不同季節桃金娘光合特性與光系統PSII活性研究
2022-03-15 09:07:14楊通文高秀梅韓維棟
西南農業學報 2022年12期
關鍵詞:植物
楊通文, 高秀梅, 韓維棟
(廣東海洋大學濱海農業學院, 廣東 湛江 524088)
【研究意義】光合作用是植物有機物積累和代謝的重要途徑,是有機物生產的基礎,決定植物的生產力[1],光合作用受溫度、光照輻射、蒸騰速率、氣孔導度、胞間CO2濃度等因子影響,這些因子隨季節變化而改變,在不同季節里生態因子對植物光合作用影響程度也會發生改變[2]。植物光合作用內的凈光合速率、蒸騰速率、氣孔導度、胞間CO2濃度等光合參數能夠反應植物對光能的利用能力[3]。植物葉片內葉綠素熒光參數包含豐富的光合作用信息[4-5],能夠更加直觀地展示植物光化學活性,反應葉片光合作用過程中光系統對光能的吸收、傳遞、耗散、分配等方面具有獨特作用, 與“表觀性”的氣體交換指標相比, 更能反映“內在性”特點[6]。葉綠素熒光測定技術能夠靈敏反應植物光系統PSII反應中心氧化-還原狀態、類囊體膜的高能狀態和色素間的能量傳遞效率[7];植物熒光變化容易受到外界環境的影響,可以直接或間接了解植物光合作用過程,反應植物光合能力的重要參數[8]。葉綠素熒光測定技術具有快速、靈敏、非破壞測量等優點,在植物生理研究、作物育種、栽培管理等領域中得到廣泛應用[9-11]。桃金娘(Rhodomyrtustomentosa)是桃金娘科,屬灌木,野生桃金娘資源在我國廣泛分布于海南、廣西、云南、江西、臺灣、貴州和湖南南部地區[12-13]。果實含有多種人體需要的微量元素和多糖、多酚以及黃酮類有機物具有抗氧化、抗菌、抗病毒、降糖和抑制乙酰膽堿酶活性等藥理作用[14-16];桃金娘的株型大小與生長環境相適應,葉小且革質化,根系發達,耐高溫和干旱,對水肥要求不高,移栽容易成活;開花期長達到一個月以上,在氣溫、水肥管理以及修剪合適的情況下,葉片能夠不斷新發出來,在城市綠化建設、園林應用等方面也具有潛在價值[17]。【前人研究進展】國內外關于桃金娘的研究集中在根、莖、葉和果實的化學成分以及所含化合物提取和藥用價值應用方面;其次是種子發芽處理和苗的栽培移植等方面[18];最后是關于桃金娘在我國地理區域分布和種子傳播等方面[13, 19]。【本研究切入點】植物光合作用強弱與環境因子有極大關系,環境因子隨著季節變化而改變,植物為了適應環境生理生化也會做出相應變化。【擬解決的關鍵問題】本試驗以3年生桃金娘苗為試驗材料,研究在不同季節下光合日變化與環境因子的關系以及不同季節葉綠素熒光參數變化情況,為以后桃金娘在生理生化研究方面奠定基礎,對桃金娘推廣種植和移植栽培提供理論指導。
1 材料與方法
1.1 試驗地概況
廣東省湛江市麻章區廣東海洋大學(湖光校區)位于110°17′ E,21°9′ N,處于北回歸線以南,屬于熱帶和亞熱帶地區,海拔25 m,離海近,全年受到海洋氣候的調節,夏天炎熱多雨,冬無嚴寒,降雨量少,年平均降雨量1400~1700 mm,年平均氣溫23 ℃,土壤偏酸性,多為紅壤、磚紅壤土等,適合熱帶、亞熱帶植物生長,多數為試驗用地,植物大多數為農作物和試驗樹種,周圍植物生長豐富。
1.2 試驗材料
2020年10月從廣州苗圃公司購買3年生桃金娘苗100株,種植在廣東省湛江市廣東海洋大學(湖光校區)種苗實驗實習基地,每株間隔50 cm,2021年1月存活95株,不定期除草施肥澆水。
1.3 試驗方法
1.3.1 光合指標測量 于春季(4月)、夏季(7月)、秋季(10月)、冬季(1月)之中選取連續晴天的3 d測量,測量時間為7:00—18:00,每隔1 h測量1次,使用便攜式光合儀(LCi-SD)測量,選取桃金娘頂端向下展開第2~3片葉片,共選取10株,每株1片葉子,每片葉子測5次,取總平均值。測定光合指標:有效光合輻射(PAR)、葉片溫度(Ti)、空氣CO2濃度(Ca)、空氣相對濕度(RH)、凈光合速率(Pn)、蒸騰速率(Tr)、胞間CO2濃度(Ci)、氣孔導度(Gs)、葉片水分利用率[(WUE)=Pn/Tr]、葉片光能利用率[(LUE)=Pn/PAR]。
1.3.2 熒光參數測量 于春季、夏季、秋季、冬季各選3 d,每天9:00 —11:30,使用葉綠素熒光氟筆(FP110 PAR-Fluor Pen)測量桃金娘熒光參數,同樣是由頂端向下選取第2~3片葉子,共10株,使用儀器配帶的夾子,夾住葉片進行20 min暗處理,然后進行各項熒光參數指標的測定。
非化學淬滅(NPQ)協議曝光和暗恢復階段持續時間、脈沖數量、間隔時間見表1。
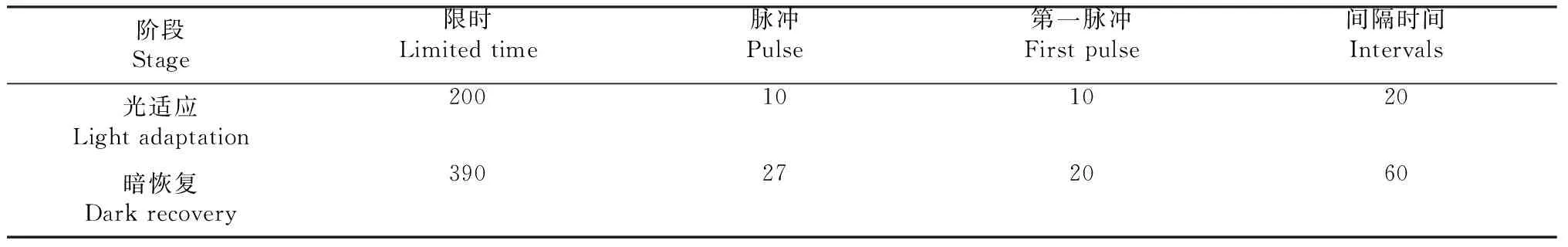
表1 曝光和暗恢復階段的持續時間以及脈沖之間的數量和間隔
OJIP協議:最小熒光(Fo)、最大熒光(Fm)、可變熒光(Fv)、PSII潛在活性(Fm/Fo)、PSII光能轉化率(Fv/Fm)、單位反應中心吸收的光能(ABS/RC)、單位反應中心捕獲的用于還原Qa的能量(TR0/RC)、單位反應中心捕獲的用于電子傳遞的能量(Eto/RC)、單位反應中心耗散掉的能量(Dio/RC)。
非化學淬滅(NPQ)協議測定熒光參數:光穩態下化學淬滅(QP-Lss)、光穩定下非化學淬滅(NPQ-Lss)、光稱定下量子產率(QY-Lss)最大量子產率(QY-max)和Kautsky誘導峰值。Kautsky誘導峰值:樣本暗處理使其光系統完全開放后,將樣品暴露在光照下持續幾十到幾百秒后引出一種叫做Kautsky效應的舜態。
光曲線(Lc)協議:光照強度設置10、20、50、100、300、500、1000 μmol/(m2·s)等7個梯度,測量熒光參數:最大熒光(Fm)、可變熒光(Fv)、量子產率(QY)。
1.4 試驗數據處理
試驗測量數據使用Excel 進行整理和作圖,使用SPSS 22.0軟件對數進行相關分析和多元線性回歸分析。
2 結果與分析
2.1 桃金娘不同季節生態因子日變化
由圖1可知,日均有效光合輻射(PAR)春季[1133 μmol/(m2·s)]>夏季[1121 μmol/(m2·s)]>秋季[997 μmol/(m2·s)]>冬季[982 μmol/(m2·s)];葉片溫度(Ti)春季和夏季最高溫度達40 ℃以上,日均溫春季(38.3 ℃)>夏季(38.1 ℃)>秋季(34.7 ℃)>冬季(31.9 ℃);冬季空氣CO2濃度(Ca)高于其它3個季節,Ca日均值冬季(432 mmol/mol)>秋季(394 mmol/mol)>春季(384 mmol/mol)>夏季(380 mmol/mol);空氣相對濕度(HR)日變化總體均呈現先下降后上升趨勢。
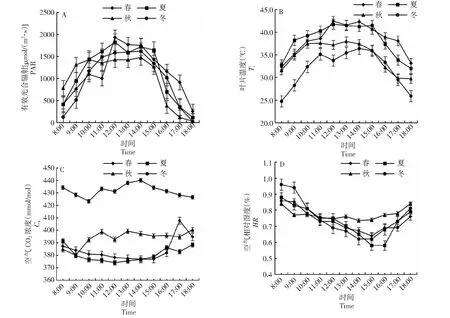
圖1 桃金娘不同月份下生態因子日變化Fig.1 Diurnal variation of ecological factors in different months of R. tomentosa
2.2 桃金娘不同季節凈光合速率(Pn)與生理因子日變化
由圖2-A可知,夏季桃金娘蒸騰速率(Tr)日變化呈“雙峰型”,春季、秋季、冬季桃金娘Tr日變化呈“單峰型”;夏季Tr峰值[3.77 mmol/(m2·s)]、日均值[2.79 mmol/(m2·s)],高于其它3個季節。由圖2-B可知,秋季Pn日變化呈“單峰型”,春季、夏季和冬季Pn日變化均呈“雙峰型”;春季在14:00時出現“光合午休”,夏季在13:00時出現“光合午休”,冬季在15:00時出現“光合午休”;日均Pn:秋季[7.74 mmol/(m2·s)]>夏季[5.91 mmol/(m2·s)]>冬季[4.97 mmol/(m2·s)]>春季[4.09 mmol/(m2·s)],Pn峰值:秋季[11.8 mmol/(m2·s)]>冬季[9.18 mmol/(m2·s)]>夏季[7.11 mmol/(m2·s)]>春[6.21 mmol/(m2·s)]。由圖2-C可知,夏季、秋季和冬季桃金娘胞間CO2濃度(Ci)日變化呈“U型”,早上和傍晚高,中午低,春季桃金娘Ci日變化呈“W型”;Ci日均值冬季[287 mmol/(m2·s)]>秋季[268 mmol/(m2·s)]>夏季[267 mmol/(m2·s)]>春季[197 mmol/(m2·s)]。由圖2-D可知,桃金娘夏季和秋季氣孔導度(Gs)日變化均呈現先下降后上升再下降趨勢,冬季Gs日變化呈“M型”;氣孔導度日均值:夏季[0.17 mmol/(m2·s)]>秋季[0.14 mmol/(m2·s)]>冬季[0.09 mmol/(m2·s)]>春季[0.05 mmol/(m2·s)]。
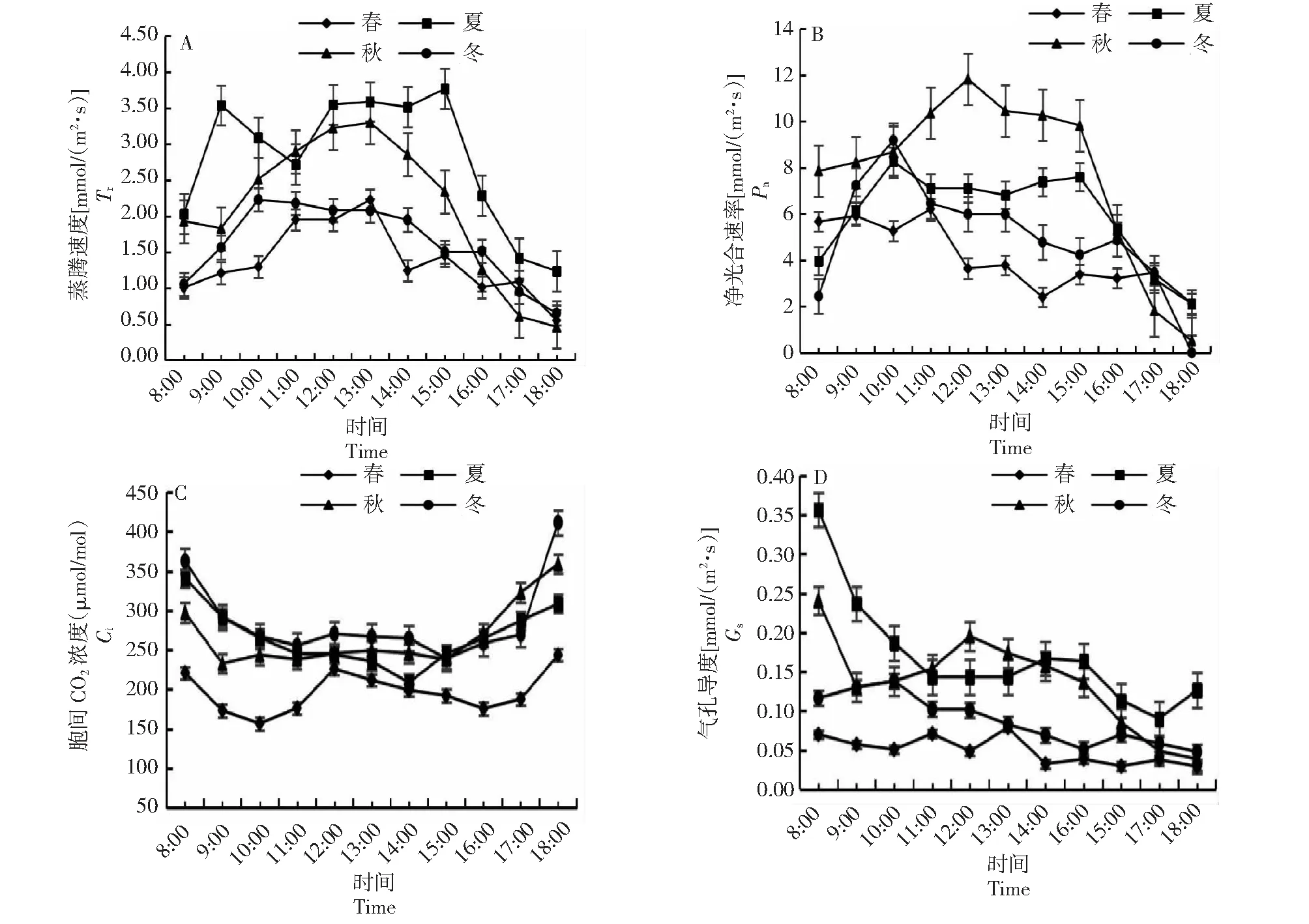
圖2 桃金娘不同月份凈光合速率,蒸騰速率、胞間CO2濃度、氣孔導度日變化Fig.2 Diurnal changes of net photosynthetic rate, transpiration rate, intercellular CO2 concentration and stomatal conductance of R.tomentosa in different months
2.3 桃金娘不同季節水分利用率(WUE)和光能利用率日變化(LUE)
由圖3-A可知,春季、夏季和冬季桃金娘對光能利用率(LUE)日變化總體均呈“U型”,秋季桃金娘LUE日變化呈“W型”;秋季日均LUE(0.0097 mmol/mol)最高,春季日均LUE(0.005 mmol/mol)最低。從圖3-B可知,春季和冬季水分利用率(WUE)日變化呈“U型”,夏季和秋季WUE日變化呈“M型”;日均WUE:秋季(3.51 mmol/mol)>春季(3.26 mmol/mol)>冬季(2.90 mmol/mol)>夏季2.12 mmol/mol。
2.4 桃金娘不同季節凈光合速率(Pn)與生理生態因子相關分析
從表2可知,桃金娘春季凈光合速率(Pn)與HR、Gs呈正相關;夏季凈光合速率(Pn)與PAR、Ti、Tr呈極顯著正相關,與Ca呈極顯著負相關,與Ci呈負相關;秋季凈光合速率(Pn)與PAR、Ti、Tr、Gs呈極顯著正相關,與Ci呈極顯著負相關,與HR呈負相關;冬季凈光合速率(Pn)與Tr呈極顯著正相關,與PAR、Gs呈正相關,與Ci呈負相關。
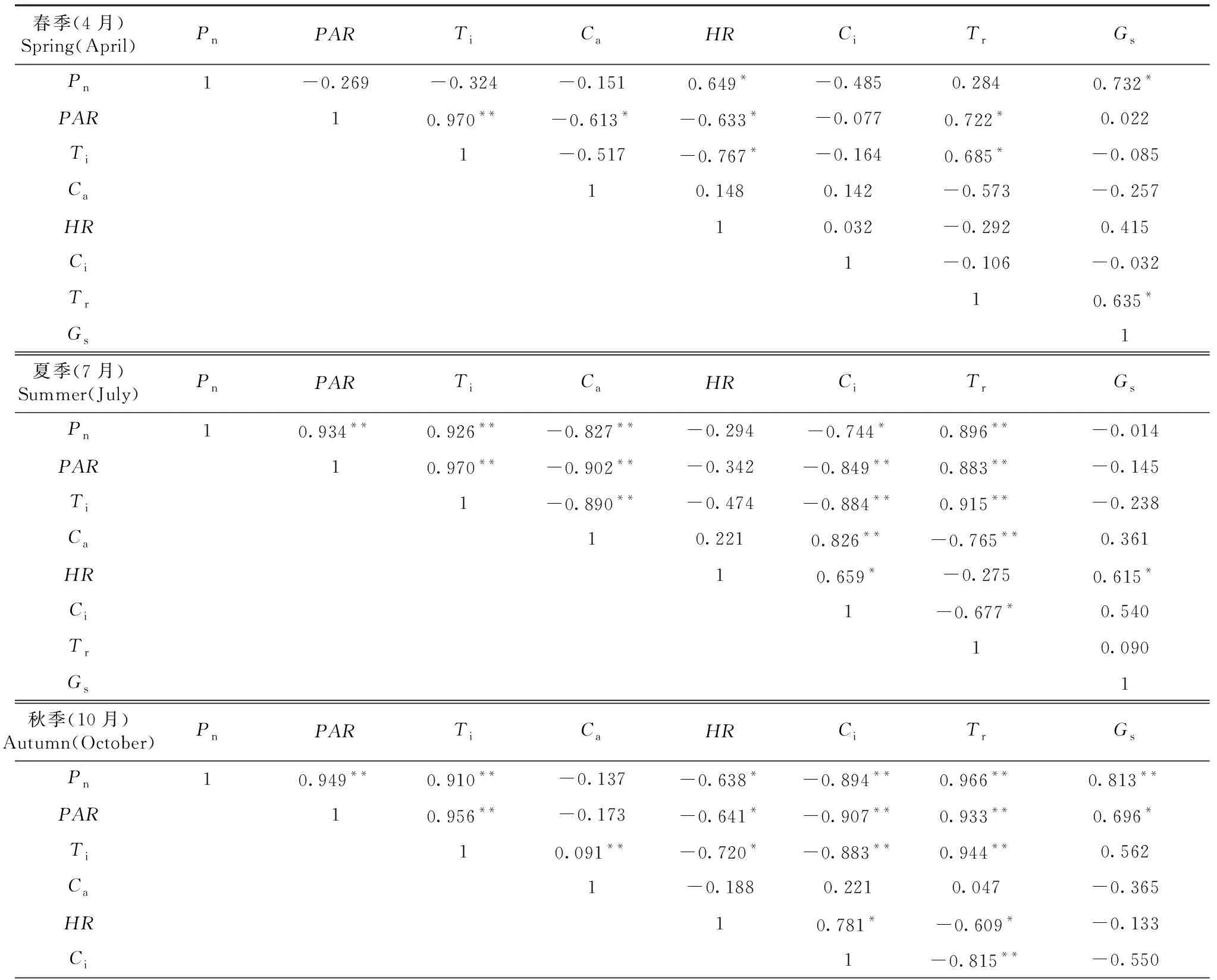
表2 桃金娘春季、夏季、秋季、冬季凈光合速率(Pn)與生理生態因子相關矩陣
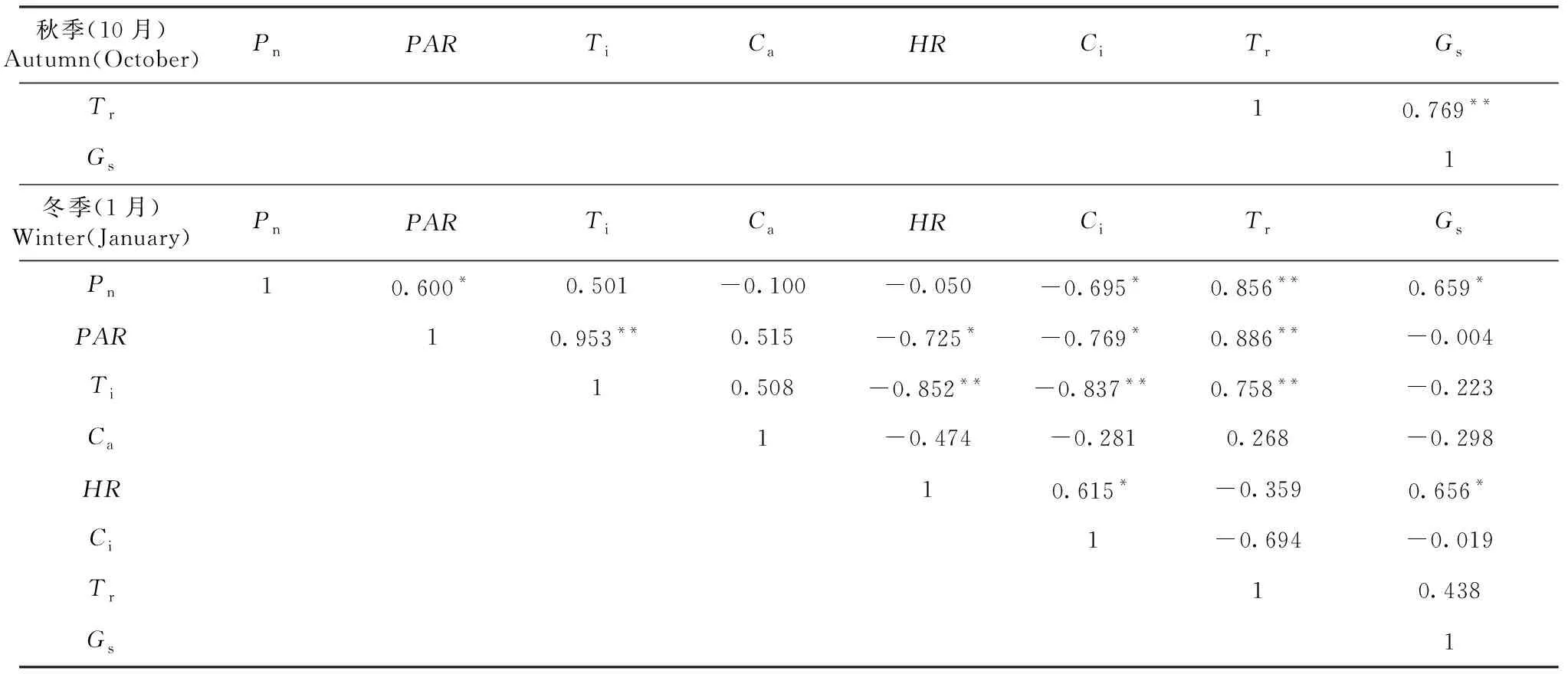
續表2 Continued table 2
對不同季節桃金娘Pn進行正態分布檢測采用小樣本Shapiro-wilk檢測(春季sig.=0.229,夏季sig.=0.153,秋季sig.=0.051,冬季sig.=0.974),檢測結果值均大于0.05符合正態分布,可以進行線性回歸方程分析。對不同季節桃金娘Pn(Y)日變化與生理生態影響因子(XCi、XGs、XCa、XTi、XPAR、XTr)日變化進行逐步多元線性回歸方程分析。得到不同季節下桃金娘最優多元線性回歸方程(表3):Gs、HR、Ci是影響春季桃金娘Pn的主要因子,PAR是夏季凈光合速率主要影響因子,Tr、Gs、Ci是秋季凈光合速率主要影響因子,Tr、Ca是冬季凈光合速率主要影響因子。
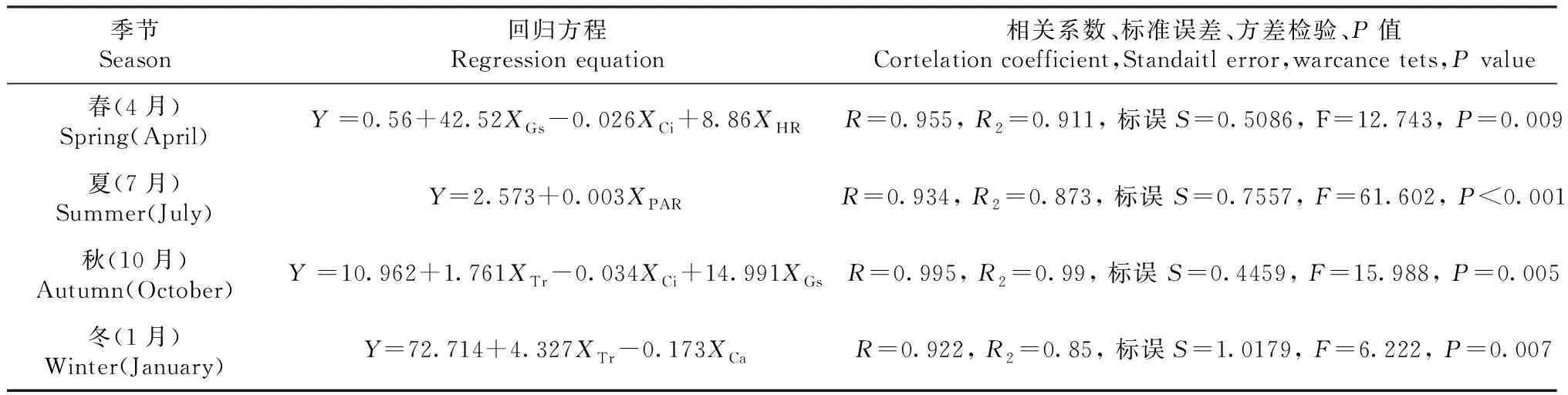
表3 不同季節桃金娘多元線性回歸方程
2.5 不同季節桃金娘光系統PSII活性參數
2.5.1 桃金娘不同季節Fo、Fm、Fv值 從圖4~5可知,桃金娘夏季的Fo、Fm、Fv值最大,冬季的值最小;夏季的光系統PSII的潛在活性(Fm/Fo)和PSII光能實際轉化率(Fv/Fm)最高,其數值分別為2.76、0.64,春季和秋季居中,冬季桃金娘的光系統PSII的熒光數值最低。
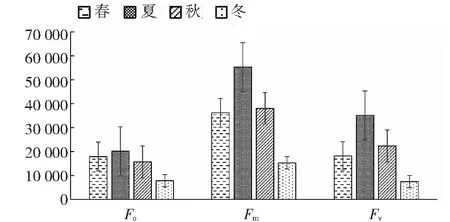
圖4 不同季節桃金娘的Fo、Fm、Fv值Fig.4 The Fo, Fm and Fv values of R. tomentosa in different seasons
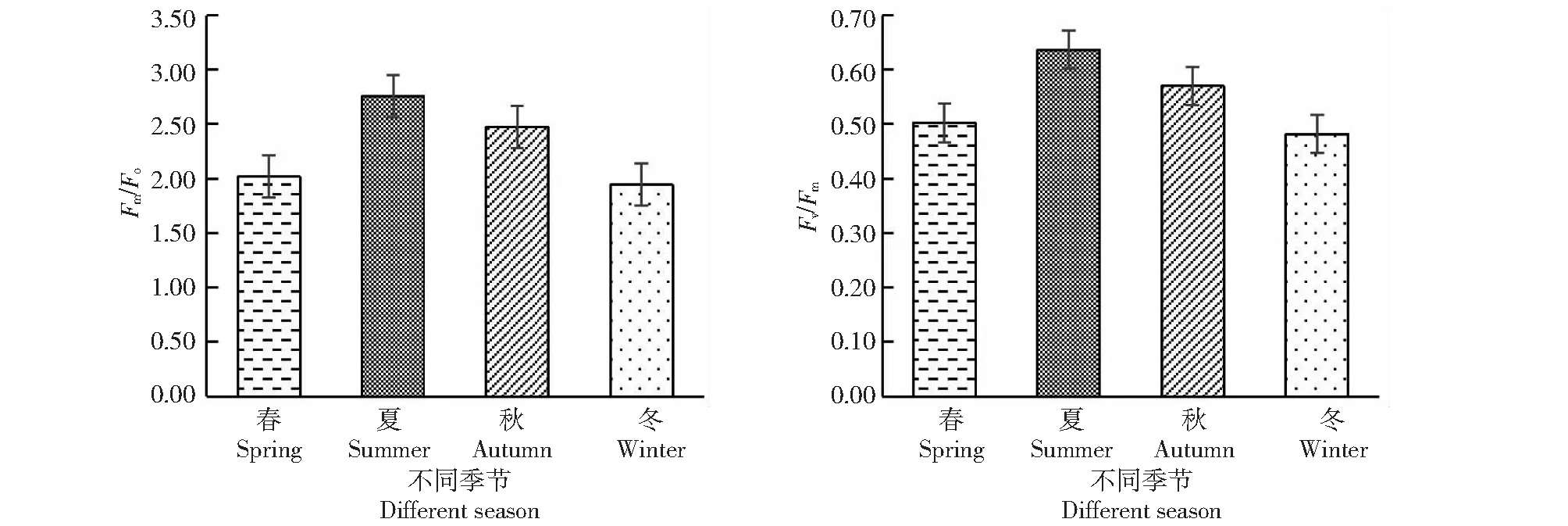
圖5 不同季節桃金娘光系統PSII的Fm/Fo、Fv/Fm值Fig.5 Fm/Fo and Fv/Fm values of PSII of R.tomentosa optical system in different seasons
2.5.2 不同季節桃金娘光系統中單位反應中心能量捕獲、利用以及消散情況 從圖6可知,桃金娘光系統中反應單位捕獲能量多少、利用以及消耗等情況,春季桃金娘的單位反應中心吸收(ABS/RC)的能力最大,冬季最小;夏季用于還原質體醌a的能量(TR0/RC)最大,冬季最小;秋季用于電子傳遞的能量(Eto/RC)最多,春季最小;春季單位反映中心耗散掉的能量(Dio/RC)最多,夏季最小。
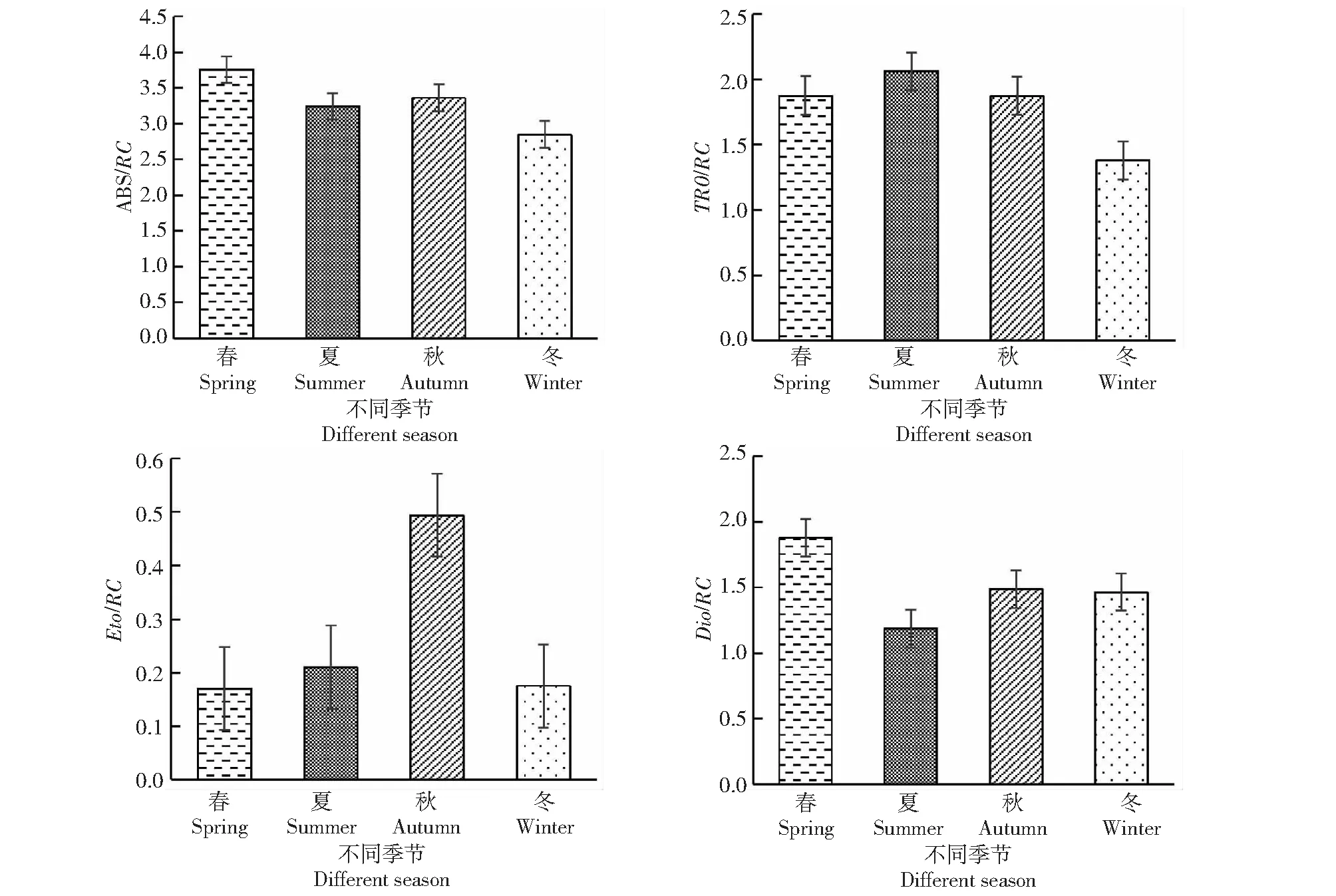
圖6 不同季節桃金娘單位反應中心能量吸收、分配以及消耗情況Fig.6 Energy absorption, distribution and consumption of R. tomentosa unit reaction center in different seasons
2.5.3 桃金娘不同季節下的化學淬滅(Qp-Lss)與非化學淬滅(NPQ-Lss) 從圖7-A~7-B可知,夏季桃金娘的Kuatsky值最大,其次是秋季和春季、冬季最小。在光穩態條件下,夏季的Fm最大,冬季最小;春季、冬季的化學淬滅(QP_Lss)高于夏季、秋季,夏季、秋季的非化學淬滅(NPQ_Lss)高于春季和冬季,夏季、冬季的量子產率(QY_Lss)高于春季和秋季。暗處里后的量子產率(QY_max)結果是夏季=冬季>秋季>春季。
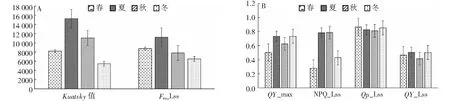
圖7 不同季節下桃金娘Kuatsky值、暗處里下QY和光穩態下QP、NPQ、QY值Fig.7 R.tomentosa Kuatsky value, QY in the dark and QP, NPQ, QY in light steady state in different seasons
2.5.4 不同光照強度下桃金娘的量子產率(QY)、光系統PSII活性(Fv/Fm) 從圖8-A~8-B可知,春、夏、秋3季量子產率(QY)隨著光照強度增加呈下降趨勢,冬季的量子產率(QY)隨著光照強度增加呈先上升后下降趨勢,光照強度為50 μmol/(m2·s)時冬季桃金娘的QY最大;隨著光照強度增加,桃金娘春、夏、秋、冬四季的PSII光能轉化率(Fv/Fm)總體呈上升趨勢,光照強度大于100 μmol/(m2·s)時PSII光能轉化率(Fv/Fm)上升相對較快。
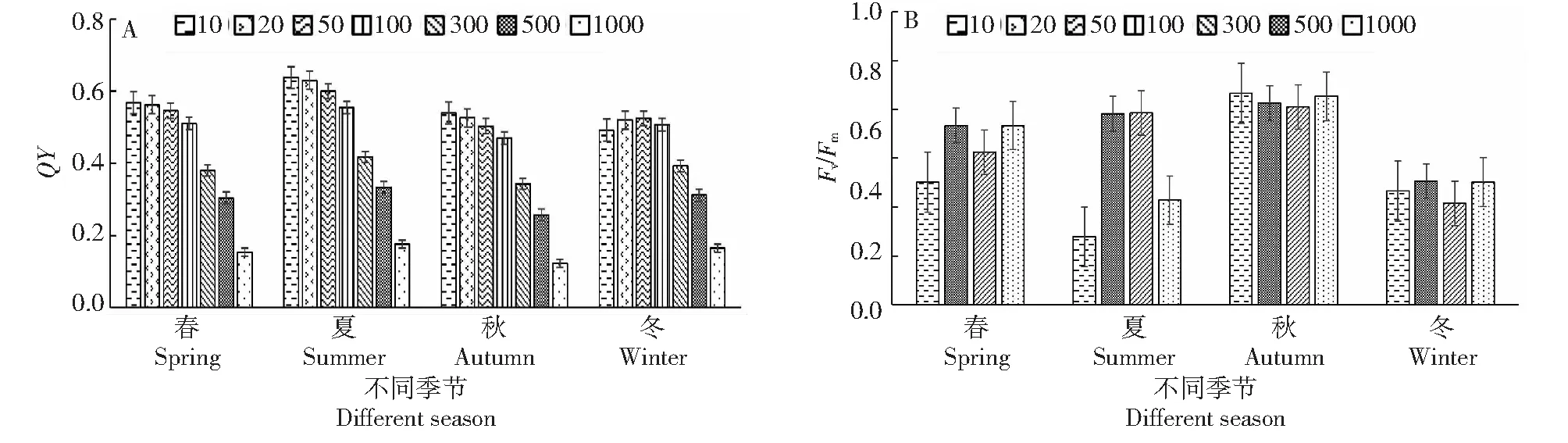
圖8 不同光照強度下桃金娘的QY、Fv/Fm值Fig.8 QY,Fv/Fm values of R.tomentosa under different light intensities
3 討 論
光合作用是植物生長發育的重要代謝途徑,凈光合速率是衡量植物光合作用的一個重要指標[20],凈光合速率受外界生態因子和植物自身生理因子的影響,生理生態因子的變化與季節變化具有很大關系[21]。本試驗研究顯示:桃金娘冬季凈光合速率與蒸騰速率呈極顯著正相關,春季凈光合速率與生理生態因子沒有存在極顯著的相關性,這一研究結果與劉旻霞等[22]對黃土高原中部3種典型綠化植物光合特性的季節變化研究結果一致。多元線性回歸分析能夠在多個因子中找出對因變量影響最大的因子[23],桃金娘凈光合速率與生理生態因子做線性回歸分析,找出影響凈光合速率的主要因子,本研究結果顯示,春季影響凈光合速率(Pn)的主要因子有Gs、HR、Ci, 夏季則是PAR 為主要因子,Tr、Gs、Ci是影響秋季Pn主要因子,春季影響Pn的主要因子是Tr、Ca, 隨著季節變化對桃金娘Pn影響的主要因子也隨著改變。
植物的光能利用率(LUE)和水分利用率(WUE)在一定程度上可以反應植物在不同環境下資源配置,也是一種抗逆性的重要參考指標[24]。研究結果顯示,桃金娘在不同季節對水分和光能利用不同,秋季WUE 最高,夏季WUE 最小,可能與湛江氣候變化有關;秋季LUE 最高,同時光能利用率時間相對與其它3季較長。
植物光合日變化出現“光合午休”是植物適應環境的一個重要策略[25-26],許大全等[27]對植物“光合午休”生理特征研究表明,Pn下降、Gs下降、Ci下降為氣孔限制所引起的“光合午休”;Pn下降、Gs下降、Ci上升為非氣孔限制引起的“光合午休”。本研究結果顯示:桃金娘春季、夏季、冬季Pn日變化都存在“光合午休”,春季Pn下降、Gs下降、Ci下降,表明春季“光合午休”是由氣孔限制所引起的;夏季Pn下降、Gs下降、Ci上升,表明夏季“光合午休”由非氣孔限制引起;冬季Pn下降、Gs下降、Ci下降,表明冬季“光合午休”也是由氣孔限制引起,不同季節引起桃金娘“光合午休”類型各不相同。
植物熒光與光合作用密切相關,能夠反應葉片葉綠體內部一些機構情況[4],暗適應后Fo指PSII反應中心完全開放的是熒光,Fo越大表明PSII反應中心遭到的破壞越嚴重[28-29];植物PSII反應中心完全關閉時熒光水平最大,受到光抑制時Fm值下降。Fm/Fo表示光系統PSII潛在活性[30]。本研究顯示,夏季的Fo、Fv、Fm最大,冬季Fo、Fv、Fm最小,可能是夏季生態因子中有效光合輻射相對較高造成Fo升高,有效光合輻射升高使葉片溫度升高,進而提升光系統PSII中相關酶活性升高,夏季光系統PSII潛在活性(Fm/Fo)高于其它3季,光系統PSII實際轉效率(Fv/Fm)是夏季>秋季>春季>冬季,桃金娘夏季最大熒光、可變熒光增大可能有利于增加光能的存儲量以及提高利用率。
植物光合作用吸收的光能要轉變成穩定的化學能主要經過光系統PSII等一系列電子傳遞,把光能轉化為不穩定的化學能ATP,然后在經過碳同化形成穩定有機物[31]。在整個光系統PSII過程中質體醌周轉次數、質體醌庫大小、質體醌還原速率等都會影響光合作用[32]。本研究結果顯示,春季ABS/RC最多,同時Dio/RC也最大;夏季對TRO/RC利用使最高,消散掉的能量最少;秋季Eto/RC最大,不同季節下桃金娘光系統PSII對捕獲能量分配的策略也各有不同。
當大量過剩光能不能及時用于光化學反應時, 就能形成光抑制現象, 植物為了避免PSII受到破壞, 往往通過葉黃素循環、熒光發射、光呼吸等熱耗散途徑來消耗過剩光能[33]。Kautsky峰值指將植物葉片暗適應后突然暴露在光照下,葉綠體分子吸收能量后由低能量的穩態轉變為高能量不穩定的激發態,激發態的電子不穩定壽命非常短很快又回到穩態中去,在此過程中小部分激發態能量以紅色的熒光形式消散掉的值[7]。本研究結果顯示,突然暴露在光照條件下,桃金娘Kautsky峰值:夏季>秋季>春季>冬季,葉綠色素捕獲的光能以紅色的熒光形式消散掉的能量損耗:夏季>秋季>春季>冬季。
從植物捕獲光能分配上來看,用于植物光合作用稱為化學淬滅(Qp),沒有用于光合作用而是以熱量等形式消散掉稱為非化學淬滅(NPQ)[34]。化學淬滅(Qp)越高植物對捕獲光能利用率也就越高;非化學淬滅(NPQ)是植物為保護光系統,會把捕獲過多的能量以熱能的形式消耗掉。本試驗研究結果顯示,在光穩態下桃金娘Qp為春季>冬季>夏季>秋季,NPQ則為秋季>夏季>冬季>春季,QY為夏季>冬季>春季>秋季;桃金娘經過暗處理后的QY為夏季=冬季>秋季>春季。
植物的光響應曲線對了解植物光化學過程具有重要意義[35]。當植物光系統吸收的光能超過植物被利用能量極限時,植物光合作用會受到抑制,Fv/Fm兩者變化是判斷植物是否發生光抑制的重要指標[36-37]。本試驗研究顯示,桃金娘春季和冬季在光照強度為100 μmol/(m2·s)時,QY達到最大值,超過100 μmol/(m2·s)后QY都會隨著光照強度增加而減小,夏季和秋季桃金娘QY隨著光照強度增加而減小;春季、夏季、秋季的光系統PSII活性(Fv/Fm)的變化并未出現隨著光照強度增強而下降的現象,冬季桃金娘在光照強度小于50 μmol/(m2·s)時,光系統PSII活性(Fv/Fm)隨著光照強度增加呈遞減趨勢,當光照強度超過50 μmol/(m2·s)后,隨著光照強度增加而增加。這一研究與王曉燕[38]的研究結果不一致。根據眾多研究發現高等植物在非脅迫下Fv/Fm穩定在0.83左右[39],桃金娘在光照強度小于500 μmol/(m2·s)時,Fv/Fm<0.83,說明其是否受到光抑制還有待進一步研究。
4 結 論
桃金娘在春季和冬季的“光合午休”由氣孔限制引起,夏季桃金娘的“光合午休”屬于非氣孔限制;不同季節下生理生態因子對光合作用的影響大小不同,桃金娘春季Pn受到Gs、HR以及Ci影響,夏季Pn受到PAR影響,秋季Pn受到Tr、Gs、Ci影響,冬季Pn受到Tr、Ca影響;桃金娘不同季節下光系統PSII活性(Fv/Fm)為夏季>秋季>春季>冬季,ABS/RC為春季>秋季>夏季>冬季,光穩態下QY為夏季>冬季>春季>秋季,最大QY為夏季>冬季>秋季>春季;春季和冬季桃金娘QY在光照強度100 μmol/(m2·s)時最大,夏季和秋季桃金娘QY隨著光照強度增加而減小。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
