黃瓜纖維素合成酶CESA家族基因鑒定及非生物脅迫響應分析
2022-04-02 00:16:00趙雪芳李亞棟田哲娟吳志明
中國蔬菜 2022年3期
康 忱 趙雪芳 王 鵬 李亞棟 田哲娟 吳志明
(河北省農林科學院經濟作物研究所,河北石家莊 050051)
纖維素是植物細胞壁的主要組成成分,具有生成分子內與分子間氧鍵的能力。在植物體內,纖維素構成纖維素糖苷網,是承受機械壓力的支撐結構,能夠調控植物形態學上的某些特性與特征(Joshi &Mansfield,2007)。細胞壁在植物生長發育和響應環境脅迫中發揮重要作用,因此纖維素合成機制的相關研究在多個學科中占有重要地位(Pauly &Keegstra,2010;Youngs &Somerville,2012)。纖維素由纖維素合成酶復合體合成,其單體主要由纖維素合成酶(cellulose synthase,CESA)基因家族成員編碼,纖維素合成酶負責植物纖維素的生物合成(Somerville,2006)。目前已從多種植物中克隆多個基因的相關序列,如擬南芥()、煙草()、水稻()、毛果楊()、四倍體海島棉()和亞麻()等(Richmond &Somerville,2000;Wang et al.,2010;袁紅梅 等,2016;Yuan et al.,2016;徐宗昌和孔英珍,2017)。在擬南芥中,初生壁纖維素合成需要纖維素合成酶AtCESA1、AtCESA3 和AtCESA6 的參與,三者組成纖維素合成酶復合體,次生壁纖維素合成需要AtCESA4、AtCESA7 和AtCESA8 的參與(Desprez et al.,2007;Atanassov et al.,2009;Hill et al.,2015)。纖維素合成酶的互作蛋白(cellulose synthase interactive protein 1,CSI1)和伴侶蛋白(companion of cellulose synthase,CC)是微管和纖維素合成酶復合體的相互作用中的主要成員,其中CC 蛋白具有在鹽脅迫條件下維持纖維素合成的作用(Gu et al.,2010;Endler et al.,2015)。擬南芥AtCESA5 的磷酸化則是光敏色素調控纖維素合成和下胚軸細胞伸長生長的重要機制(Chen et al.,2010,2016;Bischoff et al.,2011)。磷酸化蛋白組學方面的研究顯示,磷酸化修飾CESAs 能響應植物生長發育及環境信號,調控由微管介導的纖維素合成的關鍵機制(Facette et al.,2013;Roitinger et al.,2015;Chen et al.,2016)。
黃瓜(L.)為葫蘆科一年生蔓生或攀援草本植物,是我國重要的蔬菜作物之一,在世界范圍內廣泛種植。2020 年我國黃瓜年種植面積達到100 萬hm,年產量達到5 600 萬t,占世界總產量的70%以上(FAO,http://www.fao.org/)。在生產過程中,黃瓜經常遭受各種非生物脅迫,例如低溫、高溫和鹽脅迫。低溫脅迫導致葉片發黃和生長減慢,降低黃瓜的產量,特別是在冬季和早春(Dong et al.,2019)。高溫導致葉片曬傷和果實畸形,從而影響夏季黃瓜的產量和果實品質(于拴倉和王永健,2003)。鹽脅迫降低了葉綠素含量和礦物質吸收,從而影響了黃瓜的正常生長發育(Yildirim et al.,2008)。植物激素在調節外界環境應激反應中很重要。脫落酸(ABA)信號傳導途徑是植物中鹽、干旱和低溫脅迫信號傳導的中心(Zhu,2016)。ABA 的生物合成和信號傳導參與植物對低溫的響應(Wang et al.,2019)。到目前為止,在黃瓜對非生物脅迫的響應中,已經對WRKY(Ling et al.,2011)、MADS(Hu &Liu,2012)、MLO(Zhou et al.,2013)、ARF(Liu &Hu,2013)、NAC(Liu et al.,2018)和GRAS(Li et al.,2020)等基因家族進行了深入研究,但是,CESAs 作為植物生長發育和非生物脅迫響應過程中的一個重要的基因家族,在黃瓜中還鮮見報道。目前纖維素合成酶基因家族的研究有較大進展,但大多局限于擬南芥、煙草等模式植物和棉花、楊樹、亞麻、毛竹等木本植物中,黃瓜中絕大部分CESA家族成員未得到鑒定,隨著黃瓜基因組序列的公布和完善,對纖維素合成酶家族基因進行全基因組挖掘和全面、系統的分析十分必要。本試驗利用生物信息學方法對黃瓜CESA 基因家族成員進行鑒定,并對它們的染色體位置、基因結構、進化分化特征以及物種間的共線性進行了系統分析。通過施加外源植物激素(ABA 和MeJA)和非生物脅迫(NaCl)處理,分析CESA 家族基因的表達模式,為更好地了解CESA 基因家族成員及其在黃瓜非生物脅迫響應中的生物學功能提供依據,為進一步探索該家族在黃瓜響應非生物脅迫調節中的作用奠定基礎。
1 材料與方法
1.1 材料和處理
供試材料為河北省農林科學院經濟作物研究所保存的黃瓜品系9930。2021 年3 月上旬將供試黃瓜材料播種于本所育苗溫室,選取兩葉一心時期的黃瓜幼苗,在塑料花盆中培養,第1 組用200 mmol·LNaCl 水培處理,第2 組用100 μmol·L的脫落酸(ABA)噴灑葉面,第3 組用100 μmol·L的茉莉酸甲酯(MeJA)噴灑葉面,第2 組和第3 組利用滅菌的營養土和蛭石混合栽培。將所有處理放置于植物組織培養室中培養,溫度為28 ℃,光照強度為2 000 lx,光照周期為光16 h/暗8 h。分別在處理后0、3、6、12 h 和24 h 對葉片取樣,經液氮速凍后置于-80 ℃冰箱中保存用于RNA 提取。每個處理采用3 個生物學重復。
1.2 黃瓜纖維素合成酶基因家族成員的獲得與 鑒定
黃瓜基因組序列于2009 年完成(Huang et al.,2009)。從黃瓜基因組數據庫CuGenDB(Cucurbit Genomics Database,http://www.cucurbitgenomics.org/organism/2)下載“黃瓜基因組9930_V2 版本”(9930 作為參考基因組)基因組數據。從TAIR數據庫(https://www.arabidopsis.org/)獲得擬南芥CESA 基因家族的基因ID 和氨基酸序列信息。利用BioEdit 軟件,以e為臨界值選取候選CESA 基因,用本地Perl 語言提取候選基因蛋白序列,初步獲得CESA 基因家族成員。結合下載于Pfam數據庫(https://pfam.xfam.org/)的CESA 蛋白的保守結構域(PF03352,PF14569),使用Pfam 中的HMMScan 和PfamScan 對上一步獲得的候選基因進行結構域分析,剔除不含CESA 蛋白典型結構域的序列,確定CESA 基因家族的最終成員。
1.3 黃瓜纖維素合成酶基因家族生物信息學分析和系統進化樹構建
通過ExPASy網站(https://web.expasy.org/compute_pi/)對黃瓜纖維素合成酶家族蛋白的等電點(pI)和分子量(Mw)進行預測。利用在線網站MEME(https://meme-suite.org/meme/index.html)進行motif 預測,數量設置為10 個,其余參數選擇默認。提取CESA 基因家族成員的gff文件信息,通過網站Gene Structure Display Server 2.0(GSDS 2.0,http://gsds.gao-lab.org/)進行基因結構分析。利用在線軟件Softberry(http://linux1.softberry.com/berry.phtml)對家族蛋白進行亞細胞定位的預測。從黃瓜基因組序列注釋文件中提取每個基因的染色體位置,利用在線網站MapGene2Chrom web v2(MG2C,http://mg2c.iask.in/mg2c_v2.0/)繪制基因在各條染色體上的位置圖譜。利用TBtools 中的MCscanX 分析黃瓜、擬南芥和番茄CESA 家族基因之間以及黃瓜、西瓜和甜瓜CESA 家族基因之間的同源性,生成共線性分析圖。
使用軟件MEGA 7.0 分別對黃瓜、擬南芥、番茄和黃瓜、西瓜、甜瓜中CESA 的蛋白序列進行多重比較,通過鄰接法(neighbor-joining,NJ)構建進化樹,設BootStrap 抽樣次數為1 000。使用在線網站ITOL(https://itol/embl.de/)優化進化樹,并將CESA 家族進一步分為不同group。
1.4 黃瓜纖維素合成酶家族基因啟動子區順式作用元件分析
為了分析黃瓜CESA 基因啟動子區域包含的順式作用元件,用軟件TBtools 提取每個基因起始密碼子‘ATG’上游2 000 bp 的序列,利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)對順式作用元件進行預測和分析,然后使用TBtools 軟件對分析結果進行可視化。
1.5 基于轉錄組測序數據的基因組織表達與脅迫響應分析
為了確定基因的組織特異性表達,從黃瓜基因組數據庫CuGenDB 下載并分析了已發表的RNA-seq 數據,分別為10 個黃瓜組織〔根、莖、葉、受精子房(已膨大)、子房(未膨大)、雌花、未受精子房(已膨大)、雄花、卷須基部和卷須〕(PRJNA80169)、NaCl處理(PRJNA437579)和低溫處理(PRJNA438923)的轉錄組數據。
1.6 黃瓜纖維素合成酶家族基因表達分析
使用植物總RNA 提取試劑盒〔天根生化科技(北京)有限公司〕從樣品中提取總RNA,使用PrimeScript ? RT reagent Kit 試劑盒〔寶日醫生物技術(北京)有限公司〕完成第一鏈cDNA 的合成,使用的儀器為ABI PRISM 7500 實時熒光定量PCR儀(操作軟件為7500 Fast Real-Time PCR Systems,v2.0.6,USA),熒光染料為TB Green。試劑盒為寶生物工程(大連)有限公司的TB Green? Premix Ex Taq ?(Tli RNaseH Plus)。使用Primer Premier 3 在線設計特異性引物,序列如表1 所示。qRT-PCR 反應體系10 μL:5 μL 的 TB Green Premix Ex(Tli RNaseH Plus)(2×),1 μL cDNA模板,primer-F/R 各0.2 μL(終濃度為0.2 μmol·L),0.2 μL ROX Reference Dye II(50×),3.4 μL ddHO。反應程序:95 ℃預變性30 s;95 ℃變性5 s,60 ℃延伸34 s,40 個循環;72 ℃ 20 s。以(Csa3G806800)為內參基因。使用2算法分析基因的相對表達水平(Rieu &Powers,2009)。

表1 qRT-PCR 引物序列
2 結果與分析
2.1 黃瓜纖維素合成酶基因家族的成員鑒定
利用已發表的擬南芥CESA 的蛋白序列作為源序列,通過BlastP 的方式采用BioEdit 軟件對黃瓜基因組蛋白數據庫進行比對,以E <10為篩選條件,結合CESA 蛋白的保守結構域(PF03352,PF14569)共鑒定得到35 個CESA 基因家族成員。根據在染色體上的位置信息,利用在線軟件MG2C 進行基因定位并繪制其在染色體上的位置圖譜(圖1),35 個基因均分布在除5 號染色體外的7 條染色體上,1~7 號染色體上基因數目分別為1、10、3、11、0、3、7個。根據在染色體上的位置,將35 個基因命名為從到。其中,有3 對片段重復基因。CsCESA 蛋白的氨基酸殘基數目在205~1 146 之間,分子量在23.2~128.4 kD 之間,蛋白等電點(PI)在5.18~8.95 之間。利用在線網站Softberry 對基因家族蛋白進行亞細胞定位的預測,大部分家族成員定位于質膜上,少部分成員定位于高爾基體上(表2)。

圖1 黃瓜纖維素合成酶基因家族在染色體上的位置

表2 黃瓜纖維素合成酶基因全基因組鑒定與分子特征分析
2.2 黃瓜纖維素合成酶家族基因結構和進化分析
通過網站GSDS 2.0 對黃瓜CESA 家族基因進行結構分析,如圖2 所示:黃瓜CESA 基因家族外顯子數目差異明顯,外顯子數目為2~15 個不等。其中含有2 個外顯子,含有15 個外顯子,分別有2 個成員含有7、12 個和13個外顯子,分別有3 個成員含有3、5 個和14 個外顯子,分別有4 個成員含有4、8 個和9 個外顯子,分別有6 個成員含有6 個外顯子。
利用在線網站MEME 對CESA 基因家族進行motif 預測,數量設置為10 個,其余參數選擇默認。黃瓜的CESA 蛋白都含有該家族的保守結構域Cellulose-synt(PF03552),除了CsCESA5、CsCESA6、CsCESA20、CsCESA21 和CsCESA31分別含有3、4、3、6、5 個motif 外,其 余CsCESA 都含有7~10 個motif,這些motif 在基因家族中的分布表現出相似的模式,說明CESA 在進化過程中保守性較高(圖2)。10 個motif 長度在27~50 個氨基酸之間,平均為36.2。

圖2 黃瓜纖維素合成酶基因家族成員的系統進化和結構特征
為了分析黃瓜纖維素合成酶基因家族的進化關系,使用MEGA 7.0 軟件和在線軟件ITOL 對黃瓜(35)、擬南芥(10)、番茄(16)、西瓜(28)和甜瓜(33)中的CESA 基因家族蛋白序列進行系統進化樹構建。如圖3-A 所示,黃瓜、擬南芥和番茄CESA 基因家族成員被分為6 個group,其中group1、group2、group4 和group5 包含3 個物種的CESA 基因,group3 僅有黃瓜CESA 基因,group6包含黃瓜和番茄CESA 基因;如圖3-B 所示,黃瓜、西瓜和甜瓜CESA 基因家族成員均勻地分布在6 個分支上(group1~group6)。

圖3 不同作物纖維素合成酶基因家族的系統發育樹
為了進一步了解黃瓜CESA 基因家族的進化關系,通過軟件MCscanX 對黃瓜、擬南芥、番茄、西瓜和甜瓜中CESA 家族基因進行了物種間共線性分析。與、家族基因之間的同源性分析結果如圖4-A 所示,和家族基因之間有15 個同源基因對;和家族基因之間有12 個同源基因對;就單條染色體上的共線性基因分布而言,黃瓜5 號染色體上無共線性基因;2、3、4、6 號和7 號染色體上共線性基因數量較多,為4~6 條;1 號染色體上共線性基因較少,為1 條。與、家族基因之間的同源性分析結果如圖4-B 所示,和家族基因之間有19 個同源基因對;和家族基因之間有20 個同源基因對。就單條染色體上的共線性基因分布而言,黃瓜5 號染色體上無共線性基因;7 號染色體上共線性基因最多,為10 條;2、3、4 號和6 號染色體上共線性基因數量較多,為6~8 條;1 號染色體上共線性基因較少,為1 條。

圖4 不同作物纖維素合成酶基因家族的共線性分析
2.3 黃瓜纖維素合成酶家族基因啟動子中順式作用元件分析
為了更好地了解CESA 家族基因的調控機制,提取了CESA 家族中各基因的啟動子區域(ATG 起始位點上游2 000 bp 的堿基序列),利用在線網站 PlantCARE 對啟動子區域的順式作用元件進行預測和分析,然后使用TBtools 軟件對結果進行可視化(圖5)。這些順式作用元件包括光響應元件、防御和脅迫響應元件(干旱、低溫)、植物激素響應元件(茉莉酸甲酯、脫落酸、赤霉素、生長素和水楊酸)、空間基因表達元件(分生組織、胚乳、種子細胞)和生物過程中涉及到的相關的響應元件(代謝、晝夜節律和細胞周期等),推測CESA 基因家族成員可能通過調控上游轉錄因子參與整合多種反應。
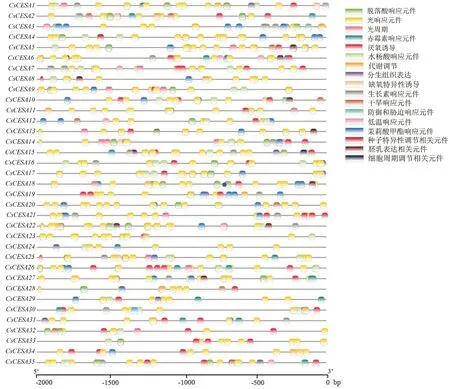
圖5 黃瓜纖維素合成酶基因家族成員啟動子序列順式作用元件分析
2.4 黃瓜纖維素合成酶家族基因的組織表達分析
利用從黃瓜基因組數據庫CuGenDB下載的10個黃瓜組織(根、莖、葉、受精子房、子房、雌花、未受精子房、雄花、卷須基部和卷須)的RNA-seq 數據,研究CESA 家族基因的組織表達情況,并將這些數據生成熱圖(圖6-A)。、和在各組織中均有較高的表達水平;、、、、、、、、、、和在各組織中均有一定的表達水平;在除去根、在除去雄花、和在除去根和雄花、在除去根、卷須和卷須基部之外的各組織中均有一定的表達;、、、、、、和在各組織中的表達量比較低。以上結果表明了其成員功能的分化和組織表達的特異性。

圖6 黃瓜纖維素合成酶家族基因的表達模式
2.5 黃瓜纖維素合成酶家族基因在非生物脅迫和激素處理下的表達分析
利用黃瓜基因組數據庫下載來自黃瓜幼苗在NaCl處理(PRJNA437579)以及低溫脅迫(PRJNA438923)下的RNA-seq數據。如圖6-B和6-C所示,大部分成員受NaCl和低溫的誘導表達,在NaCl處理下,、、、、、、、、和下調表達;、、、和表達量沒有明顯變化;其余基因均上調表達。低溫處理下,、、、、和上調表達;、、和在2h時上調表達;、和在2h和6h時上調表達;和在2h和12h時上調表達;、、、、和在6 h 和12 h 時上調表達;其余均下調表達或無明顯表達差異。對不同非生物脅迫處理有不同程度的響應,表明其可能在黃瓜植株響應非生物脅迫的過程中發揮作用。
選取黃瓜CESA 基因家族中的、、、、、、和進行非生物脅迫和植物生長調節劑處理下的表達分析,利用qRT-PCR檢測黃瓜品系9930在NaCl、ABA和MeJA不同時間梯度(0、3、6、12h和24h)處理下的表達譜,結果顯示:以上基因不同程度地受到鹽脅迫和植物激素ABA、MeJA的誘導。如圖7所示,在NaCl處理下,除和表達量下降以外,其余基因均受NaCl的誘導上調表達。外施激素ABA處理下,從0~24h,和上調表達;和下調表達;和表達量先升高后下降;表達量先下降后升高;表達量先下降后升高再降低。MeJA處理下,從0~24h,、、和上調表達;在3h上調表達,之后表達量下調;在6h上調表達,24h表達量下調;在12h表達量上調;在12h和24h表達量下調。8個基因均能響應非生物脅迫(包括低溫和鹽脅迫)和植物激素(ABA、MeJA)的誘導(圖6、7),其中、和表達量的峰值更高,表明這3個基因在黃瓜植株響應非生物脅迫的過程中更具有研究價值。

圖7 黃瓜纖維素合成酶基因在非生物脅迫(NaCl、ABA 和MeJA)誘導下的 qRT-PCR 表達分析
3 討論與結論
纖維素是植物細胞壁的主要成分,主要由36根β-1,4 糖苷鏈結晶而成的小微纖絲組成。纖維素合成酶(CESA)是一種與膜結合的糖基轉移酶,可以催化UDP-葡萄糖形成β-1,4 糖苷鏈合成纖維素(Festucci-buselli et al.,2007)。纖維素合成酶被稱之為“復合物的復合物”,其家族由多個成員組成(Somerville,2006)。近年來,人們對纖維素合成酶基因家族的研究集中于擬南芥、煙草等模式植物和棉花、楊樹、亞麻、毛竹等木本植物,黃瓜中纖維素合成酶基因家族的系統鑒定和分析還未見報道。從前人的研究中可以發現不同物種的CESA基因家族成員的數量存在很大差異,擬南芥、煙草、水稻、毛果楊及四倍體海島棉中分別鑒定出10、21、14、18個和37 個CESA(Richmond & Somerville,2000;Wang et al.,2010;袁紅梅 等,2016;Yuan et al.,2016;徐宗昌和孔英珍,2017)。
本試驗對黃瓜CESA 基因家族進行全基因組生物信息學分析,共鑒定出35 個CESA 基因家族成員,分布在除5 號染色體外的6 條染色體上,2、4、7 號染色體上分布較多,1、3、6 號染色體上分布較少,基因的不均勻分布可能與遺傳變異和物種進化相關。CESA 家族成員含有的motif 數量差異不大,其分布表現出相似的模式,說明CESA 基因家族成員在進化過程中具有較高的保守性。CESA家族成員含有的內含子、外顯子數量存在一定的差異,且所含有的氨基酸位點數量以及理化特征明顯不同,表明CsCESA 蛋白可能在其自身的微環境中發揮不同的作用。在系統發育分析中,黃瓜、擬南芥和番茄CESA 家族成員被分為6 個group,其中group1、group2、group4 和group5 包含3 個物種的CESA 基因,group3 僅有黃瓜CESA 基因,group6包含黃瓜和番茄CESA 基因,說明該家族的主要特征極有可能在雙子葉/單子葉分裂之前就已經產生并出現了分化;黃瓜、西瓜和甜瓜CESA 基因家族成員均勻地分布在6 個分支上,說明CESA 家族成員在葫蘆科植物之間具有較高的結構和功能相似性。前人研究表明,基因復制的兩種模式串聯基因復制和片段基因復制被認為是進化過程中基因家族擴張的主要推動力。發生拷貝數變異的基因可能存在表達和功能上的分化,并在獲得環境適應優勢的過程中起著重要作用(Cannon et al.,2004)。本試驗揭示了片段重復是負責CsCESA 基因家族進化的主要原因,經過物種內的共線性分析發現有3 對片段重復基因對,無串聯重復基因對(圖1)。
植物細胞壁除了人們常規認為的機械支持和物質運輸的功能外,還可以對各種逆境脅迫做出響應。植物細胞壁響應逆境脅迫是個復雜的生理生化過程,細胞壁的多種成分均參與響應的機制,表現為:細胞壁組分的含量和結構的變化,包括細胞壁多糖、細胞壁蛋白和酶以及質外體小分子物質,以上這些變化都是通過一系列相關酶的有序調節而實現的(Endler et al.,2015)。在植物中,CESA 基因家族成員具有較高的系統發育相似性,但是在不同物種中的時空表達模式不同,表達水平和表達種類具有明顯的組織特異性,對植物生長發育的調控也存在功能分化的現象(Hill et al.,2015;李秀云 等,2017;徐宗昌和孔英珍,2017)。前人研究結果表明,植物中纖維素含量的變化會引起植物對非生物和生物脅迫的響應,在擬南芥中,纖維素合成酶基因的突變導致纖維素含量減少,進而影響乙烯和茉莉酸甲酯信號途徑、病原微生物侵染、損傷誘導和干旱脅迫相關基因的表達(Ellis et al.,2002)。擬南芥的基因耐旱突變體可以通過調節脯氨酸、可溶性糖、ABA 含量和纖維素的轉化比例來調節植物的滲透能力,從而增強植物對干旱和滲透脅迫的耐受性(Chen et al.,2005)。本試驗中,低溫和鹽脅迫對基因有不同的調控作用,基因可能通過相同或者不同的調節機制來響應這些非生物脅迫。例如在低溫脅迫下上調表達,在鹽脅迫下下調表達,在低溫脅迫下下調表達,在鹽脅迫下上調表達,這些數據表明,和通過不同的機制對相同的脅迫做出反應。和在低溫和鹽脅迫下均下調表達,表明它們可能享有相同的調節機制或途徑。鹽脅迫處理下,、和未檢測到表達變化。說明CESA 基因能夠廣泛參與多種逆境脅迫的適應過程。
植物激素信號傳導網絡對于植物逆境響應至關重要。ABA 在植物對低溫和鹽脅迫的響應中起著至關重要的作用(Suzuki et al.,2016)。茉莉酸(JA)及其甲酯(MeJA)是內源性信號分子,在調節脅迫響應和植物發育中起著關鍵作用(Fung et al.,2004)。本試驗中,大多數基因受到ABA和MeJA 的誘導表達,尤其是,表達量上調的倍數最高。以上結果表明,基因家族可能在各種植物激素之間的串擾中起關鍵作用。本試驗中、和能夠響應非生物脅迫(包括低溫和鹽脅迫),此外,它們能夠受到ABA(一種整合各種脅迫信號并控制下游應答的激素)和MeJA(一種參與非生物脅迫的信號分子)的誘導表達,且這3 個基因的表達量相對較高。與和同源性最高的和的基因功能還未明確;與同源性最高的在種子粘液中的纖維素生成中起著重要的作用(Sullivan et al.,2011),并且在根組織中能受到干旱脅迫的誘導從而使表達量上調(Heidari et al.,2019)。因此,黃瓜中、和響應非生物脅迫的分子機制尚需進一步深入研究。
本試驗從9930_V2 版本基因組中鑒定到35 個CESA 基因,并對其進行了生物信息學分析和表達模式分析。在植物生長和發育過程中,基因具有明顯的組織表達差異性,協同調控了黃瓜的生長發育。基因的表達對非生物脅迫和植物激素處理的反應不同,其中、和均能響應非生物脅迫(包括低溫和鹽脅迫)和植物激素(ABA 和MeJA)的誘導,說明它們是應激反應中潛在的關鍵調控因子。本試驗結果有助于了解基因在黃瓜響應非生物脅迫過程中的作用,推動CESA 基因家族的研究與發展,接下來的試驗應進一步研究、和響應非生物脅迫的分子機制,減少非生物脅迫對黃瓜以及園藝植物生產的影響。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
