基內(nèi)按1%1#YE的總氮量分別添加其他9種氮源,按3%(V/V)的接種量接入分離菌株,檢測不同發(fā)酵時間的生物量(OD600nm值)和產(chǎn)酸量[17]。
1.3.6 氨基酸缺失實驗
通過單氨基酸缺失實驗,確定醋酸菌在酒精醋酸化過程中的必需營養(yǎng)氨基酸。具體步驟如下:設(shè)置包含10種氨基酸的培養(yǎng)基為空白組,再將10種氨基酸分別減少1種(包含9種氨基酸)的培養(yǎng)基作為實驗組,比較空白組和實驗組的產(chǎn)酸量[2]。
1.3.7 數(shù)理統(tǒng)計方法
在實驗過程中每組實驗平行做3次平行后取平均值,最終結(jié)果以“平均值±標準偏差”表示,所有實驗數(shù)據(jù)結(jié)果采用Origin 2021軟件繪圖。
2 結(jié)果與分析
2.1 醋酸菌的分離與篩選
采用分離純化培養(yǎng)基從大紅浙醋中分離得到20株有鈣透明圈的菌株,其中菌株SHQ-A發(fā)酵液產(chǎn)生紅褐色沉淀,初步判定其為產(chǎn)醋酸菌株[17,21],該菌株在分離純化培養(yǎng)基上的菌落及細胞形態(tài)見圖1。
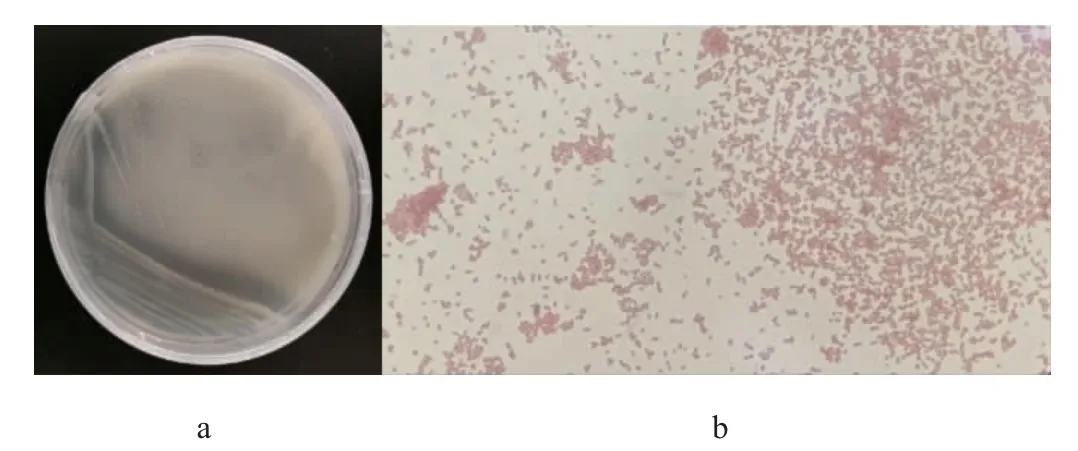
圖1 菌株SHQ-A的菌落(a)及細胞(b)形態(tài)Fig.1 Colony (a) and cell (b) morphology of strain SHQ-A
由圖1可知,菌株SHQ-A的菌落呈圓形,表面呈光滑油脂狀,邊緣整齊且具有透明圈;菌株SHQ-A革蘭氏染色后呈紅色短桿狀,為革蘭氏陰性菌,符合醋酸菌的產(chǎn)酸和革蘭氏陰性特性[22-23]。
2.2 菌株SHQ-A的分子生物學鑒定
基于16S rDNA基因序列,采用MEGA 6軟件構(gòu)建菌株SHQ-A的系統(tǒng)發(fā)育樹,結(jié)果見圖2。
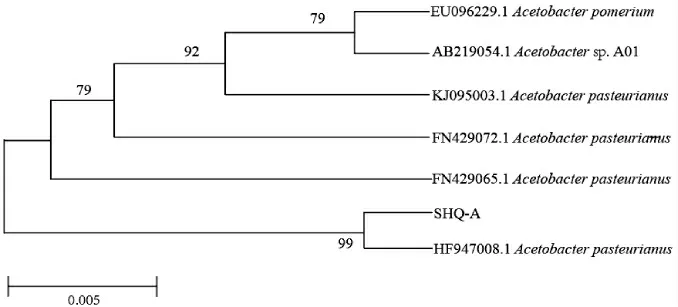
圖2 基于16S rDNA基因序列菌株SHQ-A的系統(tǒng)發(fā)育樹Fig.2 Phylogenetic tree of strain SHQ-A based on 16S rDNA gene sequences
由圖2可知,菌株SHQ-A與Acetobacter pasteurianus(HF947008.1)聚于一支,親緣關(guān)系最近,結(jié)合形態(tài)觀察,鑒定該菌株為巴氏醋酸桿菌(Acetobacter pasteurium)。
2.3 提升醋酸菌SHQ-A發(fā)酵產(chǎn)酸性能的研究
2.3.1 總氮添加量的確定
氮源提供必要氨基酸使醋酸菌生長并進行產(chǎn)酸代謝,當總氮含量過高引起氨基酸濃度過高時不但不會促進醋酸菌生長,更可能抑制它的代謝活動。因此,選擇適宜的總氮添加量是提升巴氏醋酸桿菌產(chǎn)酸的基礎(chǔ)[24]。1#YE添加量對巴氏醋酸桿菌SHQ-A生長和產(chǎn)酸量的影響見圖3。
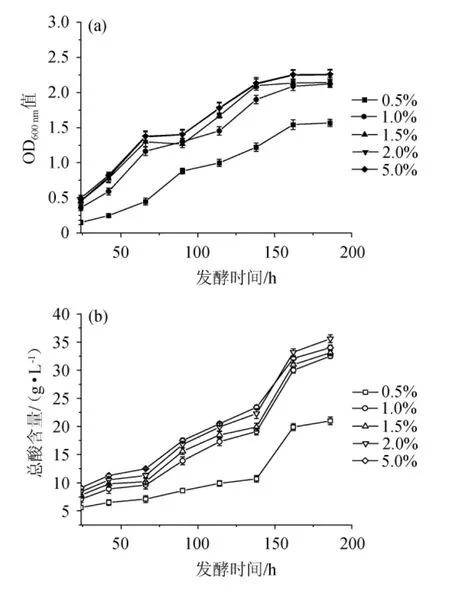
圖3 不同1#YE添加量對巴氏醋酸桿菌SHQ-A生長(a)和產(chǎn)酸量(b)的影響Fig.3 Effects of different 1#YE addition on the growth (a) and acid production (b) of Acetobacter pasteurianus SHQ-A
由圖3可知,當1#YE含量<1.0%時,巴氏醋酸桿菌SHQ-A的生長和產(chǎn)酸量極低。但當1#YE含量>1.0%之后,巴氏醋酸桿菌SHQ-A的生長和產(chǎn)酸量也并無優(yōu)勢。基于高效原則,確定1#YE的最優(yōu)添加量為1.0%。為了確保不同氮源的可比性,根據(jù)表1所測定的不同氮源的總氮含量,將1%1#YE定義為參考培養(yǎng)基,因此,每種培養(yǎng)基總氮添加量都應(yīng)保持為0.11%后添加不同種類氮源。
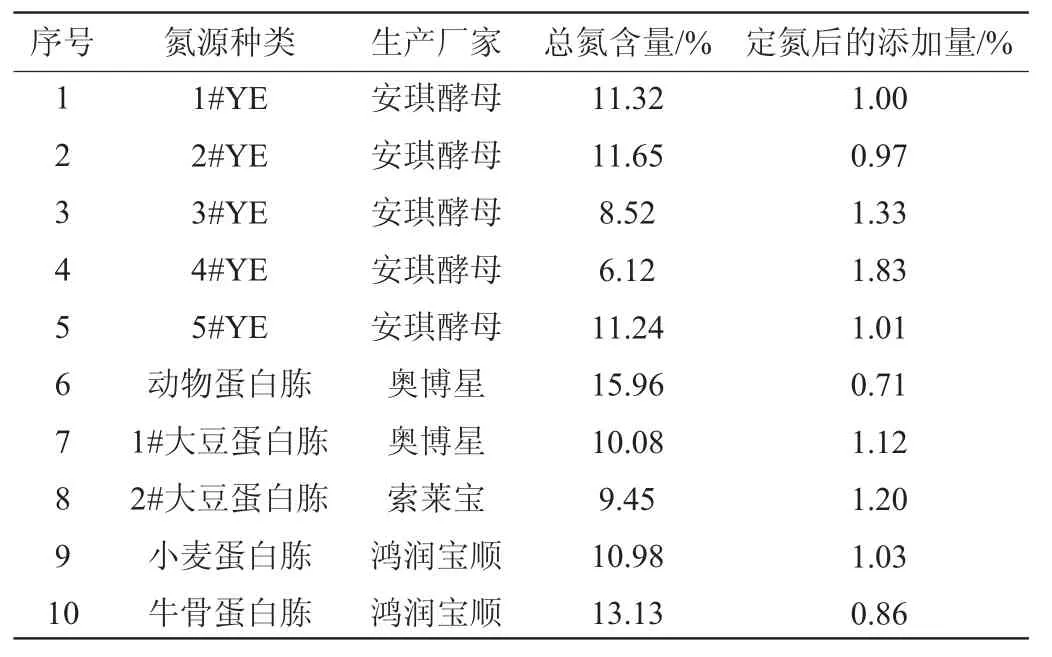
表1 添加不同氮源對總氮含量的影響Table 1 Effect of different nitrogen sources addition on total nitrogen content
2.3.2 不同氮源對菌株SHQ-A生長及總酸含量的影響
不同氮源對巴氏醋酸桿菌SHQ-A生長及產(chǎn)酸代謝的影響見圖4。
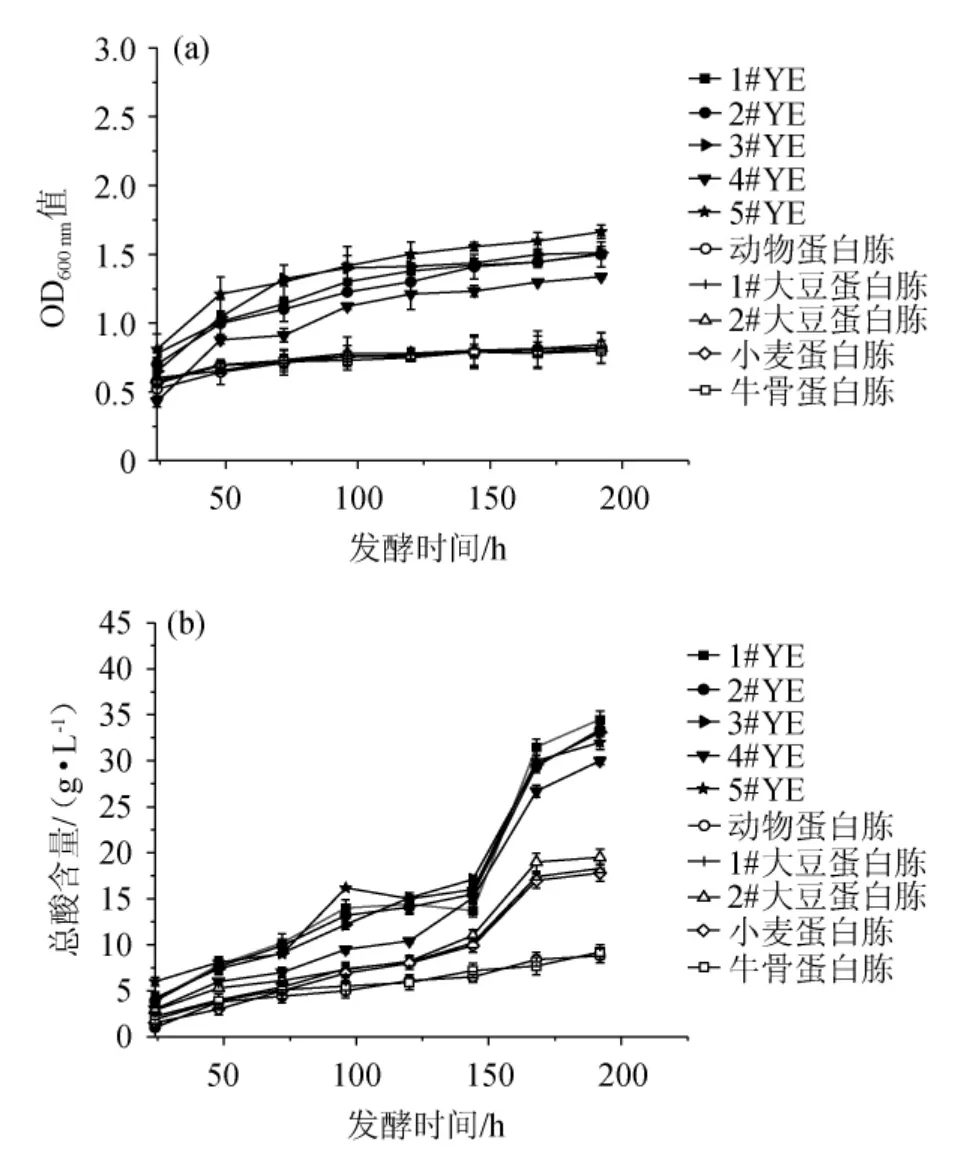
圖4 不同氮源對巴氏醋酸桿菌SHQ-A生物量(a)和產(chǎn)酸量(b)的影響Fig.4 Effects of different nitrogen sources on biomass (a) and acid production (b) of Acetobacter pasteurianus SHQ-A
由圖4可知,相比于蛋白胨,YE明顯對巴氏醋酸桿菌SHQ-A的生長和產(chǎn)酸代謝更具優(yōu)勢。在發(fā)酵終點,以5#YE為氮源時,巴氏醋酸桿菌SHQ-A的OD600nm值最高,為1.7;小麥蛋白胨和牛骨蛋白胨為氮源時,OD600nm值相同且為最低,為0.8。以1#YE為氮源時,巴氏醋酸桿菌SHQ-A的產(chǎn)酸量最高,為34.5 g/L;動物蛋白胨為氮源時,產(chǎn)酸量最低,為8.8 g/L。
2.3.3 不同氮源組成成分分析
為分析不同氮源導致巴氏醋酸桿菌SHQ-A生物量及產(chǎn)酸量差異的原因,進一步對不同氮源的組成成分進行分析,結(jié)果見圖5。
由圖5可知,整體來看YE組中的游離氨基酸的數(shù)量高于蛋白胨組。游離氨基酸和小肽比大肽和蛋白質(zhì)更容易轉(zhuǎn)運到細胞中[25-26]。此外,氨基酸可增強醋酸菌三磷酸合成和氨基酸脫氨,最終提高細胞內(nèi)氨濃度以維持細胞內(nèi)pH的穩(wěn)定性和DNA的修復,促進不飽和脂肪酸的合成和脂質(zhì)轉(zhuǎn)運,從而提高細胞膜硬度、穩(wěn)定性和完整性[6]。
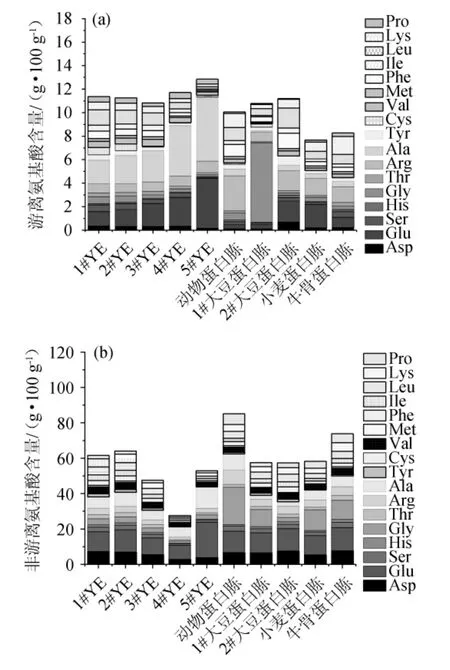
圖5 不同氮源游離氨基酸(a)和非游離氨基酸(b)含量分析結(jié)果Fig.5 Analysis results of free amino acid (a) and non-free amino acid (b) contents of different nitrogen sources
由圖5亦可知,1#大豆蛋白胨、2#大豆蛋白胨游離氨基酸總量與YE相似,但其對巴氏醋酸桿菌SHQ-A的生長和產(chǎn)酸量較YE低,推測可能是游離氨基酸和非游離氨基酸組分的差異導致。YE與蛋白胨組成最大的區(qū)別在于前者含有較高濃度的丙氨酸和游離脯氨酸,后者則含有更多的精氨酸。其中,5#YE中的游離丙氨酸含量最高為5.38 g/100 g,1#大豆蛋白胨中的游離丙氨酸含量最低為0.12 g/100 g。4#YE中的游離脯氨酸含量最高為0.53 g/100 g,2#大豆蛋白胨中的游離脯氨酸含量最低為0.07 g/100 g。動物蛋白胨中的游離精氨酸含量最高為2.98 g/100 g,2#YE中的游離精氨酸含量最低為0.76 g/100 g。1#大豆蛋白胨中含6.78 g/100 g的游離甘氨酸,是5#YE的40.65倍。因此,丙氨酸和脯氨酸很可能是促進產(chǎn)酸代謝的關(guān)鍵積極氨基酸;而甘氨酸和精氨酸可能是抑制產(chǎn)酸代謝的關(guān)鍵消極氨基酸。研究報道,谷氨酰胺、丙氨酸和色氨酸在草莓醋發(fā)酵中呈顯著下降趨勢,可能是因為它們被醋酸細菌消耗,是醋酸細菌和酵母等微生物的良好氮來源。相反,精氨酸和脯氨酸,在發(fā)酵過程中顯著增加。精氨酸顯著增加的可能原因是該氨基酸在亞硝基條件下儲存在微生物液泡中,自溶解后留在培養(yǎng)基中[3]。
2.3.4 氨基酸缺失實驗
為了闡明巴氏醋酸桿菌SHQ-A的具體營養(yǎng)需求,根據(jù)上述推測結(jié)果和巴氏醋酸桿菌中心氨基酸代謝網(wǎng)絡(luò)[5](圖6)找出10種關(guān)鍵氨基酸以作具體研究,結(jié)果見圖7。
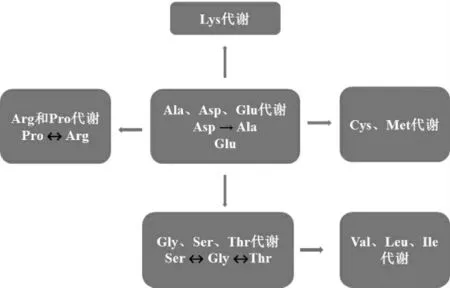
圖6 巴氏醋酸桿菌SHQ-A的中心氨基酸代謝網(wǎng)絡(luò)圖Fig.6 Central amino acid metabolic network of Acetobacter pasteurianus SHQ-A
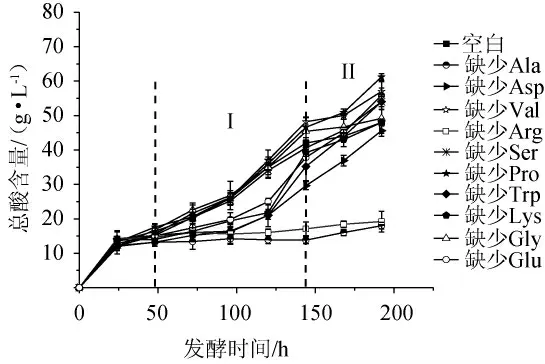
圖7 缺少單一氨基酸時巴氏醋酸桿菌SHQ-A的產(chǎn)酸量Fig.7 Acid production of Acetobacter pasteurianus SHQ-A without single amino acid
由圖7可知,在發(fā)酵終點時,空白組中總酸含量為48.0 g/L。當缺少丙氨酸和精氨酸時總酸含量顯著降低,分別為18.0 g/L和19.2 g/L,比空白組低62.5%和60%。因此,可以確定丙氨酸和精氨酸是巴氏醋酸桿菌SHQ-A代謝產(chǎn)酸的關(guān)鍵積極氨基酸。可能是因為巴氏醋酸桿菌SHQ-A的產(chǎn)酸只需要少量的精氨酸,所以蛋白胨中精氨酸高于YE中精氨酸的含量沒有產(chǎn)生實際作用。在氨基酸脫羧逆向轉(zhuǎn)運蛋白反應(yīng)中,氨基酸被轉(zhuǎn)運到細胞內(nèi)并被相關(guān)酶脫羧,然后脫羧基通過反向轉(zhuǎn)運蛋白從細胞中輸出,同時消耗一個質(zhì)子[27]。精氨酸去氨酸酶途徑,通過堿性物質(zhì)增加了細胞內(nèi)的pH,可能有助于修復由酸應(yīng)激引起的損傷或其他應(yīng)激引起的脫氧核糖核酸(DNA)的損傷[27-28]。因此,當營養(yǎng)源中缺乏精氨酸便會直接導致產(chǎn)酸受到限制。有趣的是,當發(fā)酵體系中缺乏脯氨酸時,不但不會抑制巴氏醋酸桿菌SHQ-A產(chǎn)酸代謝,產(chǎn)酸量反而比空白組高27.1%,推測脯氨酸可能是產(chǎn)酸代謝的關(guān)鍵消極氨基酸。但是在植物學研究中,脯氨酸可以穩(wěn)定生物大分子結(jié)構(gòu),降低細胞酸度,調(diào)節(jié)細胞膜氧化還原電位[5]。
為了更好的表明空白組與實驗組產(chǎn)酸量的差異分布,分為I和II發(fā)酵階段。由圖7亦可知,在Ⅰ區(qū)內(nèi),谷氨酸、賴氨酸、天冬氨酸和色氨酸缺失后產(chǎn)酸量相較于空白組低,但隨著時間延長(Ⅱ區(qū))其限制影響消失。氨基酸主要在指數(shù)生長階段被使用,因此Ⅰ區(qū)內(nèi)一些氨基酸缺乏導致了產(chǎn)酸代謝受到限制。但是考慮到氨基酸之間可以相互轉(zhuǎn)化,所以某些氨基酸的單一缺乏在發(fā)酵后期可能不足以導致產(chǎn)酸量降低。如在巴氏醋酸桿菌的中心氨基酸代謝中天冬氨酸可轉(zhuǎn)化為谷氨酸和丙氨酸,由此可以降低谷氨酸和丙氨酸缺失時產(chǎn)生的不良影響[5]。根據(jù)其他研究報道,在蘋果酒中加入谷氨酸或天冬氨酸可明顯促進菌生長,這可能是由于三羧酸循環(huán)(tricarboxylic acid cycle,TCA)周期的增強,但最終酸度較低。色氨酸有利于在發(fā)酵過程中穩(wěn)定乙醇脫氫酶(alcohol dehydrogenase,ADH)和乙醛脫氫酶(aldehyde dehydrogenase,ALDH)的活性,這可能會導致最終的酸度增加。根據(jù)京都基因與基因組百科全書(Kyoto Encyclopedia of Genes and Genomes,KEGG)數(shù)據(jù)庫可知,色氨酸參與了泛素的合成,這是醋酸細菌中酒精呼吸鏈中唯一的一個自由電子轉(zhuǎn)運體,色氨酸可能會加強酒精呼吸鏈中的電子轉(zhuǎn)運,從而增強腺嘌呤核苷三磷酸(adenosinetriphosphate,ATP)的合成,促進酒精酰化過程中乙酸的細胞應(yīng)激性[29]。但在本研究中,缺乏色氨酸時并未對巴氏醋酸桿菌SHQ-A產(chǎn)酸代謝產(chǎn)生影響。
3 結(jié)論
本研究首先采用鈣透明圈法從大紅浙醋中分離篩選得到一株產(chǎn)酸菌株,編號為SHQ-A,經(jīng)形態(tài)觀察和分子生物學鑒定為巴氏醋酸桿菌(Acetobacter pasteurianus)。該菌株利用10種氮源發(fā)酵的產(chǎn)酸量在8.8~34.5 g/L范圍內(nèi),且以酵母抽提物為氮源的產(chǎn)酸量明顯高于蛋白胨。當缺少纈氨酸、組氨酸、色氨酸、甘氨酸、天冬氨酸、賴氨酸、谷氨酸時,產(chǎn)酸量與空白組無明顯差異。然而,當缺少丙氨酸和精氨酸時,產(chǎn)酸量僅為18.0 g/L和19.2 g/L,分別低于空白組62.5%和60%,表明丙氨酸和精氨酸是產(chǎn)酸代謝的關(guān)鍵積極氨基酸;當缺少脯氨酸時,產(chǎn)酸量為61.2 g/L,高于空白組27.1%,表明脯氨酸是產(chǎn)酸代謝的關(guān)鍵消極氨基酸。
