多異瓢蟲不同發育階段過冷卻點及體液結冰點的測定
2022-04-04 12:55:33羿超群謝佼昕劉曉旭唐浩宇劉廷輝張永軍
植物保護 2022年2期
羿超群 謝佼昕 劉曉旭 唐浩宇 劉廷輝 張永軍
摘要 多異瓢蟲Hippodamia variegata是新疆棉田優勢捕食性天敵。為明確不同發育階段的多異瓢蟲的耐寒能力,本研究利用過冷卻點儀對多異瓢蟲的3齡、4齡幼蟲,蛹,2日齡雌、雄成蟲分別進行了過冷卻點和體液結冰點的測定,并對其頻次分布進行了分析。結果表明:蛹的過冷卻點(-21.05±0.75)℃和體液結冰點(-17.45±0.89)℃最低;4齡幼蟲的過冷卻點(-15.02±0.55)℃和體液結冰點(-11.80±0.64)℃次之;2日齡雄成蟲的過冷卻點(-14.74±0.42)℃和體液結冰點(-11.26±0.42)℃與3齡幼蟲的過冷卻點(-14.07±0.68)℃和體液結冰點(-11.25±0.61)℃差異不顯著;2日齡雌成蟲的過冷卻點(-12.97±0.40)℃和體液結冰點(-9.65±0.32)℃最高。結果顯示,多異瓢蟲蛹的耐寒性最強,2日齡雄成蟲的耐寒性高于雌成蟲,2日齡雌成蟲的耐寒性最弱。
關鍵詞 多異瓢蟲; 過冷卻點; 結冰點; 抗寒性
中圖分類號: S476.2
文獻標識碼: A
DOI: 10.16688/j.zwbh.2021273
Abstract Hippodamia variegata is the dominant predatory natural enemy of cotton fields in Xinjiang.In order to clarify the cold tolerance of Hippodamia variegata at different developmental stages, we determined the supercooling point and freezing points of hymolymph in 3rd-instar, 4th-instar larvae, pupae, 2-day-old female and male adults by using the supercooling point instrument, and then analyzed their frequency distribution. The results showed that the supercooling point (-21.05±0.75)℃ and freezing point (-17.45±0.89)℃ of pupae were the lowest among the larvae, pupae and adults, followed by the supercooling point (-15.02±0.55)℃ and the freezing point (-11.80±0.64)℃ of the 4th-instar larvae. The supercooling point (-14.74±0.42)℃ and freezing point (-11.26±0.42)℃ of 2-day-old male adults were not significant different with those of the 3rd-instar larvae (-14.07±0.68)℃ and (-11.25±0.61)℃, respectively. The 2-day-old female adults had the highest supercooling point (-12.97±0.40)℃ and freezing point (-9.65±0.32)℃. Our data indicated that the pupa of H.variegata had the strongest cold tolerance, and the 2-day-old male adults had higher cold tolerance than that of the female adult, and the 2-day-old female adults showed the weakest cold tolerance among the larvae, pupae and adults.
Key words Hippodamia variegata; supercooling point; freezing point; cold hardiness
多異瓢蟲Hippodamia variegata(Goeze)屬鞘翅目Coleoptera瓢甲科Coccinellidae,為完全變態昆蟲,在個體發育中經歷卵、幼蟲、蛹和成蟲4個階段。其幼蟲和成蟲均可捕食棉蚜 Aphis gossypii、白毛蚜Chaitophorus populialbae、豌豆蚜Acyrthosiphon pisum和桃蚜Myzus persicae等蚜蟲[14],以及棉鈴蟲Helicoverpa armigera、草地貪夜蛾Spodoptera frugiperda等鱗翅目昆蟲的卵[56],對葡萄斑葉蟬Erythroneura apicalis也有一定的捕食作用[7]。野外調查發現,多異瓢蟲以成蟲越冬,以內蒙古通遼市為例,多異瓢蟲多聚集在向陽的土縫、樹皮裂縫或田際雜草叢中度過低溫期[8],翌年4月下旬到5月上旬陸續到靠近地面的雜草及作物上取食蚜蟲等害蟲[9]。
溫度對多異瓢蟲各發育階段的歷期、存活率、繁殖力以及分布范圍等均有影響,是重要的環境因素之一[1011]。昆蟲屬于變溫動物,其體內一系列的生理生化反應與溫度的變化密切相關,溫度過高或者過低都會對昆蟲的生長、交配、繁殖及種群動態產生不同程度的不利影響[11]。在昆蟲的耐寒性研究中,過冷卻點(supercooling point)和結冰點(freezing point)是反映昆蟲耐寒性強弱的主要指標[12],多數不耐凍害的昆蟲,在長時間甚至短暫暴露于超過其過冷卻點的溫度下會死亡。已有學者對七星瓢蟲Coccinella septempunctata、異色瓢蟲Harmonia axyridis、草地貪夜蛾、勞氏黏蟲Mythimna loreyi、鉤紋皮蠹Dermestes ater等昆蟲的過冷卻點及耐寒性開展了研究[1317]。多異瓢蟲是新疆棉田的優勢天敵種群,其捕食范圍廣、捕食量大、對溫濕度的適應性強,對棉田蚜蟲的控制起到了非常重要的作用[18]。目前,對多異瓢蟲的研究多集中于其耐饑餓能力[19]、除草劑對其的安全性評價[20]、捕食能力[21]等方面,而關于多異瓢蟲抗寒能力方面的研究鮮見報道。基于此,為深入解析多異瓢蟲的耐寒能力,在室內測定了多異瓢蟲不同發育階段過冷卻點及體液結冰點,以期為多異瓢蟲的自然保護和利用提供數據支撐。
1 材料與方法
1.1 供試蟲源
供試多異瓢蟲、桃蚜和青豌豆Pisum sativum種子均由中國農業科學院植物保護研究所廊坊科研中試基地提供。在溫室進行青豌豆育苗,當豆苗株高2 cm,真葉剛剛長出時,移入養蟲罐并接入桃蚜,置于人工氣候箱中繁殖蚜蟲。多異瓢蟲的蛹放于高15 cm,直徑10 cm的圓柱形養蟲罐內,置于人工氣候箱中,待成蟲羽化,用桃蚜飼養成蟲,成蟲交配后,每日將養蟲罐內所有卵取出放進新的養蟲罐,標記好日期,待卵孵化為幼蟲后用蚜蟲飼喂,進行擴繁。氣候箱溫度為(25±1)℃,濕度為(65±5)%,光周期為L∥D=16 h∥8 h,光照強度為16 000 lx。
1.2 儀器設備
智能人工氣候箱(RXZ型),寧波市科技園區新江南儀器有限公司;加濕器(YC-D205型),北京亞都環保科技有限公司;智能昆蟲過冷卻點測定儀(SUN-V型),北京鵬程電子科技中心;高低溫試驗箱(BPH-606C型),上海一恒科技有限公司;電暖氣(NY 2011-16JW型),廣東美的環境電器制造有限公司;溫濕度計(HTC-1型),武強縣立誠儀器儀表廠。
1.3 多異瓢蟲過冷卻點和體液結冰點的測定
分別挑選多異瓢蟲3齡幼蟲、4齡幼蟲、2日齡蛹、2日齡雌、雄成蟲進行過冷卻點和體液結冰點的測定。先將試蟲放入0.2 mL離心管中,再將過冷卻點測定儀的熱敏電阻感溫探頭上部用脫脂棉裹緊,脫脂棉的厚度為放進0.2 mL離心管后不松動為宜,且脫脂棉不能遮擋電阻感溫探頭,處理完成后,將電阻感溫探頭伸進裝有試蟲的離心管中,使探頭與試蟲充分接觸。將連接、固定好的離心管放入高低溫試驗箱中,最低溫度設置為-40℃,以1℃/min的速率由室溫開始下降到最低溫,每秒記錄1次試蟲的體溫變化,通過過冷卻點測定儀系統軟件(V 1.3)分析得到體溫變化曲線,當試驗箱溫度降到某一溫度值時,試蟲體液開始結冰,體溫不再下降,反而會釋放出自身熱量,體溫開始回升,此時拐點的溫度值為試蟲的過冷卻點,當試驗箱溫度繼續下降時,曲線會出現第二個拐點,此時溫度值為試蟲的體液結冰點[22]。
1.4 數據分析
運用IBM SPSS Statistics 21軟件計算試驗數據的平均值和標準誤,對不同發育階段試蟲的過冷卻點和體液結冰點進行正態性檢驗和方差齊性檢驗。經方差分析確定差異顯著性后,運用Duncan氏新復極差法比較各發育階段試蟲過冷卻點和體液結冰點的差異,利用單一樣本Kolmogorov-Smirnov檢驗法檢驗各階段試蟲過冷卻點和體液結冰點的頻次分布特征,運用Origin 2018繪制頻次分布圖。
2 結果與分析
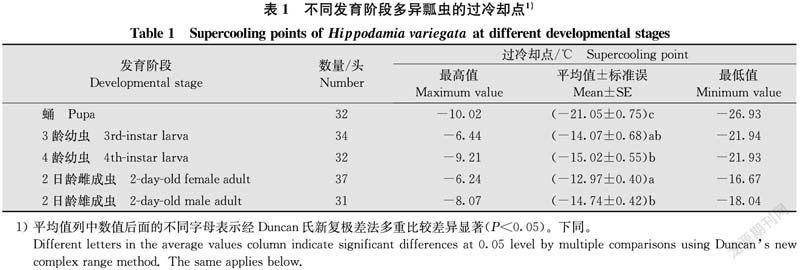
2.1 不同發育階段多異瓢蟲過冷卻點的比較
不同發育階段多異瓢蟲的過冷卻點存在顯著差異(df=4,161,F=30.25,P<0.05,表1),由低到高為:蛹(-21.05±0.75)℃<4齡幼蟲(-15.02±0.55)℃<2日齡雄成蟲(-14.74±0.42)℃<3齡幼蟲(-14.07±0.68)℃<2日齡雌成蟲(-12.97±0.40)℃。蛹期過冷卻點顯著低于幼蟲和成蟲(P<0.05),3齡、4齡幼蟲的過冷卻點差異不顯著(P>0.05),2日齡雌、雄成蟲的過冷卻點差異顯著(P<0.05)。結果表明,多異瓢蟲蛹的過冷卻點最低,雄成蟲的過冷卻點低于雌成蟲。通過過冷卻點的比較,蛹的耐寒性最強,2日齡雌成蟲的耐寒性最弱。
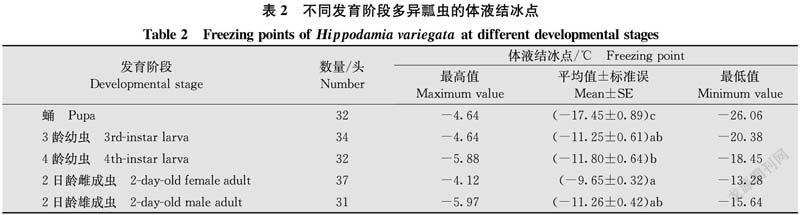
2.2 不同發育階段多異瓢蟲體液結冰點的比較
不同發育階段多異瓢蟲的體液結冰點存在顯著差異(df=4,161,F=25.52,P<0.05)(表2),由低到高為:蛹(-17.45±0.89)℃<4齡幼蟲(-11.80±0.64)℃<2日齡雄成蟲(-11.26±0.42)℃<3齡幼蟲(-11.25±0.61)℃<2日齡雌成蟲(-9.65±0.32)℃。蛹的體液結冰點顯著低于幼蟲和成蟲,3齡、4齡幼蟲的體液結冰點差異不顯著,2日齡雌、雄成蟲的體液結冰點無顯著差異。結果表明,多異瓢蟲蛹的體液結冰點最低,其耐寒性最強,2日齡雌成蟲的耐寒性最弱。
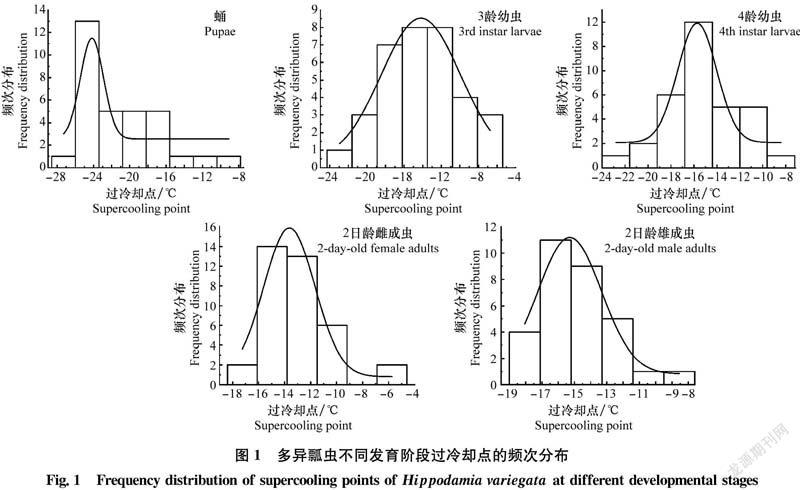
2.3 不同發育階段多異瓢蟲過冷卻點的頻次分布
不同發育階段多異瓢蟲的不同個體過冷卻點存在不同程度的差異,符合正態分布(P<0.05)(圖1)。不同發育階段不同個體過冷卻點的分布范圍有一定的寬度,過冷卻點最低值出現在蛹的個體中,為-26.93℃;最高值出現在2日齡雌成蟲的個體中,為-6.24℃。蛹的過冷卻點分布范圍最寬,為-26.93~-10.02℃;雄成蟲的過冷卻點分布范圍最窄,為-18.04~-8.07℃。其范圍寬度由大到小依次為:蛹(16.91℃)>3齡幼蟲(15.5℃)>4齡幼蟲(12.72℃)>2日齡雌成蟲(10.43℃)>2日齡雄成蟲(9.97℃)。蛹對低溫的上下浮動適應能力最強,且能適應低溫浮動的幅度較大,頻次分布多集中在-26~-25℃之間;雄成蟲對低溫上下浮動的適應能力最弱,且能適應低溫浮動的幅度較小,頻次分布多集中在-17~-13℃。
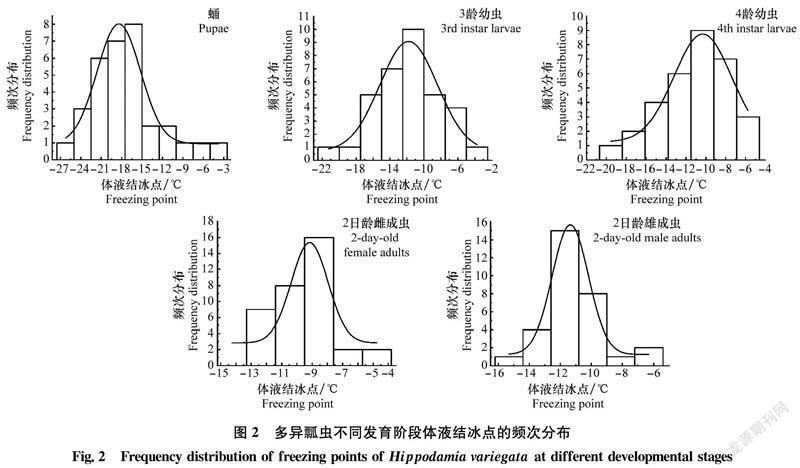
2.4 不同發育階段多異瓢蟲體液結冰點的頻次分布
不同發育階段多異瓢蟲體液結冰點存在不同程度的差異,符合正態分布(P<0.05)(圖2)。不同發育階段不同個體的體液結冰點分布范圍有一定的寬度,最低值出現在蛹的個體中,為-26.06℃;最高值出現在2日齡雌成蟲的個體中,為-4.12℃。蛹的體液結冰點分布范圍最寬,為-26.06~-4.64℃;雌成蟲的體液結冰點分布范圍最窄,為-13.28~-4.12℃。體液結冰點頻次分布范圍寬度由大到小排列順序為:蛹(21.42℃)>3齡幼蟲(15.74℃)>4齡幼蟲(12.57℃)>2日齡雄成蟲(9.67℃)>2日齡雌成蟲(9.16℃)。蛹能接受的低溫變化以及對低溫的抵抗能力最強,頻次分布多集中在-25~-15℃;從體液結冰點頻次分布圖來看,雌成蟲體液結冰點要高于雄成蟲,意味著在溫度到達過冷卻點后,雌成蟲抵抗低溫能力比雄成蟲要弱。
3 討論
在已有的昆蟲耐寒性研究中,過冷卻點和體液結冰點是評價其抗寒性的重要依據,Asahina認為過冷卻點是昆蟲所能忍受的極限低溫[23]。也有研究表明過冷卻點和體液結冰點與昆蟲的滯育和越冬密切相關,昆蟲通常為了適應低溫的到來,而改變自身的某些結構或物質組分,從而提高自身耐寒性,順利度過寒冬逆境[24]。
多異瓢蟲作為捕食性天敵昆蟲具有滯育現象,對其進行耐寒性研究是非常有必要的。七星瓢蟲和異色瓢蟲經低溫處理后與正常溫度相比,其過冷卻點和體液結冰點均下降[25],表明耐寒性增強。滯育是某些昆蟲的一種固定遺傳的適應逆境脅迫的屬性,也是一種緩慢的發育狀態,通過生理生化系統發生一系列的改變,如熱激蛋白的表達量發生變化,一些糖類和脂肪類增加,使昆蟲的活動能力減弱,進而提高昆蟲的耐寒性[2627]。
由于多異瓢蟲卵、1齡、2齡幼蟲體型較小,智能昆蟲過冷卻點測定儀的檢測探頭無法很好地與試蟲接觸,導致結果可能存在偏差。本研究選用最適溫度下飼養的多異瓢蟲3齡幼蟲、4齡幼蟲、蛹、2日齡雌成蟲和2日齡雄成蟲進行過冷卻點及體液結冰點的測定。結果表明,在多異瓢蟲整個世代周期中,蛹的耐寒性最強,成蟲的耐寒性最弱,3齡、4齡幼蟲的耐寒性介于蛹和成蟲之間,雄成蟲的耐寒性強于雌成蟲。而已有研究和田間調查表明,多異瓢蟲以成蟲越冬,其成蟲的滯育屬于低溫短日照類型,可能是隨著季節的變化,即日照逐漸縮短、溫度逐漸降低,成蟲逐漸進入滯育狀態,體內一些糖類和脂肪類增加,進而提高成蟲的耐寒性,保證其順利越冬。本研究中多異瓢蟲的飼養條件為最適生存條件,不同發育階段中成蟲的耐寒性最弱,可能由于成蟲自身與耐寒相關的多種物質在相對適合的生境中并未發揮耐寒作用,而野生多異瓢蟲越冬時環境相對較惡劣,猜測越冬成蟲能夠在寒冷的冬天調節自身的某些物質,提高自身耐寒性,保證其順利越冬。昆蟲的抗寒能力與外界環境溫度的變化密切相關,此次結果為后續不同溫度處理或冷馴化后的多異瓢蟲的抗寒性研究奠定了基礎,也為充分利用和保護天敵提供了數據支撐。
參考文獻
[1] 秦昕云, 何曉玲, 沙麗塔娜提·吐松嘎孜, 等. 多異瓢蟲對棉蚜取食行為的影響[J]. 新疆農業大學學報, 2020, 43(1):6265.
[2] 文霞, 陳鳳, 程令, 等. 多異瓢蟲對白毛蚜的捕食功能反應[J]. 新疆農業大學學報, 2019, 42(4): 272275.
[3] 杜軍利, 武德功, 劉長仲. 異色瓢蟲和多異瓢蟲對兩種色型豌豆蚜的捕食偏好研究[J]. 中國生態農業學報, 2015, 23(1): 102109.
[4] PERVEZ A, MAIA A, BOZDOGˇAN H. Reproduction and demography of an Aphidophagous ladybird, Hippodamia variegata on six aphid species [J]. International Journal of Tropical Insect Science,2020,40(3): 542548.
[5] 孔曉霞. 不同溫度條件下多異瓢蟲的生命表及控蚜作用研究[D]. 石河子: 石河子大學, 2018.
[6] 孔琳, 李玉艷, 王孟卿, 等. 多異瓢蟲和異色瓢蟲對草地貪夜蛾低齡幼蟲的捕食能力評價[J]. 中國生物防治學報, 2019, 35(5): 709714.
[7] 宋雙, 姜彩鴿, 趙亞楠, 等. 多異瓢蟲對葡萄斑葉蟬的捕食作用[J]. 中外葡萄與葡萄酒, 2019(4): 4144.
[8] 安瑞軍, 李文秀, 張繼星, 等. 多異瓢蟲Hippodamia variegata(Goeze)生物學初步研究[J].哲里木畜牧學院學報, 2000(3): 1416.
[9] 王允華, 劉寶森, 傅慧鐘, 等. 多異瓢蟲生活習性及發生規律的研究[J]. 昆蟲知識, 1984(1): 1922.
[10]岳健, 何嘉, 張蓉, 等. 不同溫度下多異瓢蟲實驗種群生命表[J]. 昆蟲知識, 2009, 46(6): 921925.
[11]JACK F, ANDREW G H, GUY W. Growth and development rates have different thermal responses [J]. The American Naturalist, 2011, 178(5): 668678.
[12]歐陽芳, 戈峰. 昆蟲抗凍耐寒能力的測定與分析方法[J]. 應用昆蟲學報, 2014, 51(6): 16461652.
[13]任小云, 張禮生, 齊曉陽, 等. 滯育七星瓢蟲的代謝適應與抗寒性評價[J]. 環境昆蟲學報, 2015, 37(6): 11951202.
[14]郭紅雙. 異色瓢蟲越冬代不同斑型的遺傳多樣性和耐寒性研究[D]. 杭州: 杭州師范大學, 2015.
[15]羅舉, 俞俊杰, 周明好, 等. 草地貪夜蛾過冷卻點和結冰點的測定[J]. 浙江農業科學, 2020, 61(3): 428432.
[16]秦建洋, 蘭建軍, 張蕾, 等. 勞氏黏蟲幼蟲和蛹過冷卻點及結冰點的測定[J]. 中國植保導刊, 2018, 38(8): 3338.
[17]段云博, 吳海盼, 鄭霞林, 等. 鉤紋皮蠹不同蟲態過冷卻點及結冰點的測定[J]. 植物保護, 2018, 44(2): 116121.
[18]姚舉, 姬華, 王東, 等. 棉田優勢天敵多異瓢蟲成蟲對棉蚜捕食功能的研究[J]. 新疆農業科學, 2005(4): 262264.
[19]王俊剛, 李永剛, 楊德松, 等. 北疆多異瓢蟲耐饑能力的研究[J]. 中國棉花, 2004(5): 1214.
[20]謝欣. 八種除草劑對堿蓬及二種天敵昆蟲的安全性評價[D]. 石河子: 石河子大學, 2020.
[21]潘洪生, 李號賓, 丁瑞豐, 等. 多異瓢蟲對棉黑蚜的捕食能力[J]. 中國生物防治學報, 2020, 36(4): 628631.
[22]RENAULT D, SALIN C, VANNIER G, et al. Survival at low temperatures in insects: what is the ecological significance of the supercooling point? [J]. CryoLetters, 2002, 23(4): 217228.
[23]ASAHINA E. Frost resistance in insects [J]. Advances in Insect Physiology, 1970, 6: 149.
[24]史彩華, 胡靜榮, 李傳仁, 等. 環境脅迫下昆蟲的耐寒適應機制研究進展[J]. 植物保護, 2016,42(6): 2128.
[25]LABRIE G, CODERRE D, LUCAS , et al. Overwintering strategy of multicolored Asian lady beetle (Coleoptera: Coccinellidae): cold-free space as a factor of invasive success [J]. Annals of the Entomological Society of America, 2008, 101(5): 860866.
[26]RAGLAND G J, DENLINGER D L, HAHN D A. Mechanisms of suspended animation are revealed by transcript profiling of diapause in the flesh fly [J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(33): 1490914914.
[27]PAPANASTASIOU S A, NESTEL D, DIAMANTIDIS A D, et al. Physiological and biological patterns of a highland and a coastal population of the European cherry fruit fly during diapause [J]. Journal of Insect Physiology, 2011, 57(1): 8399.
(責任編輯:楊明麗)
