基于固液分離預處理的餐廚垃圾厭氧發酵
2022-03-29 12:36:40張靖雪李盼盼楊廣平
中國環境科學 2022年3期
張靖雪,李盼盼,于 洋,曾 蘇,余 冉*,楊廣平
基于固液分離預處理的餐廚垃圾厭氧發酵
張靖雪1,2,李盼盼1,于 洋1,曾 蘇1,余 冉1*,楊廣平3
(1.東南大學能源與環境學院,江蘇 南京 210096;2.南京環境集團有限公司,江蘇 南京 210046;3.南京清元景和環境科技有限公司,江蘇 南京 210019)
研究不同預處理方法所獲餐廚垃圾漿料的半連續式厭氧發酵效果,評價該類餐廚垃圾的產甲烷資源化潛能.結果表明,固液油三相分離預處理后的液相餐廚垃圾作為進料較固液混合相餐廚垃圾在厭氧發酵時具有更高的VS去除率,實際產甲烷潛能和甲烷轉化率,分別為91.2%, 531.5mLCH4/gVS和54.3%,表明液相餐廚垃圾半連續中溫濕式厭氧發酵具有良好的減量化和資源化效能.微生物群落分析表明,不同預處理方式影響了餐廚垃圾厭氧發酵系統的微生物群落演替,氨氮濃度是導致古菌群落轉移和豐度變化的關鍵因素.液相餐廚垃圾厭氧發酵時是以氫營養型產甲烷菌為主,而在固液混合相餐廚垃圾厭氧發酵時則以耐高濃度氨氮的多功能產甲烷菌為主.
餐廚垃圾;厭氧發酵;預處理;微生物群落
餐廚垃圾包括餐廳垃圾,菜市場垃圾和家庭廚房垃圾等,是城市生活垃圾的重要組成部分[1].由于餐廚垃圾具有高產量(占城市生活垃圾的37%~ 62%[2]),高含水率,高含油率,高鹽和高有機質占比(約占干重的95%~98%)的特點,易腐爛發臭,滋生蚊蠅,傳播疾病,如不及時處理,將對周圍環境造成嚴重影響.在中國,厭氧發酵和好氧堆肥技術被廣泛應用于餐廚垃圾的資源化處理[3].與堆肥相比,厭氧發酵具有碳排放量低,二次污染少,運營成本低,適合集中大規模處理等優點[4],且因其可產生清潔能源沼氣逐漸成為我國固廢資源化利用方式之一.
為了提高餐廚垃圾厭氧發酵的產沼氣性能,采用預處理可以提高底物的生物可降解性.預處理方法主要包括物理處理,化學處理,生物處理及多種處理方法的結合.目前國內在實際工程運用上對餐廚垃圾厭氧發酵的主要的預處理方式為機械破碎,加熱制漿和固液油三相分離組合方法以獲得固相,液相餐廚垃圾和油脂[5].其中油脂可回收制作生物柴油,而固相和液相垃圾則有兩種不同的處理方法,一種是將固相和液相垃圾混合均質化后作為底物加入厭氧發酵系統,另一種是僅把液相垃圾加入厭氧發酵系統,固相垃圾則干化后外運焚燒發電.已有研究[6]通過餐廚垃圾分選系統獲得了5種不同類型的餐廚垃圾并進行了厭氧發酵,結果表明餐廚垃圾漿液具有更高的甲烷產率.已知單位體積的液相餐廚垃圾底物相比固液混合相具有更多的可溶性有機物,而單位體積的固液混合相垃圾比液相垃圾具有更多的有機質.但二者對厭氧發酵的運行及有機質去除率,產甲烷潛能的影響尚缺乏相關研究.
本文以南京市某餐廚垃圾處置中心預處理環節所獲餐廚垃圾底物為研究對象,參考其厭氧發酵運行參數,進行半連續中溫濕式厭氧發酵試驗,探究不同預處理方式對餐廚垃圾厭氧發酵產甲烷的影響,并通過分析微生物群落變化解析了影響厭氧發酵產甲烷的主要因素,研究結果旨在為國內餐廚垃圾厭氧發酵的實際工程應用提供借鑒.
1 材料與方法
1.1 預處理樣品采集
厭氧發酵試驗餐廚垃圾取自南京市某餐廚垃圾處置中心固液油三相分離預處理后的液相垃圾和固液混合相垃圾,采集密封后在4℃下冷藏保存作為后續厭氧發酵底物.厭氧污泥取自該處置中心厭氧發酵罐.
1.2 揮發性固體含量去除率
揮發性固體含量(VS)去除率可以表征厭氧發酵的有機質去除效果,其計算方法可按照式(1)計算:

式中:r表示VS去除率,%;表示VS,%;表示總固體含量(TS),%;下標e表示出料,f表示進料[7].
1.3 實際產甲烷潛能
實際產甲烷潛能(EMP)計算公式見式(2):

式中:表示EMP,mLCH4/gVS;CH4為標準狀態下產生CH4的體積,mL;d為反應器內實際消耗的VS量,g[7].
1.4 餐廚垃圾半連續式中溫濕式厭氧發酵
啟動馴化階段,記為S0,將5L厭氧污泥置于反應罐中(圖1).氮吹密封后每日定時進出樣.當系統運行穩定后,首先每天定時進液相餐廚垃圾,連續運行30d,該階段記為S1階段;后將固液混合相混合垃圾底物以相同有機負荷加入厭氧發酵系統,連續運行30d,該階段記為S2階段,餐廚垃圾及厭氧污泥理化特性見表1,餐廚垃圾半連續式中溫濕式厭氧發酵運行參數見表2.每日記錄產氣量,每兩日收集氣體進行氣體成分分析,每日留樣做指標分析.
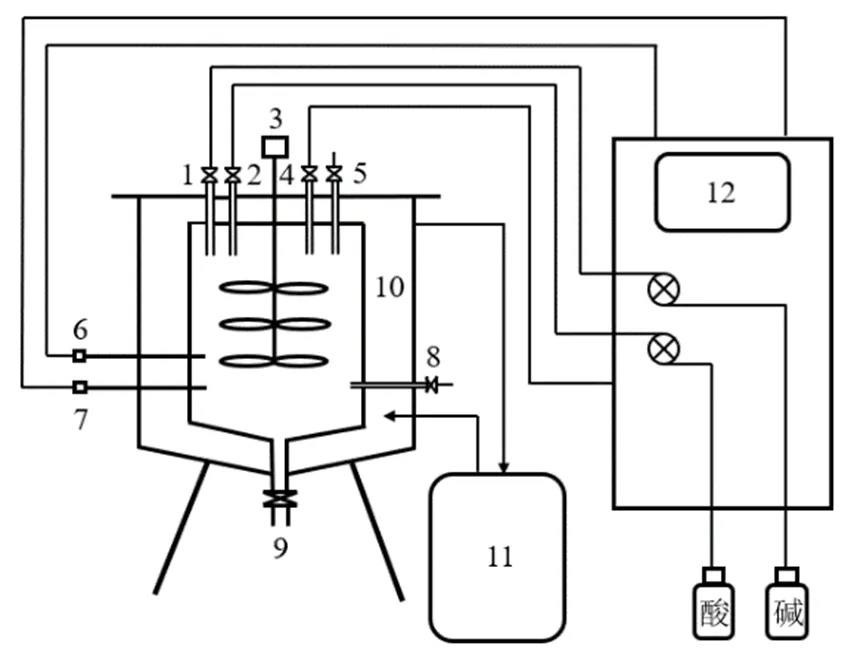
圖1 半連續厭氧發酵試驗裝置示意
1.補堿口; 2.補酸口; 3.機械攪拌器; 4.出氣口; 5.進樣口; 6.溫度監測探頭; 7.pH值監測探頭; 8.取樣口; 9.出料口; 10.控溫隔層; 11.循環水浴加熱器; 12.控制系統
1.5 微生物群落分析
試驗選取S1和S2厭氧發酵階段始末的污泥作樣品,編號分別為S1-1, S1-2, S2-2,其中因S1體系結束即為S2體系的開始,故S2體系的起始污泥樣品等同樣品S1-2.樣品DNA提取采用快速DNA提取法(FastDNA Spin Kit for Soil, MP土壤DNA提取試劑盒).將DNA樣品送至上海生工生物工程股份有限公司進行微生物高通量測序,使用16s V4引物515F和806R進行細菌群落分析,使用巢式PCR引物V3-V4進行兩輪擴增,第1輪引物340F和1000R,第2輪引物349F和806R進行古菌群落分析[8].
1.6 分析指標及測定方法
TS參考《生活垃圾采樣和物理分析方法》[9][CJ/T313-2009]的相關方法測定;VS參考《生活垃圾化學特性通用檢測方法》[10][CJ/T 96-2013]的灼燒法測定;含鹽量參考《食品中氯化物的測定》[11][GB 5009.44-2016]的直接滴定法測定;pH值采用pH計(Testo, 206pH2)測定;餐廚垃圾中C, N等有機元素采用元素分析儀測定(德國艾力蒙塔公司, Vario EL cube);甲烷濃度采用氣相色譜儀(普分科技有限公司,C3900)測定;揮發性脂肪酸(VFA)采用氣相色譜分析儀(Agilent, 7890B)測定;氨氮參考《水質氨氮的測定納氏試劑分光光度計法》[12][HJ 535-2009]測定;厭氧發酵產氣量采用排水法測定.分析指標測定每次設置3個平行樣,結果取平均值.

表1 餐廚垃圾及厭氧污泥理化特性
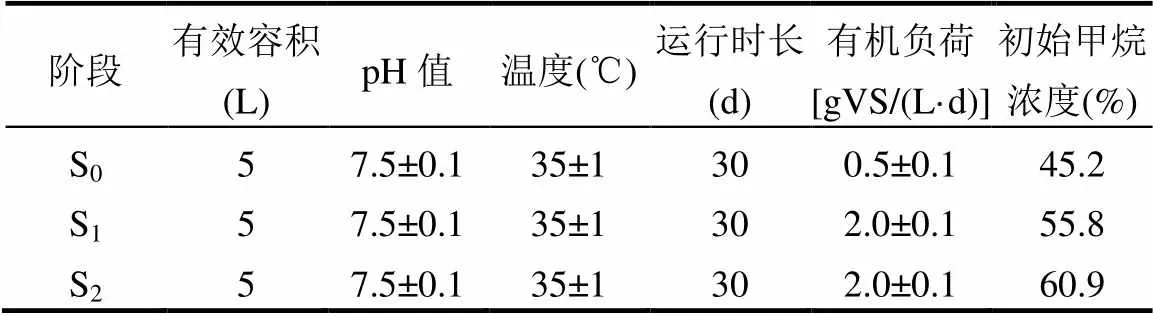
表2 厭氧發酵運行條件
2 結果與討論
2.1 沼氣成分和CH4日產量
從圖2可知,有機負荷從馴化期(S0)0.6gVS/(L·d)升至液相厭氧發酵階段(S1) 2.0gVS/(L·d)后,S1的CH4濃度在初始有小范圍上升,最高至63.5%,之后穩定在60.1%,固液混合相厭氧發酵階段(S2)的CH4濃度上升至61.3%.在產氣方面,S1穩定后的 CH4日產氣量為4859.3mL; S2的CH4日產氣量整體高于S1,其平均值為5276.0mL.固液混合相餐廚垃圾厭氧發酵S2段產生的CH4濃度和液相餐廚垃圾厭氧發酵S1段無顯著性差異,但CH4日產量在S2段高于S1段,而S2的CH4日產量波動較大,這可能與固液混合相餐廚垃圾的不均質有關.
2.2 VFA和氨氮
根據圖2可以看出,厭氧發酵正式運行后,由于有機負荷的提升,系統受到了沖擊負荷, VFA迅速積累后逐漸降低最后穩定的過程.這是因為馴化期轉液相餐廚垃圾厭氧發酵時有機負荷上調,餐廚垃圾被產酸菌快速降解,后產甲烷菌活性增強將VFA降解轉化.在厭氧發酵進行12d后丙酸含量已低出檢測限,VFA主要以乙酸的形態存在,S1和S2兩個階段并無明顯差異.VFA趨于穩定后在2.0gVS/(L·d)有機負荷下濃度低于200mg/L,可知該半連續式中溫濕式厭氧發酵系統可以在較高濃度的有機負荷下正常運行.氨氮積累方面,S1階段氨氮積累較緩慢,經過30d半連續厭氧發酵后氨氮增長了16.9%,而S2階段氨氮積累較快,氨氮增長了29.0%.發酵液中的氨氮主要來自于微生物對蛋白質等有機物的降解,以游離氨和銨根離子的形式存在,其中游離氨被認為是抑制厭氧微生物的主要原因.餐廚垃圾經過固液油三相分離后的固液混合相垃圾在單位體積下比液相垃圾具有更多的蛋白質,更多的有機氮可轉化為無機氮,導致分解蛋白質產生的氨氮大于微生物消耗氨氮的量,從而氨氮濃度呈上升趨勢,長期運行會導致厭氧微生物尤其是產甲烷菌生長受到抑制[13].結合EMP可知S2氨氮的快速積累抑制了厭氧發酵的有機質轉化效率.結合表1可知,厭氧發酵所用餐廚垃圾的C/N偏低,即氮元素太多造成氨氮不能被微生物及時轉化利用,導致系統的氨氮濃度升高,故提高厭氧發酵底物碳源,調節C/N至20~30可緩解氨氮積累.

圖2 餐廚垃圾半連續式厭氧發酵運行性能
2.3 VS去除率和EMP
從圖2可知,雖然S1和S2階段VS去除率平均值相近,分別為91.2%和91.0%,但S1的VS去除率是逐漸上升的趨勢,VS去除率維持在87.3%~92.9%,而在S2階段整體呈下降趨勢,VS去除率維持在89.4%~92.6%.對比已有文獻[7,14]報道的中溫半連續餐廚垃圾厭氧發酵VS去除率在85%~94%,本實驗S1和S2兩階段VS去除率均處于較高水平.而液相垃圾厭氧發酵相較固液混合相垃圾厭氧發酵具有更高的VS去除率,一方面是因為氨氮快速積累抑制固液混合相垃圾厭氧發酵,另一方面因為液相垃圾的小分子和可溶性有機質占比一般高于粒徑不一的固液混合相垃圾,生物可降解性更高.從圖2可以看出,EMP在S1階段逐漸上升,最高時可達到633.0mLCH4/gVS,平均值為531.5mLCH4/gVS;與之對比在S2階段,EMP值整體下降,最高時僅達到545.8mLCH4/gVS,平均值為465.8mLCH4/gVS,且數值較為波動.對比Tampio等[15]進行中溫半連續餐廚垃圾厭氧發酵實驗,在3gVS/(L·d)的有機負荷下得到最高483mLCH4/gVS的甲烷產率,本厭氧發酵試驗在2gVS/(L·d)有機負荷下具有更高EMP. Zhang等[6]對5種不同類型的餐廚垃圾進行先中溫35℃后高溫55℃的厭氧發酵結果表明,餐廚和蔬菜殘渣混合物壓濾后的漿液厭氧發酵表現出最高的比甲烷產率,而植物殘渣經過壓濾的漿液厭氧發酵最低.故液相餐廚垃圾因為其具有大量的可溶性有機質從而具有更高的EMP,在半連續式中溫濕式厭氧發酵工藝中液相餐廚垃圾比固液混合相餐廚垃圾產甲烷效率更高.
2.4 厭氧發酵系統物料平衡
純液相和固液混合相餐廚垃圾厭氧發酵2個階段每日進料150mL,純液相餐廚垃圾VS為6.6%,固液混合相餐廚垃圾的VS為8.3%.2個階段各選擇穩定時即反應第15d和16d的數據進行碳平衡計算,根據質量守恒可得式(3):
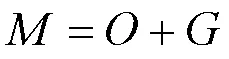
式中:為反應器每天進料的總碳質量,g;為反應器每天出料的總碳質量,g;為每天產生的沼氣總碳質量,g.已知兩個階段選取的第15d和16d數據見表3.其中總碳TC≈VS×47%[16].

表3 碳平衡計算選取參數
注:進料碳含量以干基計算.
可計算出S1階段和S2階段,,和TC利用率及甲烷轉化率(表4).由碳平衡計算可知S1階段的TC利用率和甲烷轉化率比S2階段分別高4.3%和1.4%,即液相餐廚垃圾進行半連續式中溫濕式厭氧發酵時可以將更多的有機質轉化為沼氣和甲烷.

表4 餐廚垃圾厭氧發酵碳平衡計算
2.5 厭氧發酵微生物群落分析
如表5所示,S1階段隨著厭氧發酵的進行,細菌群落分布豐富度(Chao1和ACE指數)呈增大的趨勢而S2階段在降低.用來估算群落分布多樣性的Shannon指數值越大,說明群落多樣性越高,而Simpson指數數值越大說明群落多樣性越低.S1階段Shannon指數和Simpson指數分別降低和增加,表示細菌群落多樣性在降低.與之對比S2階段細菌群落多樣性則表現出增大的趨勢.
S1階段隨著發酵的進行古菌群落豐富度(Chao1和ACE指數)無顯著變化,而古菌群落多樣性(Shannon和Simpson指數)在增大.在S2階段隨著厭氧發酵的進行,古菌群落豐富度和多樣性均在減小.
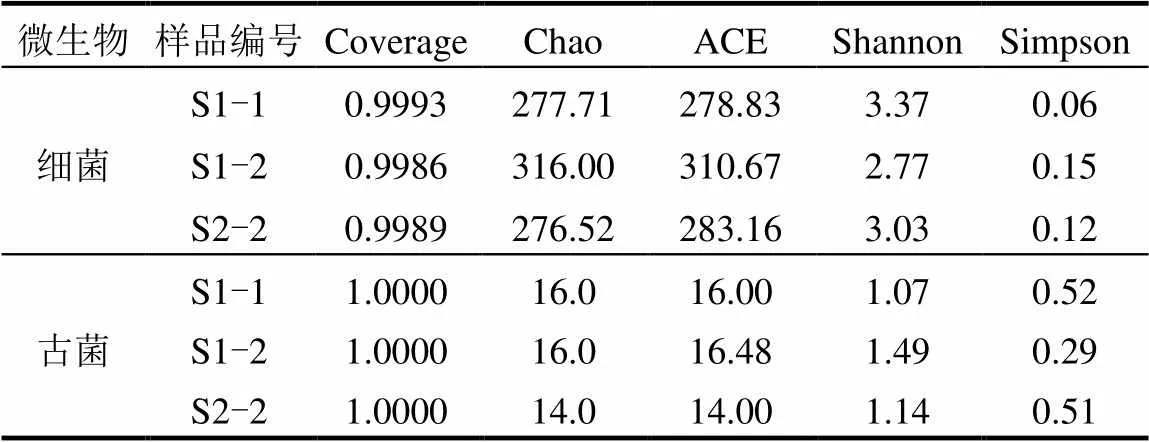
表5 不同預處理進料條件下的厭氧發酵反應器中細菌和古菌群落分析
如圖3所示,因為兩個厭氧發酵階段的古菌在門水平上均屬于Euryarchaeota,故在屬水平上進行群落結構分析.圖中橫坐標為樣品編號,縱坐標為相對豐度比例.顏色對應了此分類水平各物種的名稱,顏色寬度表示不同物種相對豐度比例.

圖3 餐廚垃圾厭氧發酵反應器中細菌群落在門水平和古菌群落在屬水平上的相對豐度
如圖3所示,Bacteroidetes, Cloacimonetes和Firmicutes是兩個厭氧發酵階段中最主要的細菌門類.其中Bacteroidetes的微生物菌群被報道存于在碳水化合物發酵,生理穩態和類固醇代謝中,可以生產各種分解酶,包括降解復雜的有機化合物的水解酶,蛋白酶和脂肪酶[17-19]. Cloacimonetes被報道與其他耗氫和利用乙酸鹽的細菌共同作用時,可以在低氨氮條件下降解纖維素,參與丙酸的共氧氧化和氨基酸降解為乙酸鹽,CO2和H2[20]. Firmicutes由大量分泌裂解酶包括纖維素酶,脂肪酶,蛋白酶和其它胞外酶的細菌組成,由于可以形成孢子,這些細菌可以在極端環境條件下存活[21-22].在S1體系開始時Bacteroidetes(60.8%)和Firmicutes(22.4%)占優勢,在厭氧發酵結束時Bacteroidetes(61.6%)無明顯變化,而Cloacimonetes(14.4%)成為優勢門之一, Firmicutes(9.8%)降低了12.6%成為第3優勢門.在S2階段Bacteroidetes(43.8%)相對豐度相比起始降低了17.8%,Cloacimonetes(28.1%)和Firmicutes(16.85%)分別增長了13.8%和7.1%. Cloacimonetes的出現并在S2體系內作為優勢門,說明固液混合相餐廚垃圾的復雜成分使體系內的功能菌群發生了變化,固液混合相餐廚垃圾厭氧發酵體系除了需要Bacteroidetes和Firmicutes分泌大量水解酶來降解大分子有機物,還需要Cloacimonetes的協同降解纖維素、氨基酸等.
由圖3可以看出,和是2個厭氧發酵階段中產甲烷古菌的主要優勢屬.可以利用甲醇,甲胺,乙酸鹽以及H2和CO2生產甲烷,能夠利用不少于9種的產甲烷基質,且具有較快的生長速率可以快速適應周圍環境變化[23-24].是氫營養型產甲烷菌,可以利用H2和甲酸鹽作為主要的電子供體將CO2還原為CH4[25]. S2階段中相對豐度較高的和同為氫營養型產甲烷菌,都是利用H2作為電子供體將CO2還原為CH4[25-26].在S1階段可以看出隨著厭氧發酵的進行相對豐度下降37%,與之對應相對豐度升高至49.0%,成為主要優勢屬.在S2階段回到了最高相對豐度(70.1%),下降至7.2%,其相對豐度下降了41.8%. S1反應體系內主要為氫營養型產甲烷菌,占據了相對豐度的57.9%, S2反應體系內主要為多功能的為主,占據了相對豐度的70.1%,其可以在甲基營養型,乙酸營養型和氫營養型三種模式下產甲烷[27].S2階段的大量富集,是因為對高濃度的氨氮具有更強的抵抗力[28-29],結合兩個厭氧發酵階段的氨氮濃度變化,可以解釋該富集現象.與之相反因為S1階段的氨氮濃度相對較低,適宜更多種類的產甲烷菌生長,所以氫營養型和乙酸營養型產甲烷菌開始富集.結合表5的2個厭氧發酵階段中古菌多樣性分析,與S2相比S1具有更高的群落分布豐富度和多樣性,證明了氨氮濃度是導致古菌種群轉移的關鍵因素[30].
3 結論
3.1 固液油三相分離預處理后,液相餐廚垃圾比固液混合相餐廚垃圾在厭氧發酵過程中具有更高的VS去除率(91.2%),實際產甲烷潛能(531.5mLCH4/ gVS)和甲烷轉化率(54.3%),且氨氮積累較慢.
3.2 微生物群落分析表明氨氮濃度是導致古菌群落轉移和豐度變化的關鍵因素,在液相餐廚垃圾厭氧發酵時因其氨氮濃度較低,致使古菌群落具有更高的豐富度和多樣性,古菌群落向氫營養型和乙酸營養型產甲烷模式進化;而在固液混合相餐廚垃圾厭氧發酵時因其氨氮濃度較高使古菌群落以耐高濃度氨氮的多功能產甲烷菌為主.
[1] Jin C, Sun S, Yang D, et al. Anaerobic digestion: An alternative resource treatment option for food waste in China [J]. Science of The Total Environment, 2021,779:146397.
[2] 湯 晴.餐廚垃圾厭氧制沼及沼氣異位生物提純技術研究 [D]. 無錫:江南大學, 2019.
Tang Q. The biogas generation from food waste by anaerobic digestion and its ex-situ biological upgrading technology [D]. Wuxi: Jiangnan University, 2019.
[3] Lin L, Xu F, Ge X, et al. Improving the sustainability of organic waste management practices in the food-energy-water nexus: A comparative review of anaerobic digestion and composting [J]. Renewable and Sustainable Energy Reviews, 2018,89:151-167.
[4] Li Y, Jin Y, Borrion A, et al. Current status of food waste generation and management in China [J]. Bioresource Technology, 2019,273: 654-665.
[5] 文 斐.我國餐廚垃圾處理工藝技術路線選擇與分析 [J]. 環境與發展, 2020,32(9):107+109.
Wen F. Selection and analysis of food waste treatment process in China [J]. Environment and Development, 2020,32(9):107+109.
[6] Zhang J, Hu Q, Qu Y, et al. Integrating food waste sorting system with anaerobic digestion and gasification for hydrogen and methane co-production [J]. Applied Energy, 2020,257:113988.
[7] 何 琴,李 蕾,何清明,等.重慶主城餐廚垃圾理化性質及產甲烷潛能分析 [J]. 環境化學, 2014,33(12):2191-2197.
He Q, Li L, He Q M, et al. Physical and chemical properties and methane production potential of food waste in Chongqing City [J]. Environmental Chemistry, 2014,33(12):2191?2197.
[8] 王 麗.廚余-污泥混合干式厭氧發酵及其微生物學機制研究 [D]. 西安:西安工程大學, 2019.
Wang L. Study on dry anaerobic fermentation of kitchen waste-sludge and its microbiological mechanism [D]. Xi'an: Xi'an Polytechnic University, 2019.
[9] CJ/T313-2009 生活垃圾采樣和物理分析方法 [S].
CJ/T313-2009 Sampling and physical analysis methods of domestic waste [S].
[10] CJ/T 96-2013 生活垃圾化學特性通用檢測方法 [S].
CJ/T 96-2013 General test method for chemical characteristics of domestic waste [S].
[11] GB 5009.44-2016 食品中氯化物的測定 [S].
GB 5009.44-2016 Determination of chloride in food [S].
[12] HJ 535-2009 水質氨氮的測定納氏試劑分光光度計法 [S].
HJ 535-2009 Water quality determination of ammonia nitrogen nessler reagent spectrophotometric method [S].
[13] 張達飛.餐廚垃圾厭氧消化運行實驗研究 [D]. 北京:北京建筑大學, 2017.
Zhang D F. Experimental research on the anaerobic digestion of food waste [D]. Beijing: Beijing University of Civil Engineering and Architecture, 2017.
[14] 李榮平,葛亞軍,王奎升,等.餐廚垃圾特性及其厭氧消化性能研究 [J]. 可再生能源, 2010,28(1):76-80.
Li R P, Ge Y J, Wang K S, et al. Characteristics and anaerobic digestion performances of kitchen wastes [J]. Renewable Energy Resources, 2010,28(1):76?80.
[15] Tampio E, Ervasti S, Paavola T, et al. Anaerobic digestion of autoclaved and untreated food waste [J]. Waste Management, 2014, 34(2):370-377.
[16] 錢小青.泔腳廢物厭氧兩相發酵工藝及其礦化垃圾協同生物產氫過程研究 [D]. 上海:同濟大學, 2006.
Qian X Q. Anaerobic digetion and biological hydrogen production from food origin wastes in the presence of aged refuse [D]. Shanghai: Tongji University, 2006.
[17] Alalawy A I, Guo Z, Almutairi F M, et al. Explication of structural variations in the bacterial and archaeal community of anaerobic digestion sludges: An insight through metagenomics [J]. Journal of Environmental Chemical Engineering, 2021,9(5):105910.
[18] Marchesi J R, Adams D H, Fava F, et al. The gut microbiota and host health: A new clinical frontier [J]. Gut, 2016,65(2):330-339.
[19] Rivière D, Desvignes V, Pelletier E, et al. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge [J]. ISME Journal, 2009,3(6):700-714.
[20] Stolze Y, Bremges A, Maus I, et al. Targeted in situ metatranscriptomics for selected taxa from mesophilic and thermophilic biogas plants [J]. Microbial Biotechnology, 2018,11(4): 667-679.
[21] Pasalari H, Gholami M, Rezaee A, et al. Perspectives on microbial community in anaerobic digestion with emphasis on environmental parameters: A systematic review [J]. Chemosphere, 2020,270:128618.
[22] Lim J W, Chiam J A, Wang J-Y. Microbial community structure reveals how microaeration improves fermentation during anaerobic co-digestion of brown water and food waste [J]. Bioresource Technology, 2014,171:132-138.
[23] Jenney F E, Adams M W W. Hydrogenases of the model hyperthermophiles [J]. Annals of the New York Academy of Sciences, 2008,1125(1):252-266.
[24] Goberna M, Gadermaier M, Franke-Whittle I H, et al. Start-up strategies in manure-fed biogas reactors: Process parameters and methanogenic communities [J]. Biomass and Bioenergy, 2015,75:46- 56.
[25] Guo J H, Peng Y Z, Ni B J, et al. Dissecting microbial community structure and methane-producing pathways of a full-scale anaerobic reactor digesting activated sludge from wastewater treatment by metagenomic sequencing [J]. Microb. Cell. Fact., 2015,14:11.
[26] Cadillo-Quiroz H, Brauer S L, Goodson N, et al. Methanobacterium paludis sp nov and a novel strain of Methanobacterium lacus isolated from northern peatlands [J]. International Journal of Systematic and Evolutionary Microbiology, 2014,64:1473-1480.
[27] Mehta P, Deshmukh K, Dagar S S, et al. Genome sequencing and analysis of a psychrotrophic methanogen Methanosarcina sp. nov. MSH10X1cultured from methane hydrate deposits of Krishna Godavari Basin of India [J]. Marine Genomics, 2021:100864.
[28] Zhang J, Lv C, Tong J, et al. Optimization and microbial community analysis of anaerobic co-digestion of food waste and sewage sludge based on microwave pretreatment [J]. Bioresource Technology, 2016, 200:253-261.
[29] Poirier S, Desmond-Le Quéméner E, Madigou C, et al. Anaerobic digestion of biowaste under extreme ammonia concentration: Identification of key microbial phylotypes [J]. Bioresource Technology, 2016,207:92-101.
[30] Zamanzadeh M, Hagen L H, Svensson K, et al. Anaerobic digestion of food waste – Effect of recirculation and temperature on performance and microbiology [J]. Water Research, 2016,96:246-254.
Study on anaerobic digestion of kitchen waste based on solid-liquid separation pretreatment.
ZHANG Jing-xue1,2, LI Pan-pan1, YU Yang1, ZENG Su1, YU Ran1*, YANG Guang-ping3
(1.Department of Energy and Environment, Southeast University, Nanjing 210096, China;2.Nanjing Environment Group Co.LTD, Nanjing 210046, China;3.Chinair Environmental Science and Technology Co. LTD, Nanjing 210019, China)., 2022,42(3):1252~1258
A semi-continuous anaerobic digestion (AD) experiment was set up to evaluate the methanogenesis potential of kitchen waste (KW) after different pretreatment modes. After the solid, liquid and oil three-phase separation pretreatment, the liquid-phase KW achieved a higher volatile solid (VS) removal rate, experimental methane potential (EMP) and methane conversion rate than the mixed one of solid-liquid phases, which were 91.2%, 531.5mLCH4/gVS, and 54.3%, respectively. The results indicated that the medium-temperature wet AD displayed a satisfying effect on KW reduction and resource utilization when using the liquid-phase KW as the feedstock. The microbial community analysis showed that the microbial community succession in the AD system was affected by the KW pretreatment method. The ammonia concentration was the key factor to induce the variation and abundance shifts of the archaeal community. The hydrogenotrophic methanogen,was the dominant methanogens in the AD system with the KW liquid phase as the feedstock. The, which was reported to be able to resist high concentrations of ammonia nitrogen was the dominant methanogens in the AD system with the feedstock of the KW solid-liquid phase mixture.
kitchen waste;anaerobic digestion;pretreatment;microbial community
X705
A
1000-6923(2022)03-1252-07
張靖雪(1995-),女,河南平頂山人,東南大學博士研究生,主要從事固體廢棄物處理與處置研究.
2021-08-01
2020年度江蘇省建設系統科技項目(指導類)(2020ZD61)
*責任作者, 教授, yuran@seu.edu.cn
