基于外殼蛋白基因的湖南煙草黃瓜花葉病毒遺傳多樣性分析
2022-04-04 12:55:33龔玉娟滕凱肖志鵬蔡海林周向平肖艷松劉天波唐前君
植物保護 2022年2期
關(guān)鍵詞:煙草
龔玉娟 滕凱 肖志鵬 蔡海林 周向平 肖艷松 劉天波 唐前君







摘要 為探明湖南煙草上發(fā)生的黃瓜花葉病毒Cucumber mosaic virus (CMV)的遺傳多樣性及分子進化特征,對來自湖南煙區(qū)的303份疑似感染病毒的煙草樣品進行檢測,分析CMV系統(tǒng)發(fā)育、遺傳變異和群體結(jié)構(gòu)等特征。結(jié)果表明:部分分離物的外殼蛋白(coat protein, CP)基因與NCBI上登錄的CMV分離物的一致性為86.34%~98.42%;系統(tǒng)發(fā)育分析發(fā)現(xiàn)湖南煙草CMV分離物屬ⅠB組,不同組間的分離物地理特征不明顯,無重組現(xiàn)象,進化的主要驅(qū)動力是負選擇;組間遺傳變異比較明顯,基因交流頻率較低,受到遺傳漂變影響,遺傳多樣性高,群體趨于擴張。研究結(jié)果為煙草抗CMV育種提供了理論依據(jù),對病害防治具有重要意義。
關(guān)鍵詞 煙草; 黃瓜花葉病毒; CP基因; 遺傳多樣性; 分子進化
中圖分類號: S435.72
文獻標識碼: A
DOI: 10.16688/j.zwbh.2021061
Abstract In order to investigate the genetic diversity and molecular evolution of Cucumber mosaic virus (CMV) on tobacco in Hunan province, 303 suspected diseased samples from tobacco-growing areas in Hunan were detected, and the phylogeny, genetic variation and population structure of CMV were analyzed. The coat protein (CP) genes of some samples were cloned and compared with CMV isolates registered on NCBI. The results showed that the sequence of CP from Hunan isolates shared identity of 86.34%-98.42% with isolate from NCBI. Phylogenetic analysis showed that CMV isolates from Hunan tobacco plants belonged toⅠB group. The geographical characteristics of these isolates were not distinct among different populations, and there was no recombination phenomenon. The main driving force for evolution was negative selection. The genetic variation among groups was obvious; the frequency of gene flow was low; the genetic diversity was high, and the population tended to expand. These results provide a theoretical basis for breeding tobacco for disease resistance to CMV and are of great significance for disease control.
Key words tobacco; Cucumber mosaic virus; CP gene; genetic diversity; molecular evolution
黃瓜花葉病毒Cucumber mosaic virus (CMV)是嚴重危害植物的病毒之一,其侵染煙草引發(fā)的病毒病在煙草上普遍發(fā)生,嚴重威脅我國煙葉生產(chǎn)[12]。培育抗病毒品種被認為是最有效的方法之一[3],但因CMV受寄主和地域影響而變異較頻繁,抗病品種抗性易減弱或喪失,使抗病育種難度加大。通過生物信息學(xué)技術(shù)和方法分析CMV不同分離物的遺傳變異和分子進化,對抗CMV的品種選育和病害精準防控有重要意義。
CMV株系眾多,寄主范圍廣泛,存在較顯著的遺傳多樣性[4]。CMV基因組有3條正義單鏈的RNA分子,編碼復(fù)制酶1a、2a、2b、移動蛋白(movement protein, MP)、外殼蛋白(coat protein, CP)[5]共5個蛋白。CMV CP基因序列保守性較高,常被應(yīng)用于病毒遺傳多樣性研究[6]。
根據(jù)癥狀、寄主范圍、致病性、抗原性、蚜傳特性以及核苷酸序列同源性的差異,CMV株系被分為亞組 Ⅰ和亞組 Ⅱ,在亞組 Ⅰ內(nèi)根據(jù)CMV RNA3的5′端和非編碼區(qū)序列的比對結(jié)果,亞組Ⅰ進一步分為ⅠA和ⅠB[7]。
宋麗云[8]、劉勇等[9]鑒定分析了我國云南、湖南和福建等地CMV分離物,發(fā)現(xiàn)云南和湖南煙草CMV以亞組Ⅰ為主,存在較明顯的遺傳多樣性。
趙雪君等[10]報道四川CMV分離物屬亞組ⅠB。
隨著產(chǎn)區(qū)調(diào)整、種植品種和耕作制度等變化,湖南煙草CMV群體變化和分子變異情況尚不清楚。因此,利用CMV CP基因序列開展群體遺傳多樣性和分子進化分析,探明CMV的遺傳變異機制很有必要。本研究對湖南煙草中的CMV分離物CP基因進行RT-PCR擴增和測序,分析CMV分離物的遺傳多樣性和分子進化特征,以期為CMV株系鑒定和進化機制研究提供參考,為CMV的抗病育種和防治提供理論依據(jù)。
1 材料與方法
1.1 試驗材料
樣品采集:2017年-2018年從湖南郴州、永州、長沙等5個煙區(qū)采集表型為花葉、皺縮、畸形及脈壞死等癥狀的煙草葉片樣品303份,樣品經(jīng)液氮速凍后于-80℃冰箱中保存?zhèn)溆谩?/p>
主要試劑:RNA提取試劑盒(EasyPure Plant RNA Kit),購自TransGen Biotech公司;cDNA合成試劑盒、Taq DNA聚合酶、dNTPs購自北京全式金生物有限公司;其他常規(guī)試劑為國產(chǎn)分析純,購自中國醫(yī)藥集團有限公司。
1.2 煙草總RNA的提取及檢測
取煙草葉片樣品1 g,使用RNA提取試劑盒提取總RNA,具體方法參照試劑盒說明書。以提取的總RNA為模板,反轉(zhuǎn)錄合成cDNA。用于RT-PCR擴增的引物[11]序列為CMVF: 5′-CGGATGCTAACTTTAGAGTCTTGT-3′和CMVR: 5′-GAATGCGTTGGTGCTCGAT-3′,由生工生物工程(上海)股份有限公司合成,目標片段長度約650 bp。PCR反應(yīng)參照Liu等[12]的方法,反應(yīng)體系(25 μL):10×buffer 2.5 μL,rTaq DNA聚合酶0.3 μL,10 mmol/L dNTPs 0.5 μL,10 μmol/L引物各0.5 μL,1 μmol/L模板DNA 0.5 μL,滅菌ddH2O補足至25 μL。擴增條件:95℃變性30 s,56℃退火50 s,72℃延伸110 s,30個循環(huán);72℃延伸10 min。PCR擴增產(chǎn)物送生工生物工程(上海)股份有限公司測序,計算檢出率。
1.3 重組分析
將獲得的湖南煙草CMV分離物CP基因序列進行整理,先利用RDP 4.0[13]軟件包中的MAXCHI、RDP、CHIMAERA等軟件檢測可能存在的重組位點,然后利用Simplot軟件驗證這些重組位點存在的可能性[14]。如檢測結(jié)果P<1.0×10-6,則表明CMV分離物存在重組的可能;如P>1.0×10-6,則表明CMV分離物不存在重組[15]。
1.4 系統(tǒng)發(fā)育分析
將測序獲得的陽性樣品序列采用DNAMAN進行同源性比對。參照比對結(jié)果選取采自湖南不同地區(qū)、差異相對較大的11個樣品,同時從NCBI上下載32個來源不同國家或地區(qū)的CMV分離物 CP基因序列進行系統(tǒng)發(fā)育分析,采用MEGA 5.0中的Clustal W軟件進行序列比對,樣品編號或登錄號和來源地區(qū)或報道國家見表1。采用最大似然法構(gòu)建系統(tǒng)進化樹(自展值1 000),以與CMV同屬的花生矮化病毒病Peanut stunt virus (PSV) 作為外群。依據(jù)Kimura two-parameter和DayhoffPAM 250 matrix計算分析核苷酸和氨基酸相似性[16]。
1.5 遺傳差異和基因交流
利用DnaSP 5.0軟件計算群體間遺傳差異與基因交流頻率。統(tǒng)計KS、Z和Snn 3個參數(shù),Snn指近鄰統(tǒng)計(nearest-neighbor statistic),計算兩個或兩個以上的地區(qū)樣品近鄰序列在相同地域的地理學(xué)空間上出現(xiàn)的頻率;
Ks和Z是指以序列為基礎(chǔ)的數(shù)據(jù)統(tǒng)計分析[17],
Ks由不同序列的數(shù)量所決定,Z是秩統(tǒng)計量(rank statistic)。如果Z和 KS統(tǒng)計值很小,P<0.05(兩個亞群之間沒有遺傳分化),則拒絕無遺傳差異的假說[18]。利用DnaSP 5.0 軟件計算Fst和Nm,依據(jù)Fst和Nm大小評估基因交流情況,F(xiàn)st是指群體間遺傳差異的構(gòu)成,表示群體間等位基因頻率的標準差異;Nm是指有效大小的單個群體數(shù)量以及它們之間的遷移率。如果Fst的絕對值大于0.33,說明基因交流不頻繁,反之,則交流頻繁[19];如果Nm大于1則說明基因漂變在群體間很容易發(fā)生;如果Nm小于1,則說明群體發(fā)生遺傳差異的主要原因是基因交流[20]。
1.6 選擇壓力分析
選擇壓力評價采用MEGA 5.0 中Pamilo-Bianchi-Li法,分別計算非同義突變(dN)和同義突變(dS)的值,利用dN/dS值預(yù)測各個基因所承受的選擇壓力[21]。若dN/dS=1則說明該組分離物處于中性選擇;若dN/dS<1則說明該組分離物處于負選擇壓力作用下;若dN/dS>1則說明該組分離物處于正向選擇作用下。
1.7 突變位點和保守區(qū)域分析
利用DnaSP 5.0 軟件分析CMV CP基因突變位點和保守區(qū)域。
1.8 群體結(jié)構(gòu)分析
群體結(jié)構(gòu)利用核苷酸多樣性與單體型多樣性評價,采用DnaSP 5.0計算核苷酸多樣性與單體型多樣性的差異值[22]。核苷酸多樣性差異值以0.005為臨界值,單體型多樣性差異值以0.5為臨界值,二者的值越大,表示群體的多樣性程度越高。
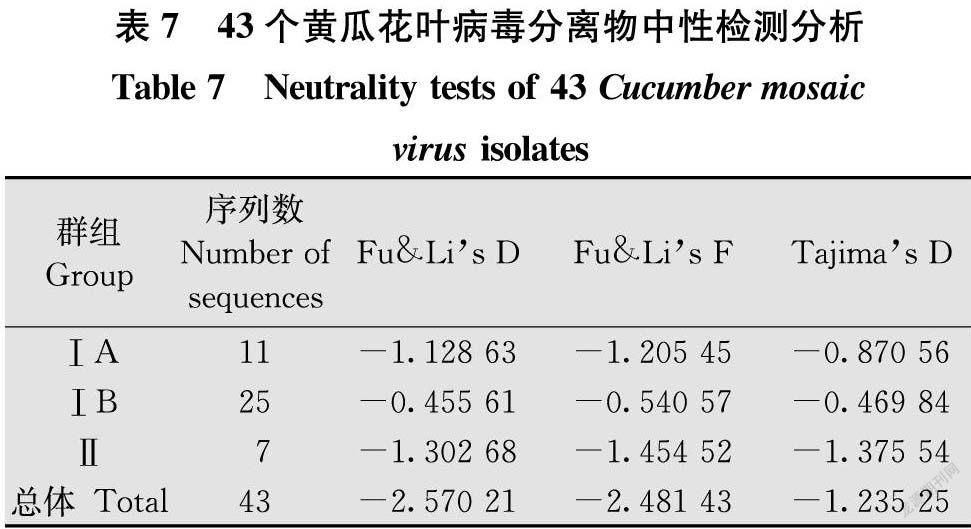
1.9 中性檢驗分析
中性檢驗分析基于Fu & Li’s D、Fu & Li’s F和Tajima’s D值評價,若上述3個參數(shù)均小于零,說明該群體多樣性較低,群體存在選擇壓力或呈擴張趨勢;若
這3個參數(shù)值大于零,說明群體數(shù)量趨于減少,群體處于收縮態(tài)勢[23]。
2 結(jié)果與分析
2.1 湖南煙區(qū)CMV檢測
對303份有花葉、皺縮、葉緣上卷等癥狀的煙草樣品進行PCR擴增,有192份樣品得到大小約為650 bp的片段(圖1),條帶大小符合預(yù)期,陽性檢出率63.37%(表2)。對PCR產(chǎn)物(CP基因)進行測序,并與NCBI上登錄的CMV分離物CP基因序列進行比對,結(jié)果顯示湖南煙區(qū)CMV分離物與NCBI上登錄的CMV分離物(登錄號KU976486、EF424776、HQ844075等)的一致性為86.34%~98.42%。
2.2 重組分析
對得到的CMV分離物CP基因序列進行重組分析,結(jié)果表明,P>1.0×10-6,CMV分離物CP基因無重組現(xiàn)象或重組現(xiàn)象不明顯。利用Simplot進一步確認,結(jié)果顯示不存在明顯重組事件。
2.3 系統(tǒng)發(fā)育分析
同源性比對顯示,192份陽性樣品序列一致性為93.35%。選取 11個湖南不同地區(qū)CMV分離物與32個已報道的不同國家或地區(qū)的CMV分離物 CP基因序列構(gòu)建系統(tǒng)發(fā)育樹(圖2),結(jié)果發(fā)現(xiàn)CMV不同分離物形成ⅠA、ⅠB和Ⅱ3個組,ⅠA 組以亞洲(中國、日本、韓國)分離物為主,少部分為美國、匈牙利和西班牙分離物,ⅠB組以中國(湖南)分離物為主,其他為意大利、西班牙、印度尼西亞和菲律賓分離物,Ⅱ組主要為日本、南非、澳大利亞和美國分離物,不同組間的分離物地理特征不明顯。湖南CMV分離物屬于亞組ⅠB,不同地區(qū)的分離物在系統(tǒng)發(fā)育樹中聚集分布,來自長沙寧鄉(xiāng)(CS-NX)、湘西龍山(XX-LS)、郴州永興(CZ-YX)的3個分離物稍微分散,其他地區(qū)分離物地理特征不明顯,遺傳距離較近,一致性較高,核苷酸序列的一致性為93.8%~96.6%,氨基酸的一致性為95.9%~96.9%。
2.4 遺傳差異和基因交流
系統(tǒng)發(fā)育分析將43個CMV分離物分成ⅠA、ⅠB和Ⅱ 3個組,對這3個組進行遺傳差異分析,結(jié)果顯示不同組間的KS和Z值較小,P值也較小,接近于0,遠小于0.001,表明CMV在不同組間的遺傳差異比較明顯(表3)。基因交流分析結(jié)果顯示,CMV不同組間的Fst值均大于0.33,表明CMV在不同組間基因交流頻率不高。CMV不同組間Nm均小于1,表明群體可能受到遺傳漂變影響。
2.5 選擇壓力分析
利用dN和dS預(yù)測不同群組分離物非同義突變率和同義突變率,分別計算dN和dS值,估量CP基因序列的選擇壓程度(表4)。結(jié)果顯示dN值小于dS值,dN/dS均小于1,表明CMV不同群體受負選擇壓力較強,負選擇可能是CMV進化的主要驅(qū)動力。
2.6 突變位點和保守區(qū)域分析
突變位點分析發(fā)現(xiàn),CMV CP基因有可變位點260個,單突變位點128個,其中2個變異型位點119個,3個變異型位點9個。保守區(qū)域分析發(fā)現(xiàn)(表5),CMV CP基因保守區(qū)域共有3個,保守性為0.54,表明 CMV CP基因突變率很低,比較保守。
2.7 群體結(jié)構(gòu)分析
群體結(jié)構(gòu)分析結(jié)果表明(表6),ⅠA、ⅠB和Ⅱ 3個組的核苷酸多樣性差異值分別為0.021 55、0.055 47和0.013 96,差異值均大于0.005,其中ⅠA和ⅠB組的核苷酸多樣性差異值均大于組Ⅱ,表明ⅠA和ⅠB組遺傳多樣性高于組Ⅱ。3個組的單體型多樣性差異值接近,均大于0.5,接近1,表現(xiàn)出較高的單體型多樣性。總體來看,核苷酸多樣性總體差異值為0.110 32(大于0.005),單體型多樣性差異值為0.998(大于0.5),說明3個組總體具有較高的核苷酸多樣性和單體型多樣性。
2.8 中性檢測分析
中性檢測結(jié)果顯示,3個組CP基因序列的Fu & Li’s D、Fu & Li’s F和Tajima’s D 3個參數(shù)檢驗值均小于0(表7),總體值也均為負值,表明 CMV群體處于擴張趨勢,群體在不斷擴張。
3 討論
CMV寄主范圍廣、地域適應(yīng)性強,在與寄主植物互作過程中,相互協(xié)同進化,易發(fā)生變異。開展CMV群體遺傳多樣性研究,探明CMV的分子進化特征,對科學(xué)精準防控CMV意義重大。
據(jù)報道我國CMV分離物多為亞組Ⅰ(ⅠA、ⅠB),而亞組Ⅱ較少發(fā)現(xiàn)[24]。如宋麗云[8]對全國32個CMV分離物進行鑒定分析發(fā)現(xiàn)有18個屬于亞組Ⅰ。劉勇等[9]鑒定發(fā)現(xiàn)湖南煙區(qū)CMV株系主要為亞組Ⅰ,占比94.1%,但未細分出ⅠA和ⅠB。本研究對湖南煙草上CMV分離物的CP基因進行了序列分析并構(gòu)建了系統(tǒng)發(fā)育樹,結(jié)果表明湖南CMV分離物均屬于亞組ⅠB,與劉勇、宋麗云等的研究結(jié)果一致。劉勇等[9]在云南和貴州煙草發(fā)現(xiàn)了少量的病毒樣品屬亞組Ⅱ,但本研究未發(fā)現(xiàn)湖南煙草上有亞組Ⅱ分離物,這可能與煙草品種差異或采樣的點還不全面有關(guān)。CMV的發(fā)生、進化和變異與不同寄主、地理位置、生長環(huán)境等因素有關(guān),而且不同CMV亞組可能與其致病性強弱有關(guān)[25],樣品陽性檢出率是否能夠反映CMV實際發(fā)生率,這些問題還有待進一步探究。另外,本研究僅基于CP基因進行系統(tǒng)分析,也有可能存在誤判,還需基于不同功能基因進一步分析。鑒于我國煙草上主要以CMV亞組ⅠB株系為主,故煙草CMV的抗病育種與防治應(yīng)以亞組ⅠB株系為主要靶標,研究結(jié)果為煙草CMV的抗病品種選育提供了理論依據(jù)和支撐。
病毒豐富的遺傳多樣性主要來自基因重組、突變、基因流、遺傳漂變和自然選擇等因素長期相互作用[26]。推動病毒不斷進化的主要動力是基因重組[27],它增強病毒適應(yīng)新寄主和新環(huán)境的能力,進而擴大病毒的寄主范圍[28]。重組分析發(fā)現(xiàn)湖南煙草CMV CP基因無重組現(xiàn)象或重組現(xiàn)象不明顯,推測可能因為CMV不存在重組,以致煙草上CMV的發(fā)生一直比較穩(wěn)定。遺傳漂變對病毒群體突變的頻率有重要作用,CMV存在ⅠA和ⅠB兩個相對獨立的群體,遺傳變異顯著,ⅠA和ⅠB之間的基因交流頻率較低。中國煙草CMV Ⅰ A主要與東亞的日本、韓國分離物親緣關(guān)系較近,相互交流頻率高,而Ⅰ B主要與東南亞的菲律賓、印度尼西亞交流較頻繁,與屬于亞組Ⅱ的歐洲、美國分離物之間交流頻率較低。推測中國煙草CMV起源有可能與煙草傳入中國的路徑相似,煙草攜帶CMV由日本和菲律賓南北兩條路徑傳入中國沿海,再傳入中國內(nèi)地[29],在現(xiàn)代煙草種植過程中,中國內(nèi)地不同地區(qū)間攜帶CMV的煙草種子交流頻繁,導(dǎo)致CMV群體間同時存在遺傳差異和基因交流。
突變是驅(qū)動CMV進化的主要作用力之一[30]。選擇壓力和突變分析表明負選擇是CMV進化的主要驅(qū)動力,CMV CP基因序列變異較少,
遺傳結(jié)構(gòu)相對比較穩(wěn)定,這與趙雪君等[10]的研究結(jié)果相一致。遺傳多樣性參數(shù)(核苷酸多樣性和單體型多樣性)分析結(jié)果表明CMV群體遺傳多樣性較高,群組ⅠA和ⅠB遺傳多樣性高于Ⅱ,表明兩個群體相對穩(wěn)定且長期演化,但群體之間呈現(xiàn)一定程度的分化,
整體處于擴張趨勢。群體內(nèi)這種高遺傳多樣性的形成是一個長期過程,推測可能由于中國煙草種植區(qū)之間的種苗調(diào)運頻率較高,且CMV為重要的蚜傳病毒,經(jīng)蚜蟲傳播導(dǎo)致不同地區(qū)和寄主間的CMV交流頻繁,易發(fā)生突變和自然選擇,從而造成不同群體的遺傳多樣性較高。
4 結(jié)論
對來自湖南煙草CMV 的CP基因進行了RT-PCR和測序,分析了湖南煙草CMV遺傳多樣性及分子進化機制,結(jié)果表明湖南煙草CMV屬亞組ⅠB,群體間地理相關(guān)性不明顯,無明顯重組現(xiàn)象,遺傳變異明顯,基因交流頻率低,遺傳多樣性較高,負選擇是驅(qū)動進化的主要作用力,種群處于擴張趨勢。本研究開展了CMV系統(tǒng)進化和遺傳多樣性分析,揭示了CMV分子變異進化特征,為CMV的抗病品種選育和病害精準防治提供了理論依據(jù)和科學(xué)指導(dǎo)。
參考文獻
[1] 朱賢朝,王彥亭,王智發(fā).中國煙草病害[M].北京:中國農(nóng)業(yè)出版社,2002.
[2] 王鳳龍,周義和,任廣偉. 中國煙草病害圖鑒[M].北京:中國農(nóng)業(yè)出版社,2019.
[3] 王鳳龍. 煙草病毒病綜合防治技術(shù)[J].煙草科技, 2002(4): 4345.
[4] SCHOLTHOF K B, ADKINS S, CZOSNEK H, et al. Top 10 plant viruses in molecular plant pathology. Molecular Plant Pathology, 2011,12(9):938954.
[5] HELLWALD K H, PALUKAITIS P. Nucleotide sequence and infectivity of Cucumber mosaic cucumovirus (strain K) RNA2 involved in breakage of replicase-mediated resistance in tobacco [J]. Journal of General Virology, 1994, 75(8):2121–2125.
[6] ROOSSINCK M J, ZHANG L, HELLWALD K H. Rearrangements in the 5′nontranslated region and phylogenetic analyses of Cucumber mosaic virus RNA 3 indicate radial evolution of three subgroups [J]. Journal of Virology, 1999, 73(8):67526758.
[7] SAFAEIZADEH M, SAIDI A, PALUKAITIS P, et al. Molecular characterization of Cucumber mosaic virus isolates infecting tomato in Hamedan and Tehran provinces of Iran [J]. Acta Virologica, 2015, 59(2): 174178.
[8] 宋麗云. 我國煙草TMV和CMV種群結(jié)構(gòu)遺傳分析[D]. 北京: 中國農(nóng)業(yè)科學(xué)院, 2012.
[9] 劉勇, 莫笑晗, 余清, 等. 云南·福建·湖南煙區(qū)煙草花葉病主要病毒種類檢測及黃瓜花葉病毒亞組鑒定[J]. 植物病理學(xué)報, 2006, 36(4): 310313.
[10]趙雪君, 劉世超, 李斌, 等. 四川煙區(qū)CMV和PVY株系分化研究[J]. 西南大學(xué)學(xué)報(自然科學(xué)版), 2017, 39(3): 816.
[11]楊金廣, 張帥, 申莉莉, 等. 煙草中TMV、CMV和PVY多重RT-PCR檢測體系的建立與應(yīng)用[J].中國煙草學(xué)報, 2010,16(4): 8388.
[12]LIU Yuanyuan, YU Shanlin, LAN Yufei, et al. Molecular variability of five Cucumber mosaic virus isolates from China [J]. Acta Virologica, 2009, 53(2):8997.
[13]MARTIN D P. RDP4: Detection and analysis of recombination patterns in virus genomes [J/OL]. Virus Evolution, 2015, 1(1): vev003. DOI: 10.1093/ve/vev003.
[14]匡云波, 葉煒, 李金輝, 等. 太子參蠶豆萎蔫病毒2號CP基因的遺傳多樣性及分子進化分析[J]. 植物病理學(xué)報, 2017, 47(4): 470478.
[15]TAN Zhongyang, WADA Y, CHEN Jishuang, et al. Inter- and intralineage recombinants are common in natural populations of Turnip mosaic virus [J]. Journal of General Virology, 2004, 85(Pt9): 26832696.
[16]DAYHOFF M O, BARKER W C, HUNT L T, et al. Establishing homologies in protein sequences [J]. Methods in Enzymology,1983, 91: 524545.
[17]HUDSON R R. A new statistic for detecting genetic differentiation [J]. Genetics,2000, 155: 20112014.
[18]賀振,李文鳳,李世訪.云南蔗區(qū)甘蔗線條花葉病毒分離物NIa基因形成新簇[J].微生物學(xué)報,2016,56(11):18021810.
[19]劉嘉裕, 戴良英, 劉勇, 等. 長沙地區(qū)南瓜的小西葫蘆黃花葉病毒檢測及其分子進化分析[J]. 湖南農(nóng)業(yè)大學(xué)學(xué)報(自然科學(xué)版), 2017, 43(3): 274281.
[20]TOMITAKA Y, OHSHIMA K. A phylogeographical study of the Turnip mosaic virus population in East Asia reveals an ‘emergent’ lineage in Japan [J]. Molecular Ecology, 2006, 15(14): 44374457.
[21]POSADA D. jModelTest: phylogenetic model averaging [J]. Molecular Biology and Evolution, 2008, 25(7): 12531256.
[22]賀振, 李文鳳, 張志想, 等. 甘蔗線條花葉病毒HC-Pro基因的分子變異分析[J]. 植物保護, 2017, 43(2): 2936.
[23]FU Y X, LI W H. Statistical tests of neutrality of mutations [J]. Genetics, 1993, 133(3): 693709.
[24]徐平東, 周仲駒, 林奇英, 等. 黃瓜花葉病毒亞組Ⅰ和Ⅱ分離物外殼蛋白基因的序列分析與比較[J]. 病毒學(xué)報, 1999, 15(2): 165171.
[25]覃瑞, 程旺元. 黃瓜花葉病毒研究進展[J]. 中南民族大學(xué)學(xué)報(自然科學(xué)版), 2004, 23(2): 3338.
[26]宋震,劉科宏,楊方云,唐科志,李中安,周常勇.柑橘碎葉病毒外殼蛋白基因的克隆和序列分析[J].中國農(nóng)業(yè)科學(xué),2009,42(10):37413748.
[27]陳玉珍, 譚新球, 朱春暉, 等. 我國部分地區(qū)常見農(nóng)作物上黃瓜花葉病毒分離物核酸多樣性分析[J]. 植物保護學(xué)報, 2016, 43(3): 427433.
[28]WOROBEY M, HOLMES E C. Evolutionary aspects of recombination in RNA viruses [J]. Journal of General Virology, 1999, 80(10): 25352543.
[29]劉天波, 周志成, 彭曙光, 等. 基于全基因組編碼區(qū)序列的煙草花葉病毒分子進化分析[J]. 植物保護, 2017, 43(5): 8792.
[30]GARCAARENAL F, FRAILE A, MALPICA J M. Variability and genetic structure of plant virus populations [J]. Annual Review of Phytopathology, 2001, 39(1):157186.
(責(zé)任編輯:楊明麗)
猜你喜歡
奧秘(創(chuàng)新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學(xué)報(2019年5期)2019-11-14 07:54:12
首都公共衛(wèi)生(2019年5期)2019-05-21 01:08:34
浙江中西醫(yī)結(jié)合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當(dāng)代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學(xué)林(2013年2期)2013-11-12 12:58:54
中國煙草學(xué)報(2012年5期)2012-04-12 06:21:18
