外源鈣緩解花生低溫光合障礙的調控機制
2022-03-16 08:36:12劉欣悅劉軼飛易伯濤孫志宇張思威馬明珠韓曉日
植物營養與肥料學報 2022年2期
劉欣悅,劉軼飛,易伯濤,孫志宇,張思威,馬明珠,韓曉日
(土肥高效利用國家工程研究中心/沈陽農業大學土地與環境學院,遼寧沈陽 110161)
起源于南美洲熱帶地區的喜溫作物—花生(Arachis hypogaea)是世界重要的油料和經濟作物,也是重要的食用蛋白源和食用植物油源,在保障我國食用油安全方面具有舉足輕重的作用[1]。特別是,基于后疫情時代“雙循環”新發展格局下、中國大豆產需缺口加大等背景下,中央明確了油料作物結構調整主要策略之一,就是多措并舉發展國內花生生產替代大豆進口。因此,我國油料作物供給(食用油)安全和國際新形勢新挑戰已然將花生的優化施肥技術與耐冷優質高效生產推到一個前所未有的國家戰略安全高度[2]。
遼寧省是我國重要的優質花生出口基地和產區之一,花生在遼寧省大田作物中其種植面積僅次于玉米和水稻。遼寧省土壤中黃曲霉菌含量極低,遼寧產區的花生(黃曲霉毒素顯著低于其他產區)完全符合出口創匯的優級標準,這一優勢也使得“遼寧花生”在全國乃至全球花生產業中品牌效應強,不可替代。低溫是限制花生地理分布和生產的主要環境因子之一,冷害在全球花生生產中普遍存在[3],在我國尤其以北方高緯度區域、南方高海拔地區為甚。研究表明,大多數花生在幼苗期環境溫度低于
12℃~16℃時,植株生長緩慢,嚴重時停止生長,葉片會出現脫水、萎蔫甚至枯死等現象[4–5]。前人大量研究表明,低溫導致植物光合障礙,生長發育停滯,甚至死亡[6];低溫脅迫對植物生長發育和光合碳同化過程產生負面影響[7]。研究發現,低溫脅迫顯著降低了水稻、玉米、向日葵、高粱、蘋果幼苗的葉面積[8–9],抑制了玉米根系生長和干物質積累等[10]。低溫脅迫可破壞光合作用的主要機構,包括葉綠體的結構、類囊體膜功能、光合電子傳遞和氣孔運動等[11–12],降低植物光合作用水平,進而導致生物量降低、葉片壞死等問題[13]。
鈣素作為一種植物必需的礦質養分,在維持細胞壁和細胞膜的結構完整性和功能穩定性上具有多重作用,同時鈣離子作為信號物質在調節植物應對非生物脅迫中也發揮重要功能[14]。雖然土壤中含有大量鈣素,但是作物在低溫逆境下根系活力和葉片蒸騰速率驟降,時常發生嚴重吸鈣障礙[15]。為此,葉部抗寒增鈣技術作為植物應對(寒)冷逆境的外源調控措施,現已在水稻、玉米、番茄、蘋果、黃瓜、煙草等作物上廣泛應用[16–18]。外源鈣預處理有助于維持細胞膜的完整性和抗氧化系統的平衡,從而減輕膜脂的過氧化反應[19–20]。課題組前期研究發現,葉面施鈣具有顯著提升花生耐冷性的調控效應[12–13],但是外源鈣素的調控作用是否與解除低溫光合障礙有關,這有待于進一步探究。為此,本試驗應用人工氣候室盆栽培養花生苗,同時采用外源鈣離子(鈣素正向調控)及其鈣抑制劑EGTA (鈣素負向調控)對花生苗進行預處理,從正反兩方面深入探討外源鈣素對花生低溫依賴性光合反饋抑制的調控作用,這對于最終實現花生生產中科學施用鈣肥,以及進一步開發寒區花生高產優質施肥配套技術等具有重要指導意義。
1 材料與方法
1.1 試驗設計
試驗于2019年在沈陽農業大學土地與環境學院人工氣候室內進行,人工氣候室光合光子通量密度(PPFD)為 600 μmol/(m2·s),CO2濃度為 (400 ± 5) μmol/mol,相對濕度為 (60 ± 5)%。
試驗以東北主栽花生品種“小白沙”為材料,利用48孔穴盤進行花生育苗,待長出第一片真葉時選取100株長勢均勻一致的花生苗移栽至直徑為13 cm 的圓柱盆中,平均分為 4個處理組:1) CK (夜溫/晝溫 20℃/28℃ + 葉面噴施超純水),2) +H2O (夜溫/晝溫 9℃/15℃ + 葉面噴施超純水),3) +Ca (夜溫/晝溫 9℃/15℃ + 葉面噴施 15 mmol/L CaCl2),4)+EGTA (夜溫/晝溫 9℃/15℃ + 葉面噴施 5 mmol/L EGTA)。待真葉展開一周后,連續兩天(下午3:00—4:00)分別噴施超純水、15 mmol/L CaCl2和 5 mmol/LEGTA處理,以上調控處理條件均為課題組前期篩選得出的最佳條件[4, 12–13]。于葉面噴施試劑處理后的第二天夜晚開始,模擬常見的花生苗期低溫冷害情況,低溫持續處理7天。于第7天的夜間恢復常溫20℃,繼續生長7天,整個試驗處理周期為14天。
1.2 測定方法
1.2.1 生長指標的測定方法各處理隨機選取生長均一的3株花生,于低溫處理第7天和恢復常溫后的第7天上午9:00—10:00測量生長指標。株高與莖粗分別利用直尺與游標卡尺測量,葉面積和相對葉綠素含量(SPAD值)分別利用LI-3000C葉面積儀和CCM-200 plus葉綠素儀測定。
1.2.2 光合及熒光參數的測定方法光合和熒光參數分別在低溫的第1、4、7天及其常溫恢復過程中的第1、4、7天的上午9:00開始測定,每株選取“倒三葉”作為指定葉片,各處理隨機選取3株作為重復。光合氣體交換參數采用GFS-3000 (WALZCo.,德國)測定,其中瞬時水分利用率(WUEt)以及氣孔限制值(Ls)參照以下公式計算:
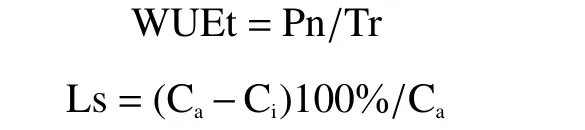
式中,Ca為大氣CO2濃度,Ci為胞間CO2濃度。
最大光化學效率Fv/Fm、PSI實際量子產量Y(I)、PSI電子傳遞速率ETR (I)、供體側限制引起的PSI處非光化學能量耗散的量子產量Y (ND)、受體側限制引起的PSI處非光化學能量耗散的量子產量Y (NA)、PSII實際量子產量 Y (II)、PSⅡ電子傳遞速率ETR (II)、PSII處非調節性能量耗散的量子產量Y(NO)與PSII處調節性能量耗散的量子產量Y (NPQ)利用雙通道葉綠素熒光儀Dual-PAM-100 (WALZCo.,德國)測定。
1.2.3 碳水化合物及活性氧含量的測定方法在低溫處理的第7天和恢復常溫的第7天取樣,測定葉片碳水化合物和活性氧含量。各處理隨機選取3株的倒三葉,105℃殺青30 min,65℃烘干,粉碎。葡萄糖、蔗糖、果糖、淀粉等含量測定參照薛應龍[21]的方法。丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法測定,過氧化氫(H2O2)含量的測定參照Jana等[22]方法并加以改進。超氧陰離子的測定方法按照王愛國等[23]的方法并加以改進。
1.3 數據分析
利用 Excel 2016制表,Origin 2021作圖。采用SPSS 19.0 軟件進行單因素方差分析和最小顯著差數法(LSD)檢驗,結果以3次生物學重復的平均值和標準誤差表示。
2 結果與分析
2.1 低溫脅迫及常溫恢復條件下外源鈣素對花生幼苗生長的影響
由表1可以看出,在低溫處理7天時,噴施H2O和EGTA處理株高顯著低于CK,而噴施Ca處理與CK無顯著差異;而在恢復常溫7天后,+H2O、+Ca和+EGTA 3個處理的株高之間無顯著差異,但均顯著低于CK。低溫脅迫下,噴施H2O處理莖粗最大,而恢復常溫后,施Ca處理莖粗最大,噴施EGTA處理最小。對照葉面積在低溫脅迫和恢復常溫后均最大,其他3個處理低溫下無明顯差異,但噴施Ca處理比施H2O和EGTA處理分別高42.9%和22.6%。低溫脅迫顯著降低了相對葉綠素含量SPAD,而恢復常溫后,除了噴施EGTA處理SPAD值顯著低于CK之外,其余處理與CK差異不顯著。低溫脅迫下,噴施Ca處理的根冠比雖然顯著高于CK,但顯著低于噴施H2O和EGTA處理;恢復常溫后,噴施EGTA處理的根冠比依然顯著高于CK。
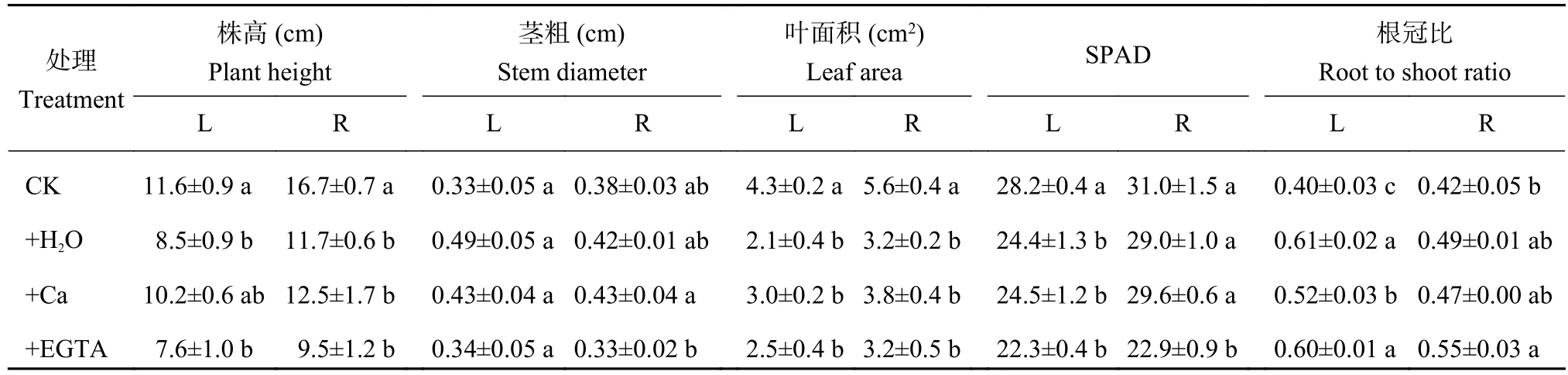
表1 低溫脅迫7天(L)及恢復常溫7天(R)時不同處理花生生長狀況Table 1 Peanut growth at 7 days of low temperature stress (L) and at 7 days after recovery to regular temperature (R)
2.2 低溫脅迫及常溫恢復條件下外源鈣素對花生葉片光合氣體交換水平的影響
從圖1可以看出,低溫處理及恢復常溫期間,凈光合速率(Pn)整體呈先下降后上升逐漸趨于平緩的趨勢,在低溫脅迫7天時,噴施Ca和施H2O處理Pn低于CK,但高于噴施EGTA處理;恢復常溫7天后,噴施Ca處理的Pn與CK無顯著差異。細胞間二氧化碳濃度(Ci)隨著低溫脅迫天數的增加,整體呈先增長后下降的趨勢。低溫脅迫下噴施EGTA處理的氣孔導度(Gs)最低,噴施Ca處理與CK差異最小,恢復常溫后,噴施H2O與噴施EGTA處理Gs有增加趨勢。蒸騰速率(Tr)隨低溫脅迫天數增加逐漸降低,恢復常溫后逐漸趨于平穩,噴施Ca處理Tr值與CK無顯著差異。瞬時水分利用率(WUEt)隨低溫脅迫天數增加逐漸降低,恢復常溫后逐漸趨于平穩,低溫脅迫下,以噴施Ca處理最大。低溫處理及恢復常溫期間,氣孔限制值(Ls)整體呈先下降后上升的趨勢,低溫脅迫1~4天快速下降,恢復常溫后上升趨勢明顯。綜上,隨著低溫脅迫時間的延長,花生葉片Pn、Gs、Tr也逐漸降低,Ci逐漸上升,恢復常溫7天后,噴施Ca處理的Pn、Tr、WUEt與CK無顯著差異。
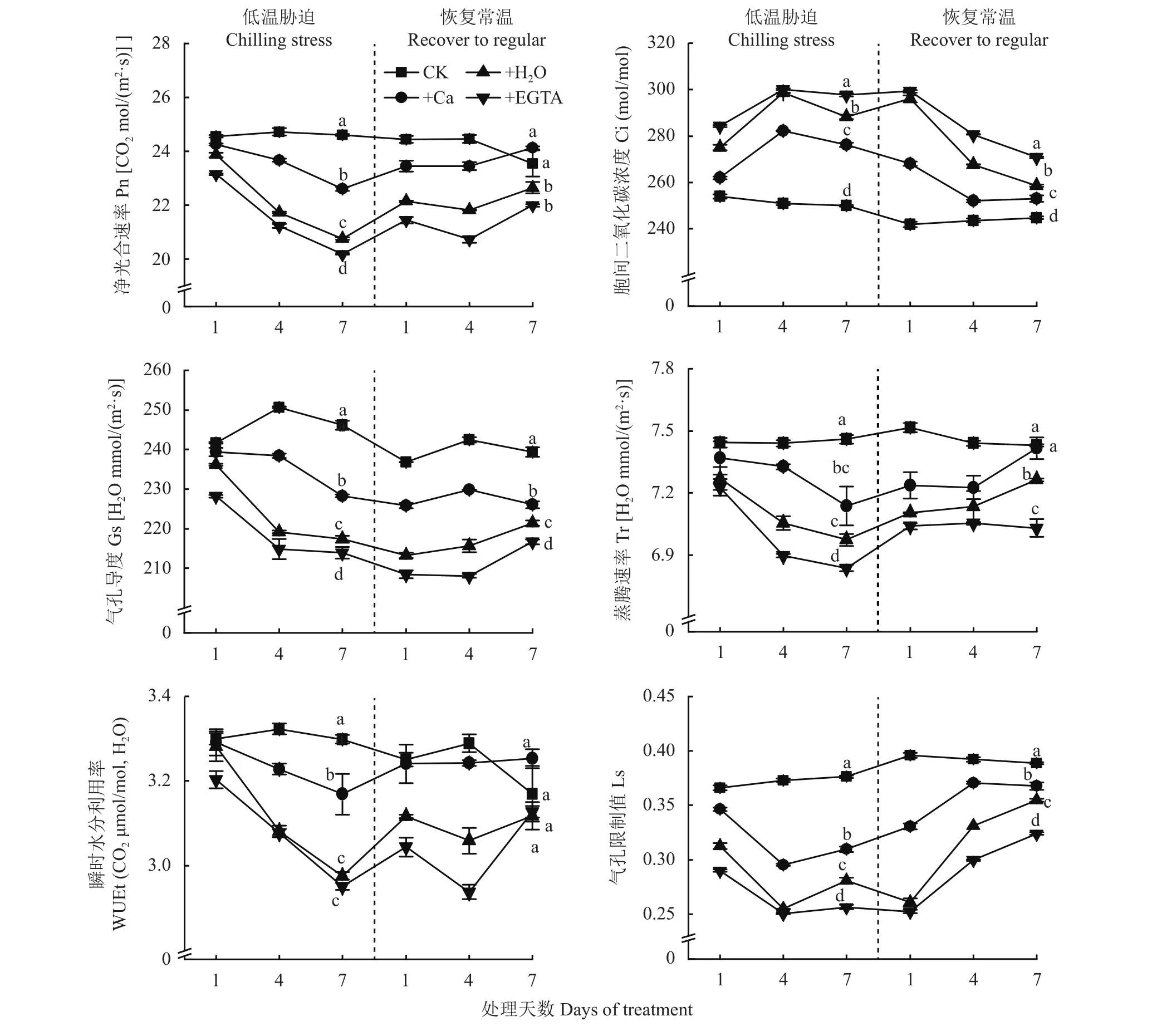
圖1 低溫脅迫及恢復常溫后各處理花生氣體交換參數Fig. 1 Leaf gas exchange parameters at different days of chilling stress and recovery to normal temperature under different treatments
2.3 低溫脅迫及常溫恢復條件下外源鈣素對花生最大光化學效率Fv/Fm的影響
圖2表明,PSII最大光化學效率Fv/Fm隨著低溫脅迫時間的延長呈現逐漸降低趨勢,3個低溫處理變化趨勢相似,隨低溫脅迫時間延長,噴施EGTA處理Fv/Fm下降速率最大,噴施H2O和噴施Ca處理Fv/Fm下降的趨勢與噴施EGTA處理相比較為緩慢;在低溫脅迫第7天,3個低溫處理與CK相比有顯著差異,其中施Ca處理Fv/Fm大于噴施H2O和EGTA處理;在恢復常溫的1~4天,除CK外的3個處理Fv/Fm呈快速上升的趨勢,噴施Ca處理在恢復常溫第7天Fv/Fm恢復到CK水平。
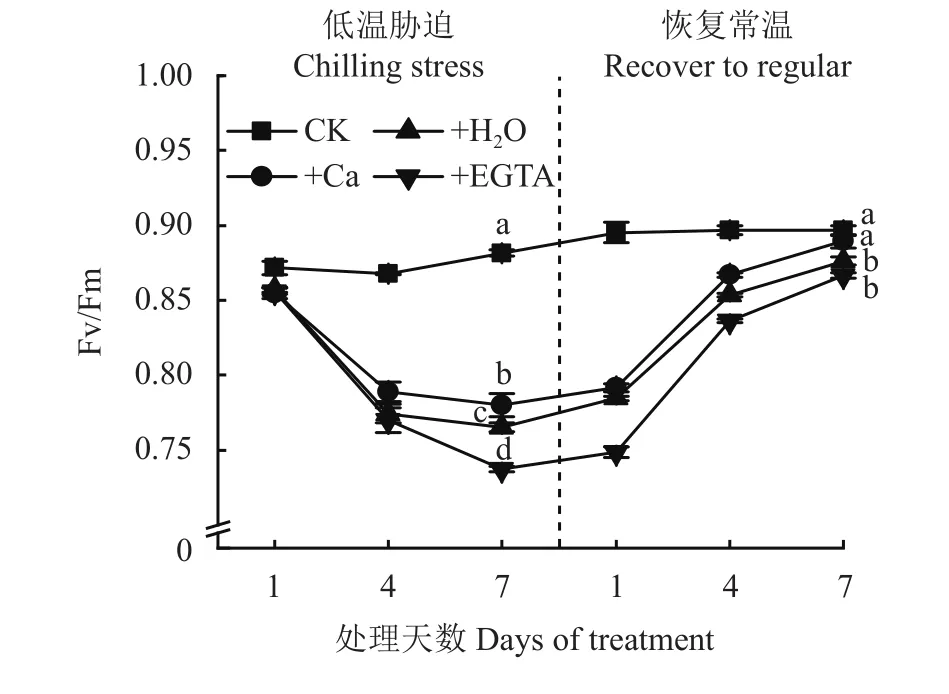
圖2 低溫脅迫及其常溫恢復條件下各處理花生葉片PSII最大光化學效率(Fv/Fm)Fig. 2 The maximum quantum yield of PSII (Fv/Fm) in peanut leaves at different days of chilling stress and recovery to normal temperature under different treatments
2.4 低溫脅迫及常溫恢復條件下外源鈣素對花生葉片PSI和PSII光化學活性的影響
由圖3可以看出,低溫脅迫1~7天時,+H2O、+Ca和+EGTA 3個處理的 Y (I)和 ETR (I)都顯著降低,但噴施Ca處理降低最少;恢復常溫后,+H2O、+Ca和+EGTA 3個處理的 Y (I)和 ETR (I)均呈上升趨勢,且在恢復常溫后的1~4天ETR (I)增加速率最大。在低溫脅迫1~7天時,Y (ND)和Y (NA)呈上升的趨勢,其中施Ca處理的Y (ND)最大,CK最小。Y(II)和ETR (II)總體趨勢與Y(I)和ETR(I)相似,其中在1~4天的低溫脅迫期間ETR(II)下降速率最大,Y(II)在恢復常溫第7天時達到CK水平。隨著低溫脅迫時間延長,Y (NO)顯著上升,噴施H2O和EGTA處理的Y (NPQ)與Y (NO)高于施Ca處理,低溫條件下的+H2O、+Ca和+EGTA 3個處理間差異顯著。
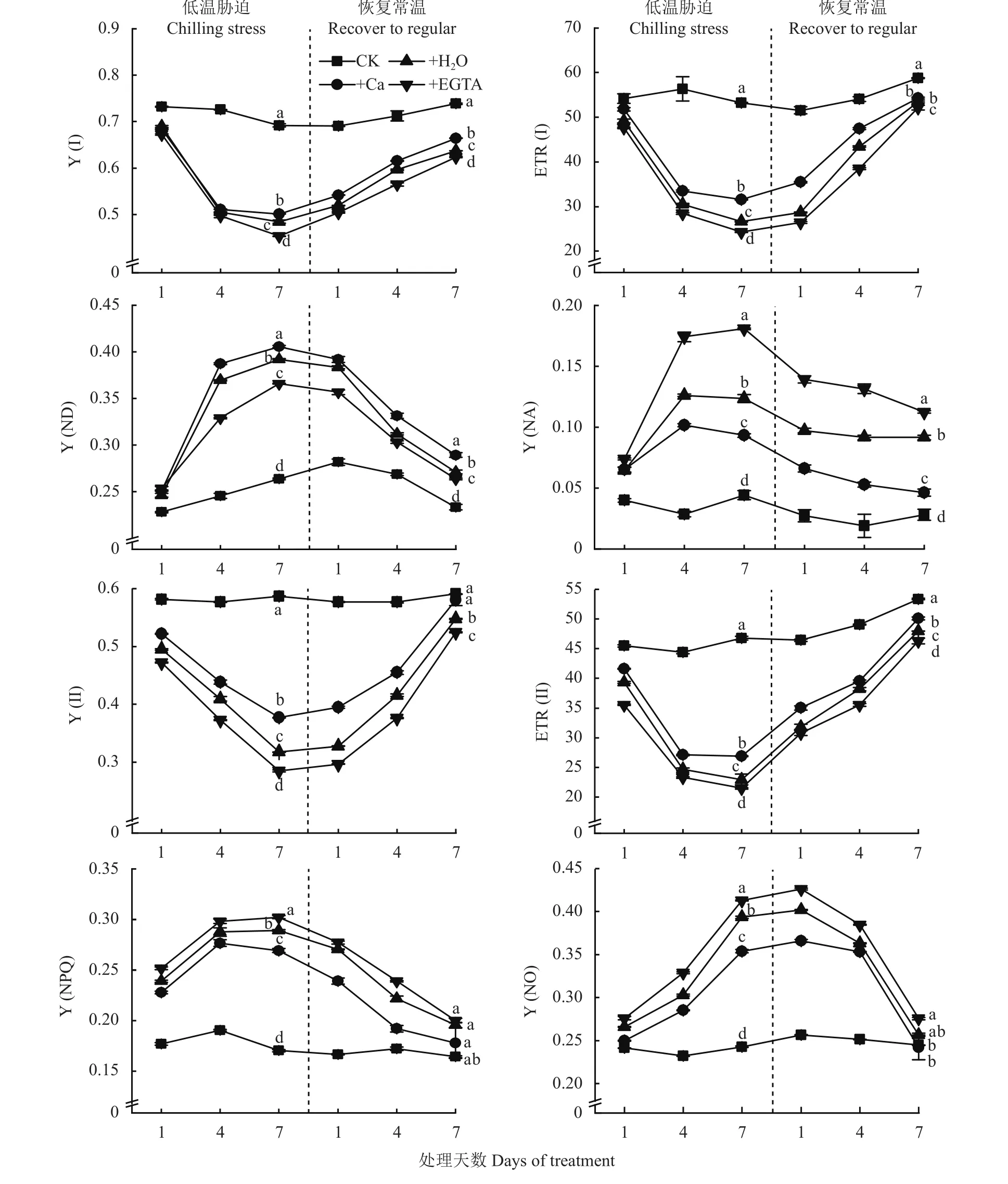
圖3 低溫脅迫及其常溫恢復條件下各處理花生葉片PSI和PSII光化學活性Fig. 3 PSI and PSII photochemical activities in peanut leaves at different days of chilling stress and recovery to normal temperature under different treatments
2.5 低溫脅迫及常溫恢復條件下外源鈣素對花生葉片非結構性碳水化合物積累的影響
由圖4可以看出,在低溫處理7天時,葡萄糖含量+Ca處理顯著高于其他處理,+H2O和+EGTA處理顯著低于對照;在常溫恢復7天,+H2O、+Ca和+EGTA顯著低于對照。與CK相比,3個低溫處理的蔗糖含量顯著增加。在恢復常溫后,蔗糖含量都大幅度降低,其中+Ca處理與對照最相近。脅迫3個低溫處理葉片果糖含量明顯高于對照,其中+Ca處理最高,其次為+H2O和+EGTA處理。恢復常溫7天,果糖含量大幅度降低,但處理間仍存在顯著差異,以+EGTA處理的果糖含量最高。低溫脅迫下+H2O處理淀粉含量最高,+Ca處理與對照無顯著差異,恢復常溫后淀粉含量增加,+Ca處理與對照無顯著差異。
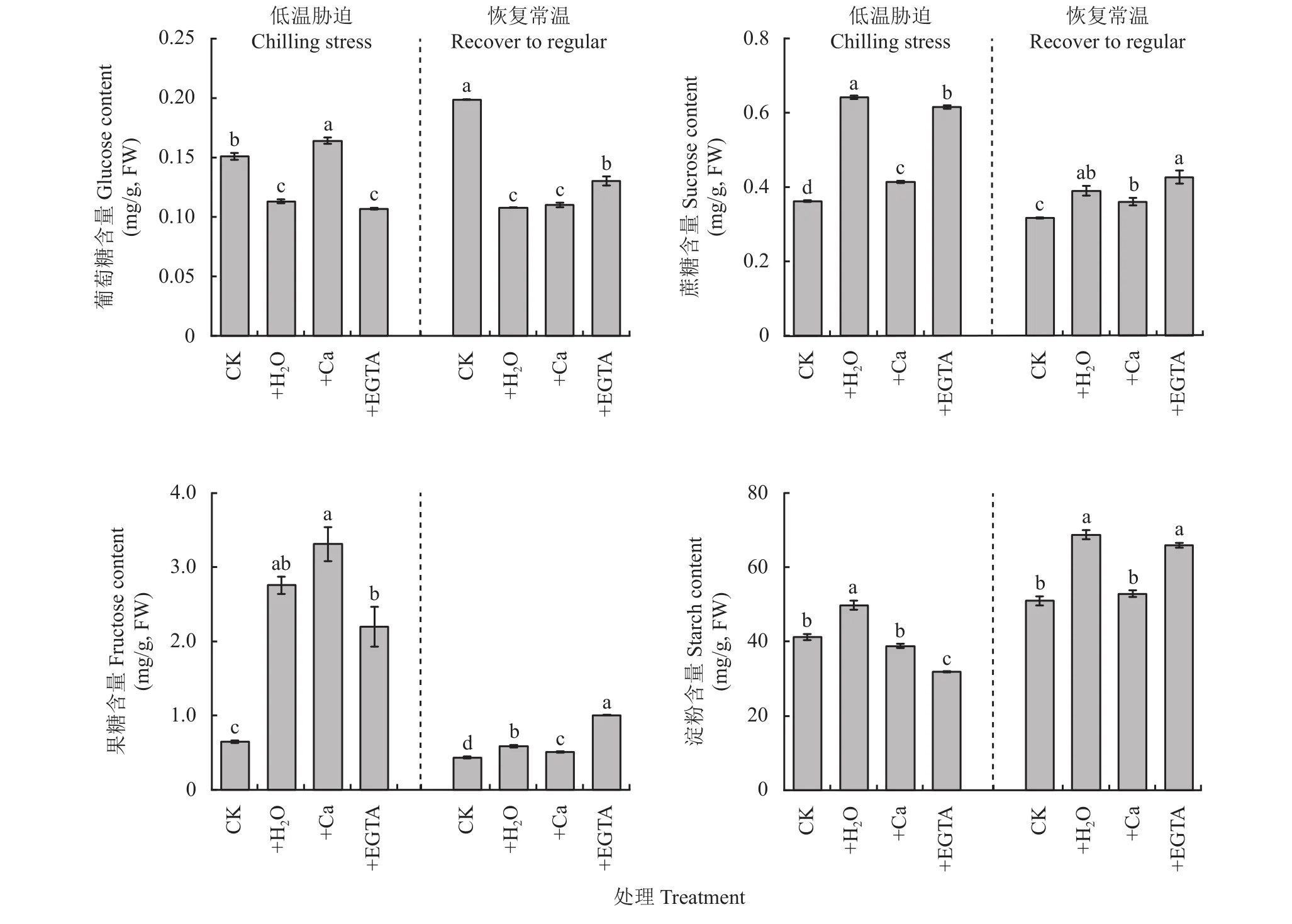
圖4 低溫脅迫7天及恢復常溫7天各處理花生葉片非結構性碳水化合物積累量Fig. 4 Non-structural carbohydrate accumulation in peanut leaves at 7 days of chilling stress and recovered to regular temperature under different treatments
2.6 低溫及常溫恢復條件下外源鈣素對花生葉片活性氧積累的影響
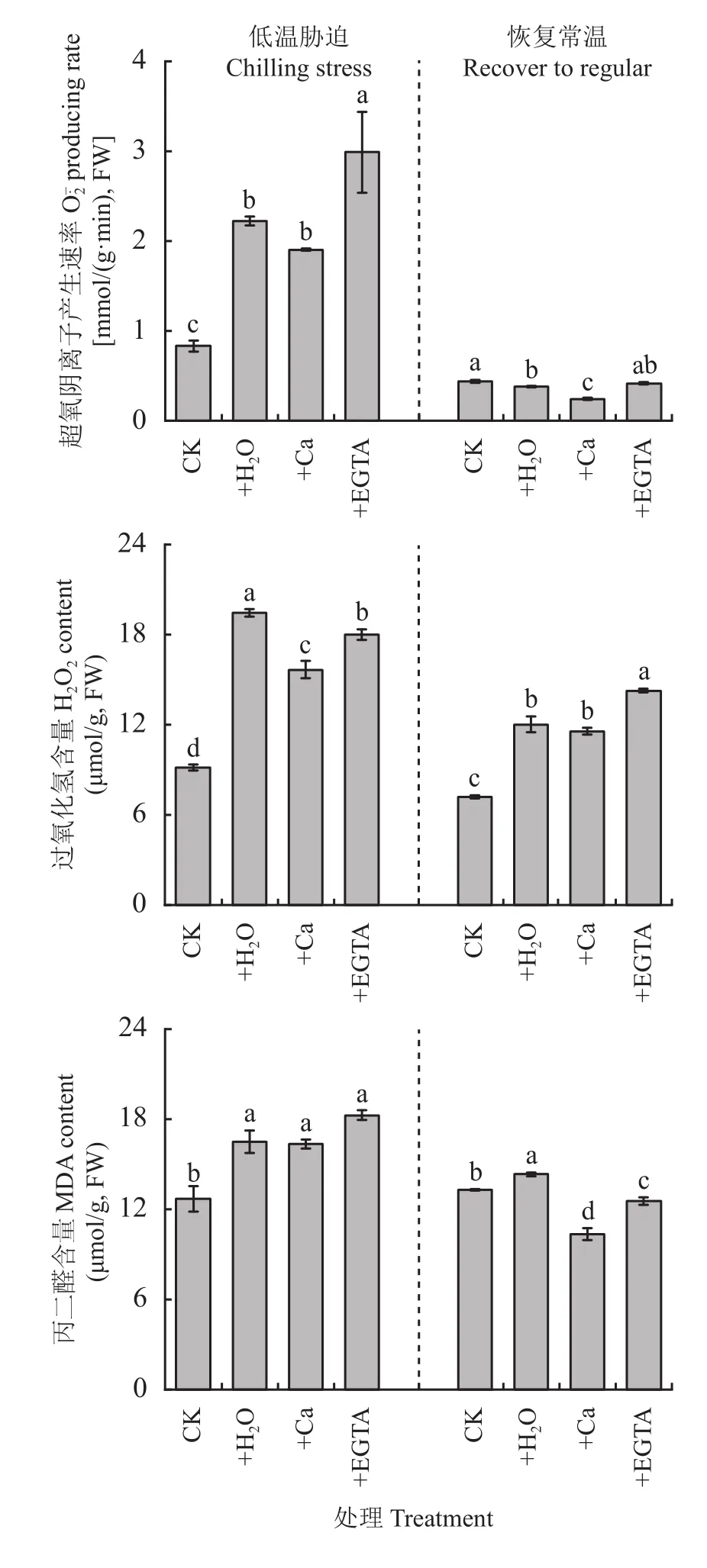
圖5 低溫脅迫7天及恢復常溫7天各處理花生葉片活性氧積累量和丙二醛含量Fig. 5 The reactive oxygen accumulation and MDA content in peanut leaves at 7 days of chilling stress and recovered to regular temperature under different treatments
3 討論
低溫影響植物的生長發育,減少生物量積累,低溫脅迫嚴重時造成不可逆傷害。花生屬于典型喜溫作物,低溫環境顯著影響花生生長發育,甚至造成缺苗斷壟、空殼減產,導致嚴重經濟損失。很多研究表明,在逆境條件下外源鈣能有效提升氣孔導度和凈光合速率等[23–24];同時也可提升Rubisco[25]與1,5-二磷酸核酮糖羧化酶的活性、增強類囊體中電子傳遞速率等進而增強光合效率[26]。本研究表明,低溫脅迫下與噴施H2O相比,葉面噴施外源Ca2+增加了花生株高、葉面積和葉綠素含量,也顯著緩解了低溫依賴性光損傷;有效緩解了花生葉片中非結構性碳水化合物的過量積累,增強了光合能力;有效降低了、H2O2和丙二醛的過量積累,保護了光合機構。前人研究表明,外源Ca2+間接緩解了夜間低溫脅迫造成的Gs和Tr的驟降,維持了Ci,防止了Pn的下降,Ca2+改善了光合碳水化合物在韌皮部的裝載和轉運等,外源Ca2+可以顯著修復低夜溫造成的光合膜損傷,緩解番茄、蘋果、煙草、花生等植物的光系統損傷,特別是緩解PSI光抑制[27–28]。以往的觀點認為,植物PSI比PSII更穩定而不易受到光損傷,然而新的研究表明,PSI復合物基質側的鐵硫簇極易受到光系統中活性氧簇(reactive oxygen species, ROS)的破壞[29–30],尤其是波動光強和光強–溫度交叉脅迫下PSI的光損傷更為嚴重,甚至PSI的光損傷要早于PSII。本研究發現,外源Ca2+有效緩解了低溫脅迫對花生生長的不利影響,特別是有效減緩了Fv/Fm的降低,這說明外源Ca2+有效恢復了低溫條件下花生葉片PSII的功能,降低了葉片中蔗糖的積累,降低活性氧帶來的損傷,有效提高了花生葉片的光合活性[31]。
前人研究表明,植物可溶性碳水化合物的積累在冬季抗寒生存中起著重要的作用[32],通常蔗糖是在低溫條件下積累的游離糖[33],另有國內外學者認為過多的碳水化合物積累使光合碳同化作用水平降低,并認為造成生物量減少的原因是碳水化合物的反饋抑制作用[34]。本研究發現,低溫脅迫使花生幼苗葉片中蔗糖、果糖及淀粉大量積累,Pn降低,ETRI和ETRII下降;噴施外源Ca2+減緩蔗糖、果糖、淀粉等的過度積累,減緩了ETR和Pn的降低,說明噴施Ca2+可減緩由非結構型碳水化合物過度積累造成的光合效率降低,從而提高了低溫脅迫下花生葉片的光合能力。Zhou等[35]、何潔等[36]研究認為,形成的大量淀粉粒會改變二氧化碳向細胞內擴散的途徑,影響葉綠體中二氧化碳的吸收與運輸使光合效率降低,出現光合速率和呼吸速率降低、同化物質運轉緩慢等應激反應,導致淀粉及可溶性糖的過度積累。由我們低溫脅迫后的恢復常溫的實驗結果可知,花生葉片中蔗糖、葡萄糖和果糖含量都顯著降低,淀粉含量依舊較高;外源施Ca2+處理有效降低非結構型碳水化合物積累量,并可恢復至對照水平。低溫阻礙葉片中蔗糖的運輸,大量非結構型碳水化合物的堆積會顯著抑制光合碳同化過程[37–38],這是使光合作用下降的主要原因之一;磷酸丙糖轉化成更多的淀粉,淀粉來不及轉化為蔗糖向外運出,導致蔗糖外運受到阻礙,進而葉片中積攢大量淀粉[39–41],最終產生嚴重的抑制作用。
有研究表明,碳水化合物與活性氧的關系密不可分,認為碳水化合物中的某些可溶性糖是活性氧代謝系統中重要的成分,可使植物抵抗過氧化損傷[42],也有研究表明活性氧的積累會上調可溶性糖的積累,從而適應非生物脅迫[43],活性氧在植物發育和生長過程及脅迫環境中起到調節作用,是不同脅迫與植物發育和生長控制之間的重要聯系[44–46]。本試驗結果顯示,在低溫脅迫后產生速率、H2O2含量以及MDA含量顯著增加,這與羅婭等[47]研究結果一致,外源鈣有效降低了、H2O2和丙二醛的大量積累,降低了光合機構的損傷。Ca2+在應對非生物脅迫及調節胞內信號也起著重要作用,可以抑制植物細胞膜脂過氧化,減輕活性氧的毒害作用。鈣和ROS最初被認為是相互作用的,因為鈣直接或者間接地對RBOHs(植物爆發式氧化酶,活性氧的主要來源)產生負調控,并可通過激活/抑制激酶/磷酸酶活性介導鈣通道的開閉[48]。本試驗結果顯示,外源Ca2+有效緩解ROS大幅度增加,噴施鈣離子抑制劑(EGTA)會顯著增加產生速率,進而反向證明了Ca2+可有效降低活性氧的產生。在低溫下ROS水平升高,ROS從源頭抑制了蛋白質的合成,特別抑制了D1蛋白的合成,D1蛋白是光系統II反應中心的組成部分[49]。在不利于CO2固定的環境條件下,光合色素吸收過多的光能會抑制光損傷PSII的修復,從而增加光抑制的程度。因此,ROS不僅抑制PSII蛋白的合成,而且抑制PSII修復速率[50–51]。此外,鈣可以穩定脂質層的基本結構,通過調節自身濃度減少ROS對膜脂過氧化損傷[52]。恢復常溫后,噴施鈣處理花生葉片丙二醛含量迅速下降[53],說明低溫脅迫及其恢復過程中,葉面噴施Ca2+有助于降低花生葉片丙二醛含量,增加植株冷適應性。
4 結論
低溫脅迫及其恢復過程中,外源Ca2+可以有效提升花生生長發育和光合作用水平,顯著降低非結構型碳水化合物和ROS的過度積累,全面緩解低溫光抑制。
