國蘭葉色突變體根狀莖差異表達基因分析
2022-03-11 04:52:56何俊蓉
核農學報 2022年3期
關鍵詞:差異
蔣 彧 何俊蓉
(四川省農業科學院園藝研究所,四川 成都 610000)
蘭科是單子葉植物第一大科,包含了5個亞科801個屬,品種數量近3萬種,約占所有種子植物種類的10%[1]。我國十大名花中的蘭花也稱國蘭,屬于蘭科(Orchidaceae)蘭屬(Cymbidium)小花型地生種,在我國有悠久的栽培歷史,是世界上最早栽培的蘭花。蘭花是高產值的花卉,隨著我國經濟及出口貿易的發展,全球蘭花市場逐漸繁榮,因此對蘭花的育種工作也提出了更高的要求。
蘭花育種的常用手段包括選擇育種、雜交育種、倍性育種、體細胞無性系變異以及分子育種等。我國蘭花品質相關的育種以選擇育種及雜交育種為主,但這兩種育種方式的育種周期長、工作量大,近年來也開始采用如倍性育種、體細胞無性系變異等方法。隨著測序技術及基因編輯技術的發展,分子育種技術在水稻、玉米、棉花等農作物中被廣泛使用,并取得了很好的育種效果[2]。蘭花分子育種起步較晚,蘭科植物如小蘭嶼蝴蝶蘭(Phalaenopsisequestris)[3]、鐵皮石斛(Dendrobiumofficinale)[4]、深圳擬蘭(Apostasiashenzhenica)[5]、白花蝴蝶蘭(Phalaenopsisaphrodite)[6]的基因組發布將有效促進蘭花分子育種工作的開展。近年來,基于多組學數據的分析,蘭花重要性狀相關基因的挖掘取得了重要進展。蘭花中受低溫抑制的CgSVP[分離自春蘭(cymbidumgoeringii)]參與了蘭花低溫開花調控,該調控機制與模式植物中的低溫響應機制存在顯著差異[7]。蘭花中的MADS-box基因家族參與了國蘭特異性唇瓣分化和重瓣化的調控,通過表達區域擴增產生花型的變異[8-9]。通過CRISPR/Cas9基因編輯技術,研究者也成功獲得了蘭科植物鐵皮石解C3H、C4H[10]以及蝴蝶蘭MADS8/36/44的突變株系[11]。上述研究表明分子育種技術在蘭花育種中能夠發揮重要作用。
葉藝是國蘭特有的一種體征,表現為葉片呈現金銀色的條紋或斑點,具有較高的觀賞價值,葉藝蘭的價格也在逐步提高,其消費市場具有巨大潛力。目前市場上銷售的葉藝蘭品種大部分屬于野生的變異種(也稱為下山蘭,即直接從山上挖回的蘭花),另有少部分來自于栽培品種芽變,但前者會對蘭花野生資源造成破壞,后者變異率較低。人工誘變技術較栽培品種芽變具有更高的變異率,對蘭花的育種工作起到了重要的推動作用。葉藝突變體成因復雜,目前的研究認為葉綠體發育缺陷和葉綠素過量降解是葉藝性狀產生的直接生理原因[12]。墨蘭葉片顏色在由紅變黃,最后轉化為綠色的過程中,有28種代謝物在葉片變綠后含量降低到近零水平,還有15種花色素苷類代謝物含量呈下降趨勢[13]。CsERF2則通過乙烯信號途徑參與調控墨蘭(Cymbidumsinense)葉色的變化[14]。但總的來說,導致國蘭葉藝表型背后的分子機制還有待進一步研究。
研究認為,60Co-γ射線可有效地對植物進行誘變產生突變體[15],通過60Co-γ射線對隆昌素根狀莖進行誘變后可獲得能產生葉藝苗的葉藝隆昌素。隆昌素與葉藝隆昌素根狀莖分別表現為綠色和黃色,具有非常明顯的差異,這兩種材料中的葉綠素與類胡蘿卜素含量有較大差別[2]。本研究采用RNA-Seq技術對這兩種根狀莖進行分析,鑒定在隆昌素與葉藝隆昌素根狀莖中差異表達的基因,旨在為解析葉藝隆昌素葉藝成因的分子機制奠定研究基礎。
1 材料與方法
1.1 試驗材料與試劑
試驗材料來自四川省農業科學院園藝研究所組培室。培養溫度為23℃(白天)/18℃(晚上),光照周期為14 h(光照)/10 h(黑暗),光照強度為2 000 lx。無菌條件下隨機選取葉藝隆昌素(YR)和隆昌素(GR)根狀莖各4瓶,分別從每個培養瓶中隨機挑選1~2 cm根狀莖各4條,置于液氮中冷凍后,-80℃超低溫冰箱貯存備用。
多糖多酚植物總RNA提取試劑盒(離心柱型),北京天根生化科技有限公司;mRNA富集用的磁珠mRNA Capture BeadsRNA,南京諾唯贊生物科技有限公司;逆轉錄試劑盒 TUREscript 1st Strand cDNA Synthesis Kit,北京艾德萊生物科技有限公司;2×SYBR?Green Supermix實時熒光定量PCR試劑盒,德國DBI?Bioscience。
1.2 儀器與設備
Eppendorf高速冷凍離心機(Eppendorf,德國);Nanodrop2000超微量分光光度計(Thermo Scientific,美國);qTOWER2.2實時PCR系統(DBI?Bioscience,德國)
1.3 試驗方法
1.3.1 總RNA提取 根據植物總RNA提取試劑盒說明書,從根狀莖組織樣品中提取總RNA,利用Nanodrop2000超微量分光光度計檢測RNA的濃度和純度,1.5%瓊脂糖凝膠電泳檢測RNA完整性。
1.3.2 cDNA文庫的構建 使用Oligo dT磁珠法從總RNA中分離出mRNA,用超聲波破碎法將mRNA隨機斷裂成200 bp左右的小片段。在逆轉錄酶的作用下合成第一鏈cDNA,在DNA聚合酶作用下進一步合成第二鏈cDNA。
1.3.3 轉錄組測序及差異表達基因篩選 將構建好的cDNA送至上海美吉生物醫藥科技有限公司使用Illumina Hiseq2500進行轉錄組測序,測序方式為雙末端法,測序長度為150 bp。測序數據經質控,截除測序接頭、引物序列,并對低質量值數據進行過濾,最終獲得高質量數據(reads)。使用Trinity軟件[16]對高質量Reads進行denovo拼接獲得單基因(unigene)。采用RSEM軟件(V 1.2.4)中的FPKM(fragments per kilobase per million reads)方法對unigene表達量水平進行計算,使用Bioconductor軟件包中的DESeq(V 1.14.0)進行基因的差異表達分析。差異表達基因(different expression genes, DEGs)篩選閾值為|log2 value|>1且FDR≤0.05。
1.3.4 unigene功能注釋及基因結構分析 使用BlastX將unigene與NR、GO、KOG數據庫進行比對,根據基因的相似性進行功能注釋,得到與給定unigene具有最高序列相似性的蛋白,從而得到該unigene的蛋白功能注釋信息。使用TransDecoder軟件(transdecoder.github.io/)對unigene的編碼區及其對應的氨基酸序列進行預測。
1.3.5 差異表達基因富集分析 分別使用GOseq與KOBAS對差異表達基因進行GO功能顯著性及KEGG Pathway富集分析。
1.3.6 實時熒光定量PCR(quantitative real-time PCR, qRT-PCR) 提取根狀莖RNA,用Aidlab公司反轉錄試劑盒(TUREscript 1st Strand cDNA Synthesis Kit)進行cDNA的合成。從轉錄組數據中篩選出準備驗證的unigene,分別設計這些unigene的特異引物,采用qTOWER2.2實時PCR系統進行qRT-PCR分析,所有反應均設置3個生物學重復和技術重復。qRT-PCR反應體系為10 μL:5 μL 2×SYBR?Green Supermix,0.5 μL 200 nmol·L-1正向引物,0.5 μL 200 nmol·L-1反向引物,1 μL cDNA模板,3 μL ddH2O。qRT-PCR反應程序為:95℃預變性3 min;95℃變性10 s,60℃擴增30 s,循環39次;溶解曲線分析,即隨后將溫度從60℃逐漸升至95℃(每次提高1℃,每提高1℃停留4 s)。ACTIN-7基因作為內參,通過2-△△Ct法計算目的基因相對表達量。
2 結果與分析
2.1 葉藝隆昌素和隆昌素根狀莖形態
葉藝隆昌素根狀莖呈淡黃色,形狀較為粗短(圖1-A);隆昌素根狀莖則呈深綠色,形狀為不規則長條形(圖1-B)。經過培養后,淡黃色的根狀莖分化出葉藝苗,綠色的則只分化成正常的綠色試管苗。

圖1 葉藝隆昌素根狀莖(A)和隆昌素根狀莖(B)觀察Fig.1 Observation of Yeyi Longchangsu (A) and Longchangsu (B) rhizome
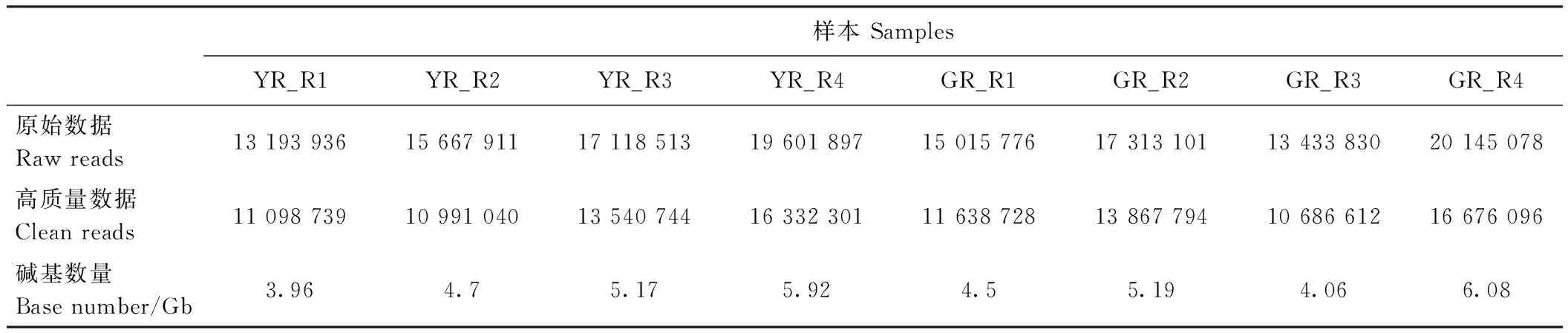
表1 測序結果Table 1 Sequencing statistics
2.2 測序數據組裝與分析
分別對葉藝隆昌素和隆昌素的根狀莖進行轉錄組測序,每個樣本分別進行4次重復,8個文庫得到的原始數據(raw reads)條數介于13 193 936~20 145 078,質控后的高質量數據(clean reads)條數介于10 686 612~16 676 096,clean reads占raw reads的比例為70.15%~84.12%,clean reads數據量大小為3.96~6.08 Gb(表1)。使用Trinity和TGICL軟件對clean reads進行從頭組裝,共得到162 407條unigene(表2)。unigene的N50長度和平均長度分別為1 049和678 bp。unigene長度分布結果顯示大于1 000 bp的unigene有26 107個(圖2)。
2.3 基因功能注釋及分類
基因功能注釋與分類為闡明細胞內特定基因的分子功能和生物代謝途徑提供了有價值的信息。將162 407 條unigene與Nr、GO和KOG數據庫進行比對,獲得unigene的注釋信息。結果顯示有26 473條unigene注釋到Nr數據庫,有20 119條unigenes注釋到KOG數據庫,有38 926條unigene注釋到GO數據庫(圖3),其中有14 380條unigene在3個數據庫中均有注釋。在GO部分,注釋到細胞組分、分子功能、生物學過程的unigene數目分別為12 114、13 170、13 642 條。
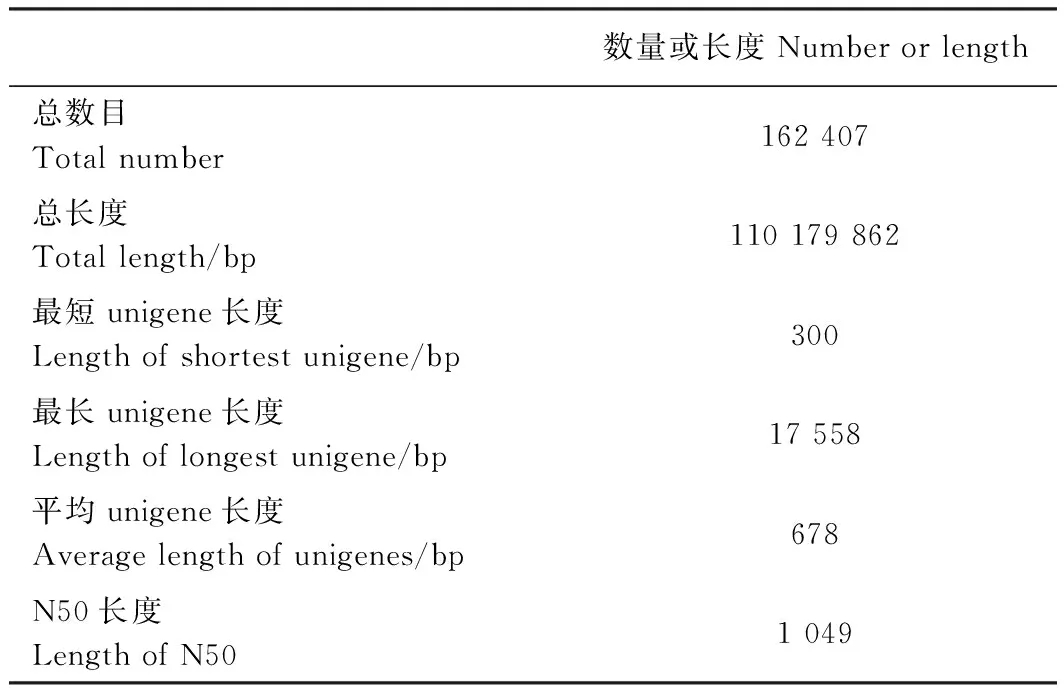
表2 轉錄組數據的組裝及unigene預測Table 2 Statistics of transcriptome assembly and predicted unigenes
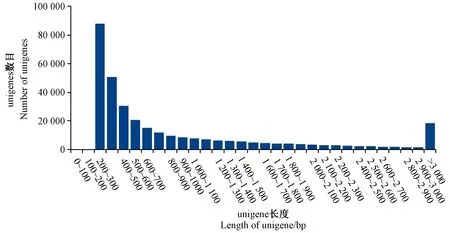
圖2 unigene長度分布圖Fig.2 Length distribution of unigene
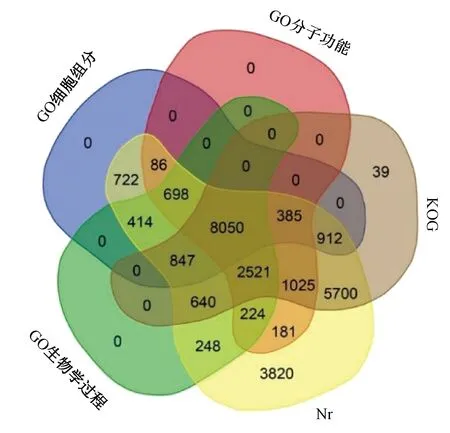
圖3 GO、Nr和KOG數據庫注釋結果統計Fig.3 GO, Nr and KOG database annotation results statistics
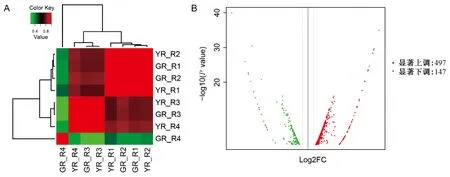
圖4 相關性熱圖(A)和差異表達基因火山圖(B)Fig.4 Correlation heat map (A) and volcano map of DEGs (B)
2.4 差異表達基因的鑒定及功能分類
在閾值為|log2 value|>1和FDR≤0.05情況下,鑒定出644個unigene在葉藝隆昌素和隆昌素根狀莖中差異表達,其中497個unigene的轉錄本在葉藝隆昌素中表達豐度較高,147個unigene的轉錄本在隆昌素中表達豐度較高(圖4)。
對差異表達基因進行GO聚類,結果表明有42個類別顯著富集,其中屬于生物學過程的有20個,屬于細胞組分的有11個,屬于分子功能的有11個(圖5)。在生物學過程類別中,代謝過程富集度最高,節律過程富集度最低;在細胞組分類別中,細胞富集度最高,膜封閉腔富集度最低;在分子功能類別中,催化活性富集度最高,金屬伴侶活性富集度最低。
對差異表達基因進行KEGG聚類,結果顯示有8個代謝過程被顯著富集(P<0.05)(表3)。這些過程涉及植物-病原物互作(P=0.000 26)、礦物質吸收(P=0.001 34)、光形態建成(P=0.001 99)、倍半萜和三

注: 1: 生物調控; 2: 細胞成分組織或起源; 3: 細胞過程; 4: 發育過程; 5: 定位系統的建立; 6: 生長; 7: 免疫系統過程; 8: 定位; 9: 代謝過程; 10: 多機體過程; 11: 多細胞組織過程; 12: 生物過程負調控; 13: 生物過程正調控; 14: 生物過程調控; 15: 再生; 16: 再生過程; 17: 刺激應答; 18: 節律過程; 19: 信號; 20: 單一的生物過程; 21: 細胞; 22: 胞間連絲; 23: 細胞要素; 24: 細胞間區域; 25: 大分子復合物; 26: 膜; 27: 膜要素; 28: 膜結合腔體; 29: 細胞器; 30: 細胞器要素; 31: 共質體; 32: 抗氧化劑活性; 33: 結合劑活性; 34: 催化劑活性; 35: 電荷載體活性; 36: 酶調控因子活性; 37: 金屬伴侶活性; 38: 分子感應器活性; 39: 核苷酸結合轉錄因子活性; 40: 蛋白質結合轉錄因子活性; 41: 結構分子活性; 42: 轉運因子活性。Note: 1: Biological regulation. 2: Cellular component organization or biogenesis. 3: Cellular process. 4: Developmental process. 5: Establishment of localization. 6: Growth. 7: Immune system process. 8: Localization. 9: Metabolic process. 10: Multi-organism process. 11: Multicellular organismal process. 12: Negative regulation of biological process. 13: Positive regulation of biological process. 14:Regulation of biological process. 15: Reproduction. 16: Reproductive process. 17: Response to stimulus. 18: Rhythmic process. 19: Signaling. 20: Single-organism process. 21: Cell. 22: Cell junction. 23: Cell part. 24: Extracellular region. 25: Macromolecular complex. 26: Membrane. 27: Membrane part. 28: Membrane-enclosed lumen. 29: Organelle. 30: Organelle part. 31: Symplast. 32: Antioxidant activity. 33: Binding activity. 34: Catalytic activity. 35: Electron carrier activity. 36: Enzyme regulator activity. 37: Metallochaperone activity. 38: Molecular transducer activity. 39: Nucleic acid binding transcription factor activity. 40: Protein binding transcription factor activity. 41: Structural molecule activity. 42: Transporter activity.圖5 不同表達基因的GO注釋分析Fig.5 The GO annotation statistical of DGEs

表3 DEG基因KEGG分析Table 3 The KEGG path way of DEG
萜生物合成(P=0.047 25)。
2.5 葉綠素與類胡蘿卜素相關基因的鑒定及表達分析
葉綠素、類黃酮、類胡蘿卜素是決定植物色澤的三大主要物質。本試驗通過unigene注釋結果進行篩選,分別獲得了15編碼參與葉綠體與12個編碼參與類胡蘿卜素生物合成的關鍵酶的基因。對15個葉綠素相關基因的表達分析發現,其中5個基因在兩種實驗材料中表達水平基本一致,10個基因表達水平存在一定差異,但未達到顯著水平(表4)。與類胡蘿卜素相關的12個unigene中(表5),編碼類胡蘿卜素解離雙加氧酶7(CDD7)的基因c1920_g1在葉藝隆昌素中表達下調。

表4 葉綠素相關unigene鑒定Table 4 Identification of chlorophyll related unigene

表5 類胡蘿卜素相關unigene鑒定Table 5 Identification of carotenoid related unigene
2.6 葉綠體發育相關基因的鑒定及表達分析
從轉錄組測序結果中篩選出一些編碼光系統I/II、 細胞色素b6f復合物、ATP合酶的unigene,其中光系統Ⅱ中的psbC、psbH以及光系統I中的Ycf3基因在葉藝隆昌素根狀莖中均表現為表達水平下調(表6)。葉綠體自身編碼的第二類基因是和葉綠體DNA復制與葉綠體基因表達相關的基因。DEG分析結果顯示,編碼RNA聚合酶亞基的glgC、rbcL以及編碼核糖體蛋白亞基的rpoB、rpsR基因在葉藝隆昌素根狀莖中均表現為表達下調(表7)。
2.7 qRT-PCR驗證
為了驗證轉錄組數據的可靠性,本試驗選擇了9個涉及葉綠素生物合成、光合系統和葉綠體發育的基因設計引物(表8)進行qRT-PCR反應,對轉錄組分析

表6 光合作用相關的葉綠體編碼基因篩選Table 6 Identification of chloroplast structure related unigenes

表7 葉綠體結構相關的基因篩選Table 7 Identification of chloroplast structure related unigenes
結果進行驗證,如圖6所示。qRF-PCR結果顯示驗證基因的表達趨勢均與轉錄組數據相符合,表明轉錄組數據具有準確性與可信性。
2.8 其他調控相關的差異基因
除了上述與葉綠素或類胡蘿卜素相關的差異基因,轉錄組測序結果還獲得了大量與植物激素信號傳導、蛋白轉運、編碼細胞色素P450蛋白、次生物質代謝、內源激素相關的差異表達基因(表9)。這些基因也可能影響葉藝隆昌素的葉色表型。
3 討論
目前已有研究對蘭科植物進行了基因組測序,其花色變異及分子標記得到了部分解析[8,17-18],但大多數蘭花物種的遺傳信息尚不清楚。轉錄組代表組織特定發育時期或階段的所有轉錄本的總和,對于從宏觀層面揭示分子調控網絡有著獨特的優勢[19-21]。隆昌素和葉藝隆昌素試管苗根狀莖的顏色呈現明顯不同,前者呈墨綠色,后者呈黃色。通過對葉藝蘭分化的研究發現,黃色根狀莖可以分化出葉藝苗,綠色根狀莖則分化正常綠色試管苗,是研究國蘭葉藝形成機制的重要材料[2]。本研究通過轉錄組分析鑒定到644個在隆昌素和葉藝隆昌素根狀莖中差異表達的unigene,通過對這些差異表達基因的篩選,初步分析了影響葉藝隆昌素根狀莖變黃的因素,為解析國蘭葉藝形成分子機制提供了一定的科學依據。
植物的色澤與類胡蘿卜素、類黃酮、葉綠素含量有關[22],植物葉綠素的合成過程要經歷大約16個步驟,其中以谷氨酰t-RNA為起點,過程中涉及16種酶和20多個基因[23],這些基因會影響葉綠素合成,進而使葉綠體中各種色素的比例與含量發生變化,最終改變葉色。水稻中已有35個葉色相關基因得到克隆,如水稻ygl1黃綠葉突變體幼葉中葉綠體因為葉片內四吡咯中間體含量的變多而發育遲緩,導致其葉綠素含量遠低于野生型[24]。相關研究結果顯示,國蘭葉片顏色變異由葉綠素代謝途徑異常而非合成途徑異常導致[12]。本研究中篩選出的編碼葉綠素合成相關酶的基因在葉藝隆昌素與隆昌素根狀莖中也未檢測出顯著差異,與前人研究結果相似。
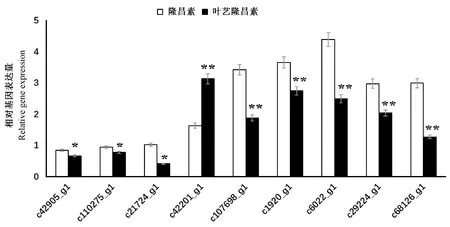
注:*表示具有顯著差異(P<0.05), ** 表示具有極顯著差異(P<0.01)。Note: * and ** indicate significant at 0.05 level and very significant difference at 0.01 level, respectively.圖6 unigene qRT-PCR驗證結果Fig.6 qRT-PCR identification of unigenes

表8 qRT-PCR引物設計Table 8 qRT-PCR primers
CCD7編碼一個ABA生物合成途徑中的關鍵限速酶。研究表明,植物體內源ABA的變化與CCD7表達變化存在顯著相關性[25]。另一方面,花瓣顏色的形成與CCDs的表達水平存在顯著相關性[25],菊花白色花瓣中CCD4表達水平遠高于黃色花瓣,CCD4沉默后的白色花瓣會轉變為黃色,原因是白色花瓣合成類胡蘿卜素之后進行了降解。在葉藝隆昌素根狀莖內,CCD7的表達量下調,可使胡蘿卜素降解速度變快,從而使得根狀莖顏色發黃,進一步說明CCD的表達參與了顏色調控。
捕光色素蛋白復合體由類囊體蛋白與分布在葉綠體類囊體膜上的光合色素組成,這些色素蛋白復合體均參與了光反應,起到轉換能量與傳遞電子的作用[26]。本研究中光系統Ⅱ中的psbC、psbH以及光系統Ⅰ中Ycf3基因在葉藝隆昌素根狀莖中表達量下調。這些基因的表達下調可能影響了葉綠體基粒片層結構,造成光合電子傳遞速率下降,光合能力下降,進而影響根狀莖增殖及分化效率。光合作用碳固定和光呼吸碳氧化過程中的關鍵酶為1,5-二磷酸核酮糖羧化酶/加氧酶(Rubisco),其大亞基編碼通過葉綠體基因實現,是植物葉片內的關鍵儲存蛋白。有研究顯示,水稻(OryzasativaL.)幼苗白化大多由Rubisco小亞基因的突變導致[27-28]。編碼Rubisco的unigene在葉藝隆昌素內的表達量會減少也可能是導致葉藝隆昌素根狀莖變黃的原因之一。前人研究發現,大亞基與小亞基是核糖體的主要組成要素,煙草(NicotianatabacumL.)葉片因為缺失葉綠體核糖體小亞基編碼基因rps18而出現了畸形并變白的現象,存活率極低;Fleischmann等[29]通過研究發現,葉片形態會因為缺失核糖體大亞基編碼基因rpl36而發生變化,光合速率和植株生長速

表9 其他相關基因篩選Table 9 Identification of others related unigenes

表9 (續)
度明顯變慢,本研究中編碼核糖體蛋白大小亞基的rpoB、rpsR基因葉藝隆昌素根狀莖均表達下調,而這些基因是否突變或缺失引起葉綠體結構破壞,有待進一步研究確定。
ABC轉運蛋白參與了細胞器(如液泡、過氧化物體、線粒體、葉綠體等)間次生代謝產物、植物激素、蛋白質、氨基酸、糖類等物質的轉運,是一種跨膜運輸蛋白,存在于大部分生物膜上[30-31]。本研究中,ABC轉運蛋白家族基因(ABCB1與ABCG2)表達下調,由此推測由于葉藝隆昌素根狀莖顏色變黃、葉綠體結構破壞,導致光合作用減弱、產生的次生代謝物質減少,從而通過信號轉導途徑反饋抑制ABC轉運蛋白基因的表達。線粒體與葉綠體在植物細胞中都具有重要作用,線粒體內膜中會出現氧化磷酸化的現象,在ATP合酶催化作用下,電子在線粒體內膜上一系列電子傳導作用下傳遞到氧,促使ADP和磷酸合成ATP,線粒體內膜上的ATP合成酶、細胞色素氧化酶、輔酶Q-細胞色素bc1還原酶、琥珀酸脫氫酶、NADH脫氫酶五種蛋白復合體都屬于電子傳遞體,其中線粒體電子傳遞鏈內以NADH脫氫酶和NADH-泛醌氧化還原酶最為重要,電子傳遞速率受到這些酶活性的影響。本試驗中,NADH脫氫酶、NADH-泛醌氧化還原酶和細胞色素氧化酶均表達下調,可以推測由于葉藝隆昌素根狀莖葉綠體結構不完整,導致體內新陳代謝減弱,通過信號傳導途徑抑制NADH脫氫酶、NADH-泛醌氧化還原酶和細胞色素氧化酶基因的表達,減少ATP的合成。作為折疊水解酶超家族成員的BDG,其主要在皮細胞中表達,擬南芥純合突變植株矮小、葉片異常、表皮毛細胞數不斷變小且細胞萎陷,存在許多畸型細胞,植株發育緩慢,推測其表皮細胞組織是在某種未知調節機制的作用下實現分化和增殖的[32]。本研究中,葉藝隆昌素根狀莖類聯苯水解酶蛋白顯著上調,但該蛋白的作用機制尚不清楚,可知其功能可能與角質的生物合成或與細胞壁本身交聯酶相關,同時該蛋白是否與根狀莖顏色變黃相關有待進一步證實。
4 結論
本研究采用轉錄組分析發現,隆昌素與葉藝隆昌素根狀莖差異基因在植物-微生物互作、礦物質吸收、光形態建成與倍半萜和三萜生物合成通路中顯著富集。此外,參與ABA合成的CDD7、光系統Ⅰ中的Ycf3、光系統Ⅱ中的psbC與psbH、編碼RNA聚合酶亞基的glgC與rbcL、編碼核糖體蛋白亞基的rpoB與rpsR等基因表達量的改變可能通過影響植株葉綠體發育從而影響植株的葉藝表型。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
